Fermentability of Novel Type-4 Resistant Starches in In Vitro System

Abstract
:1. Introduction
2. Materials and Methods
2.1. Resistant Starches Analyzed
2.2. Predigestion
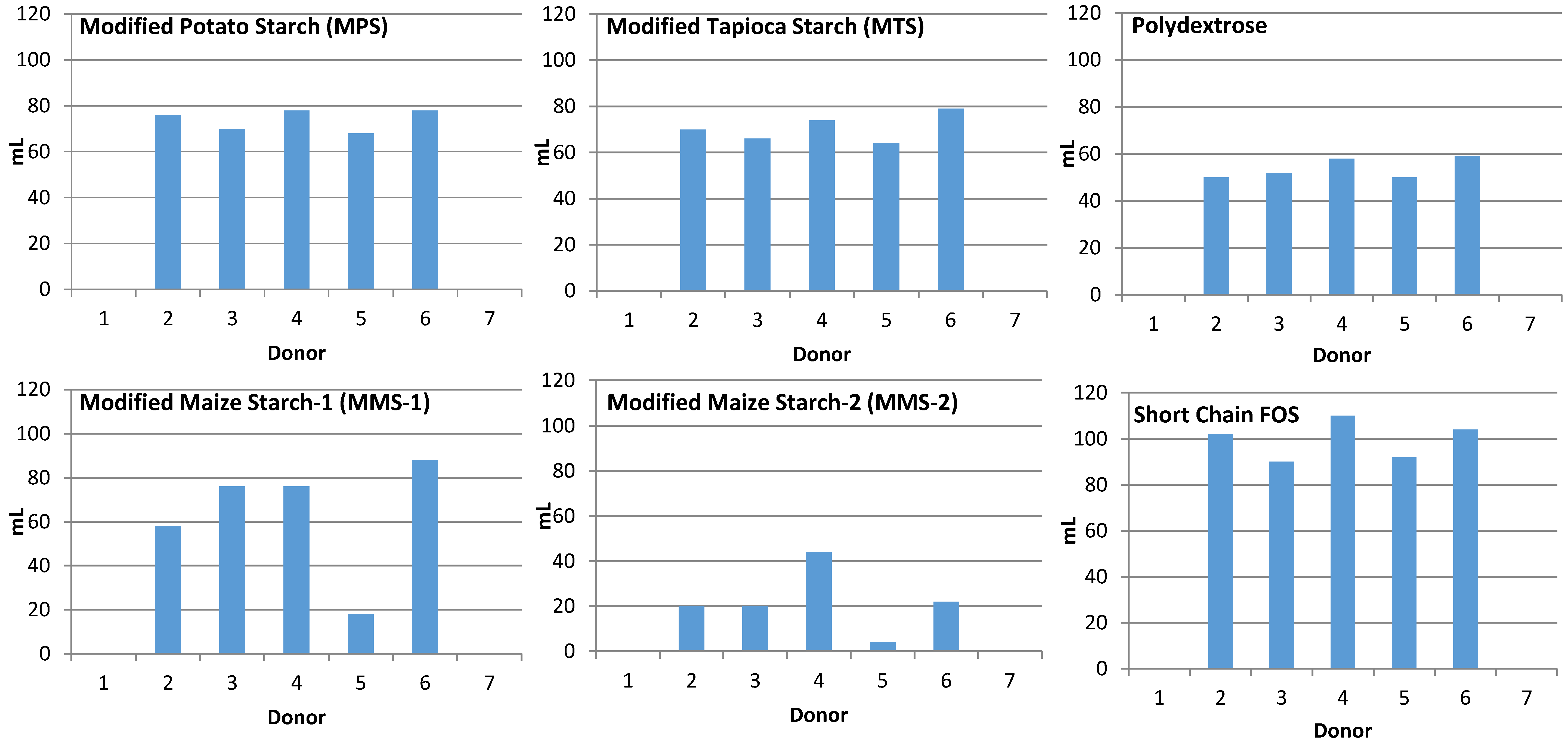
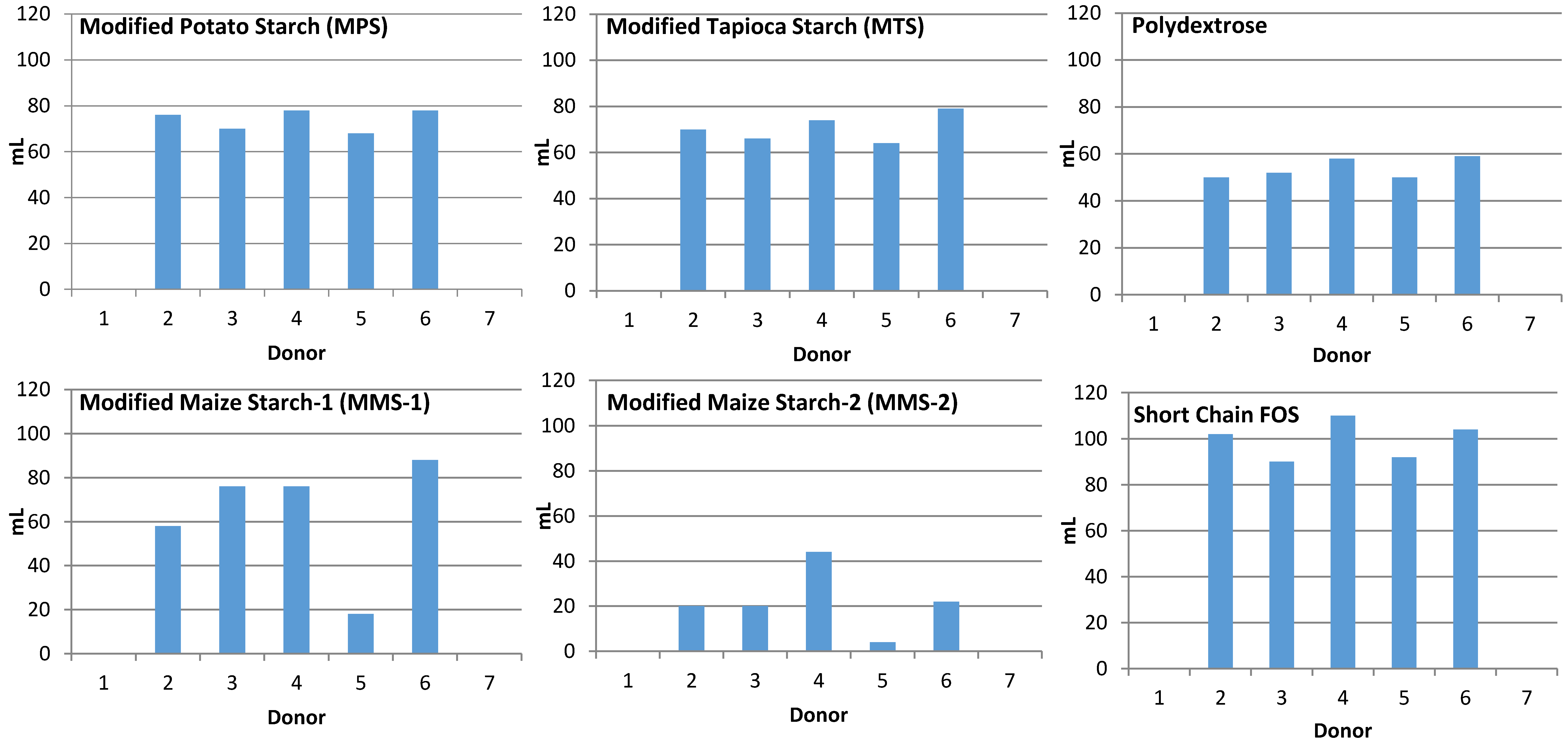
2.3. Fecal Collection and Donor Information
2.4. Consent
2.5. Fermentation
2.6. Gas Analysis
2.7. pH Analysis
2.8. SCFA Analysis
2.9. Statistical Analysis
3. Results
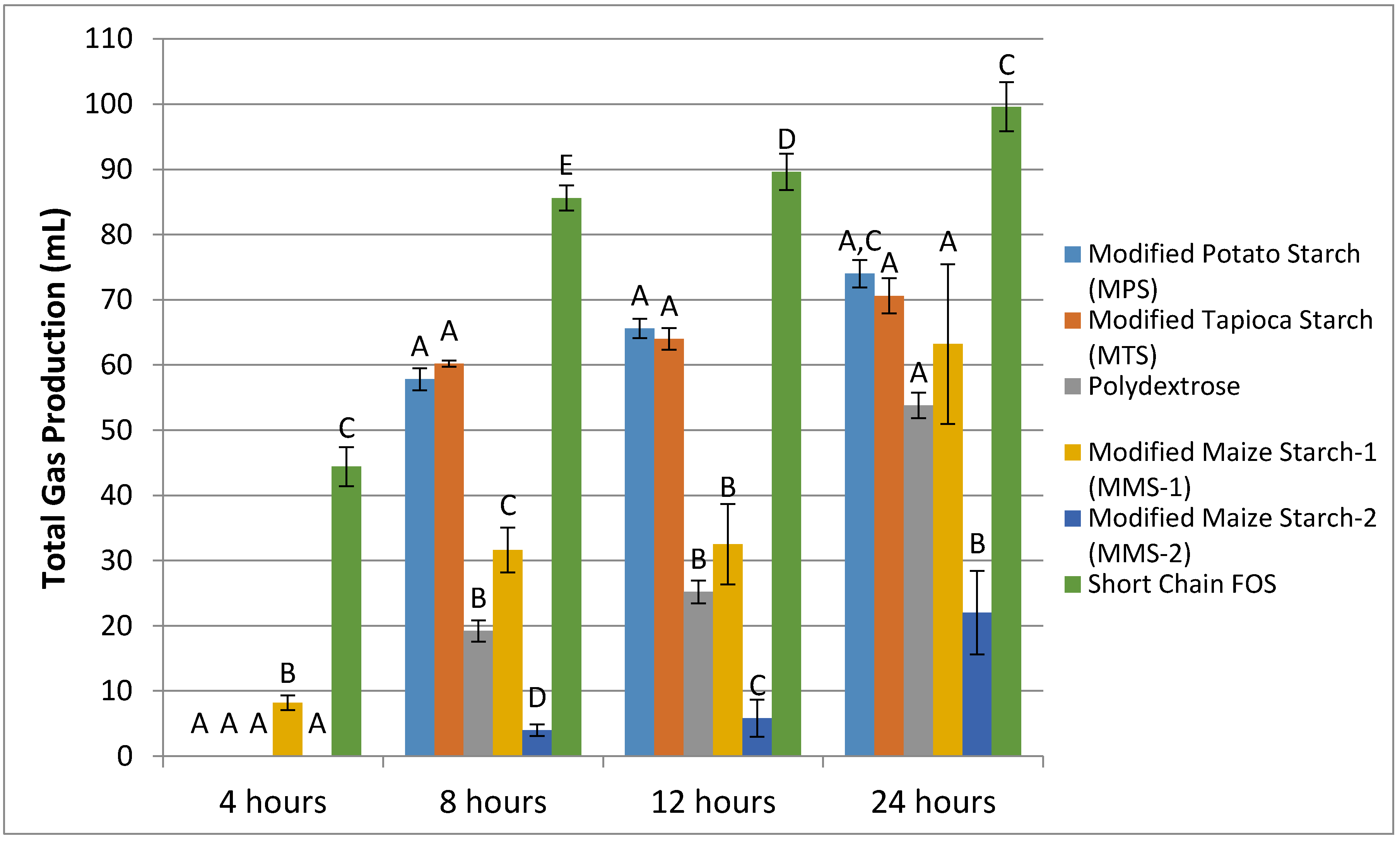
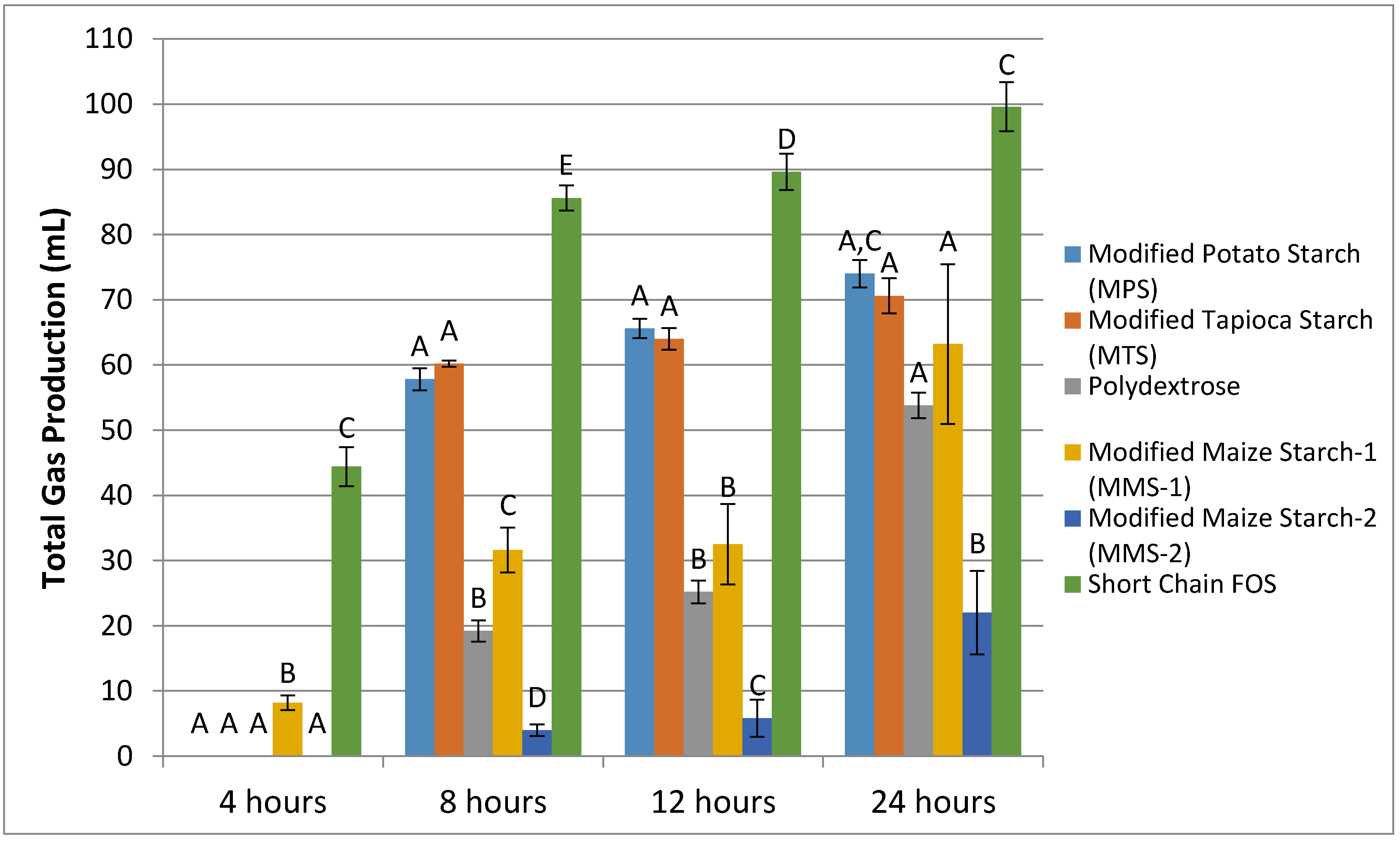
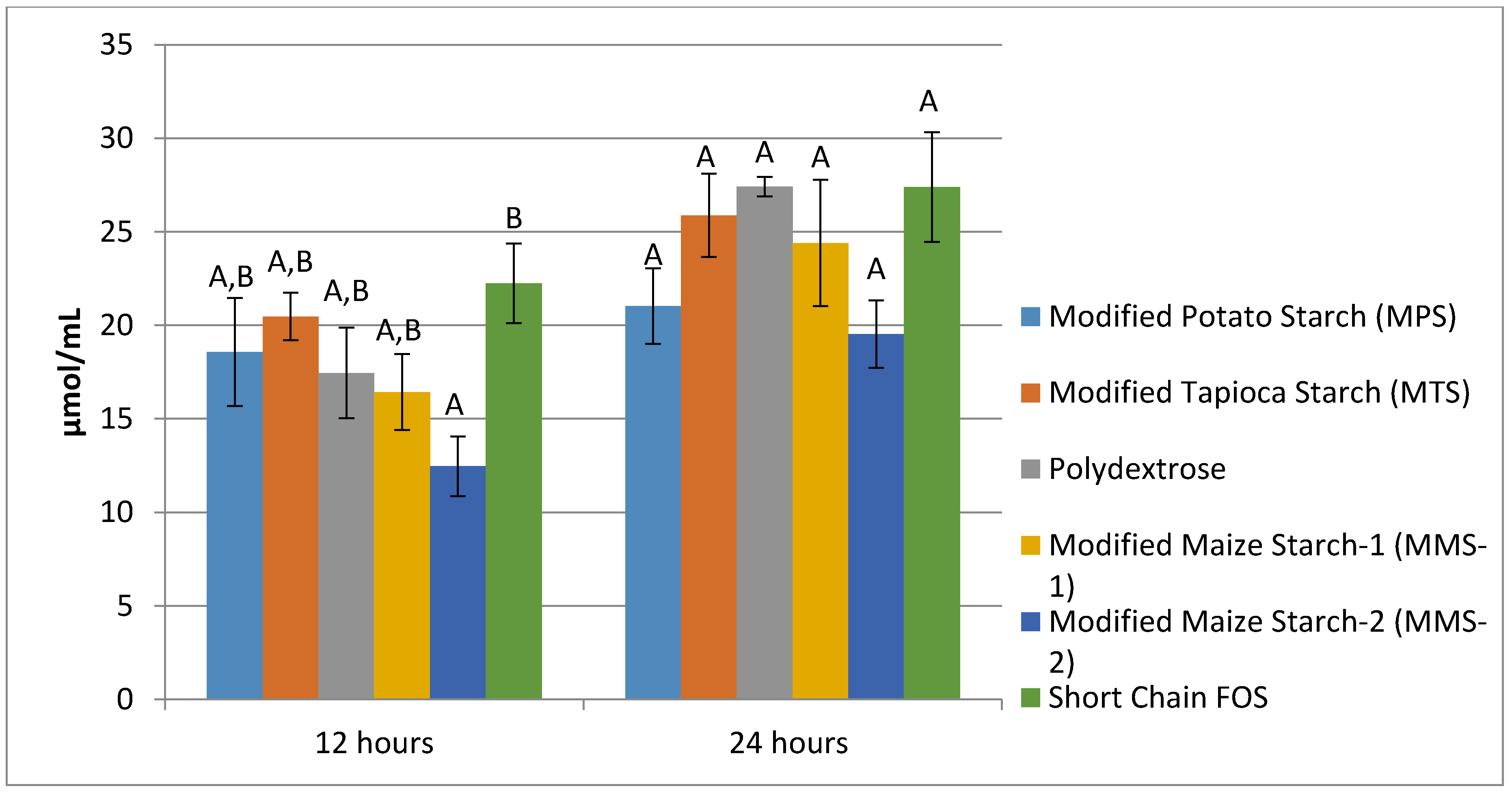
3.1. Gas Production
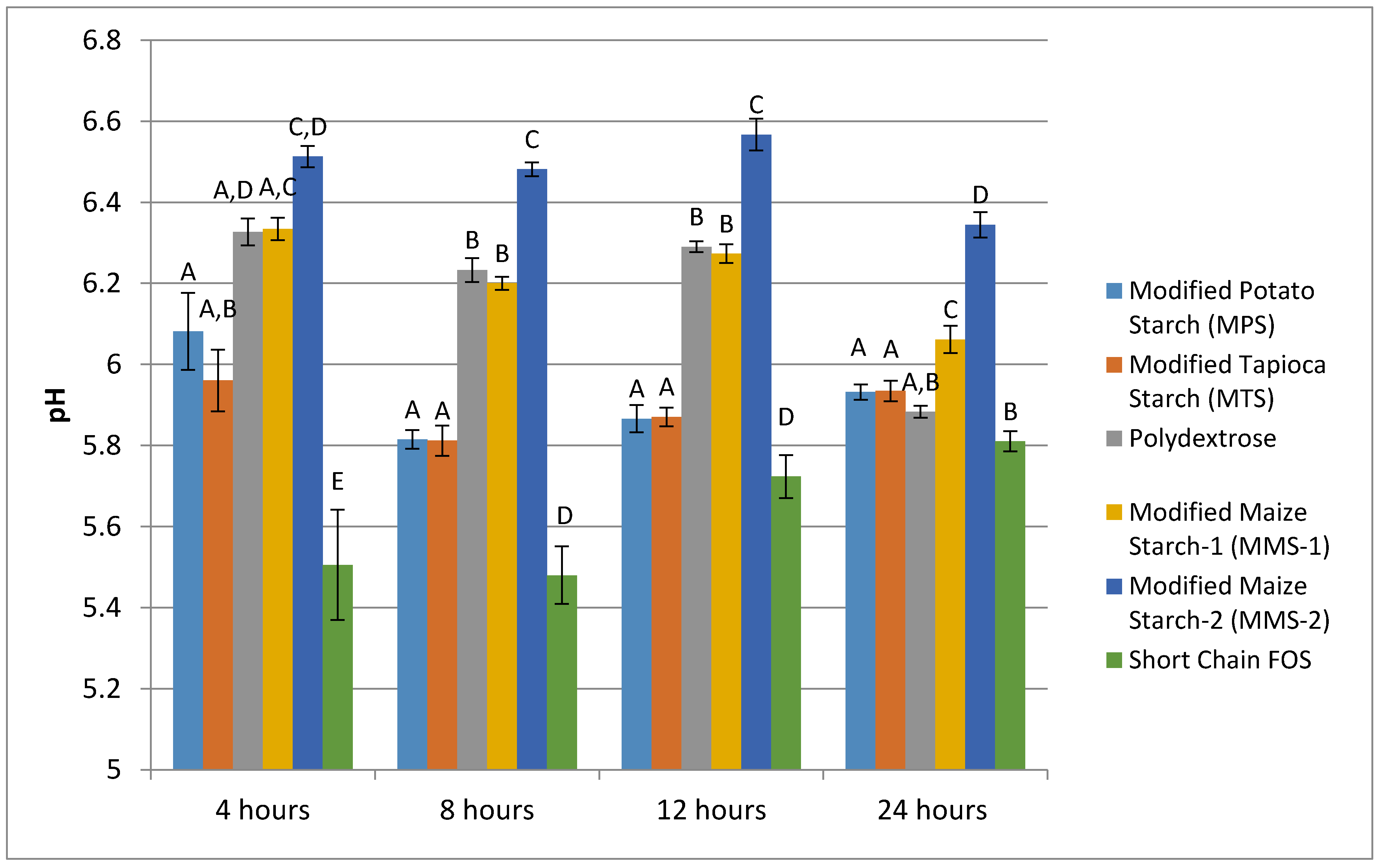
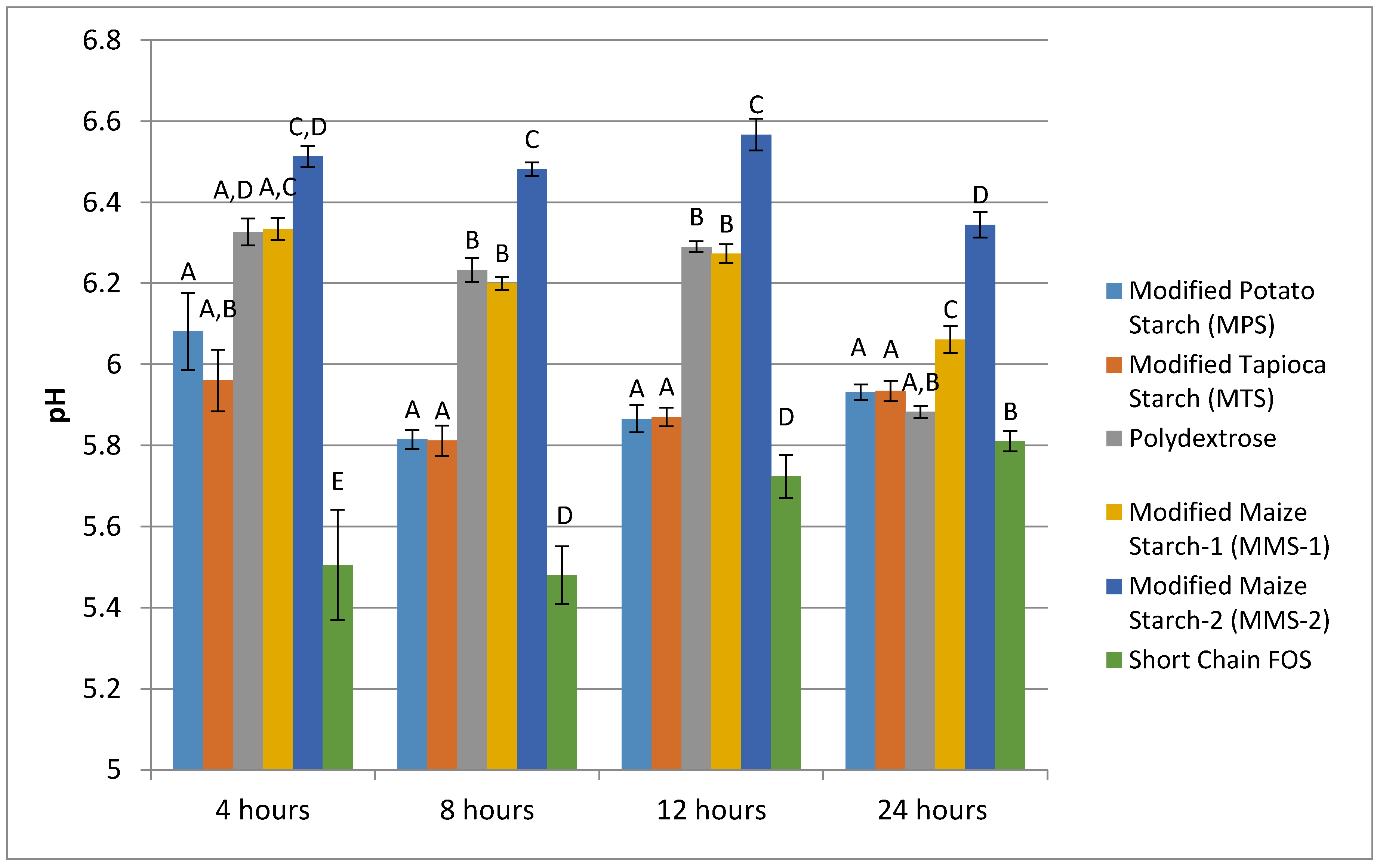
3.2. pH Production
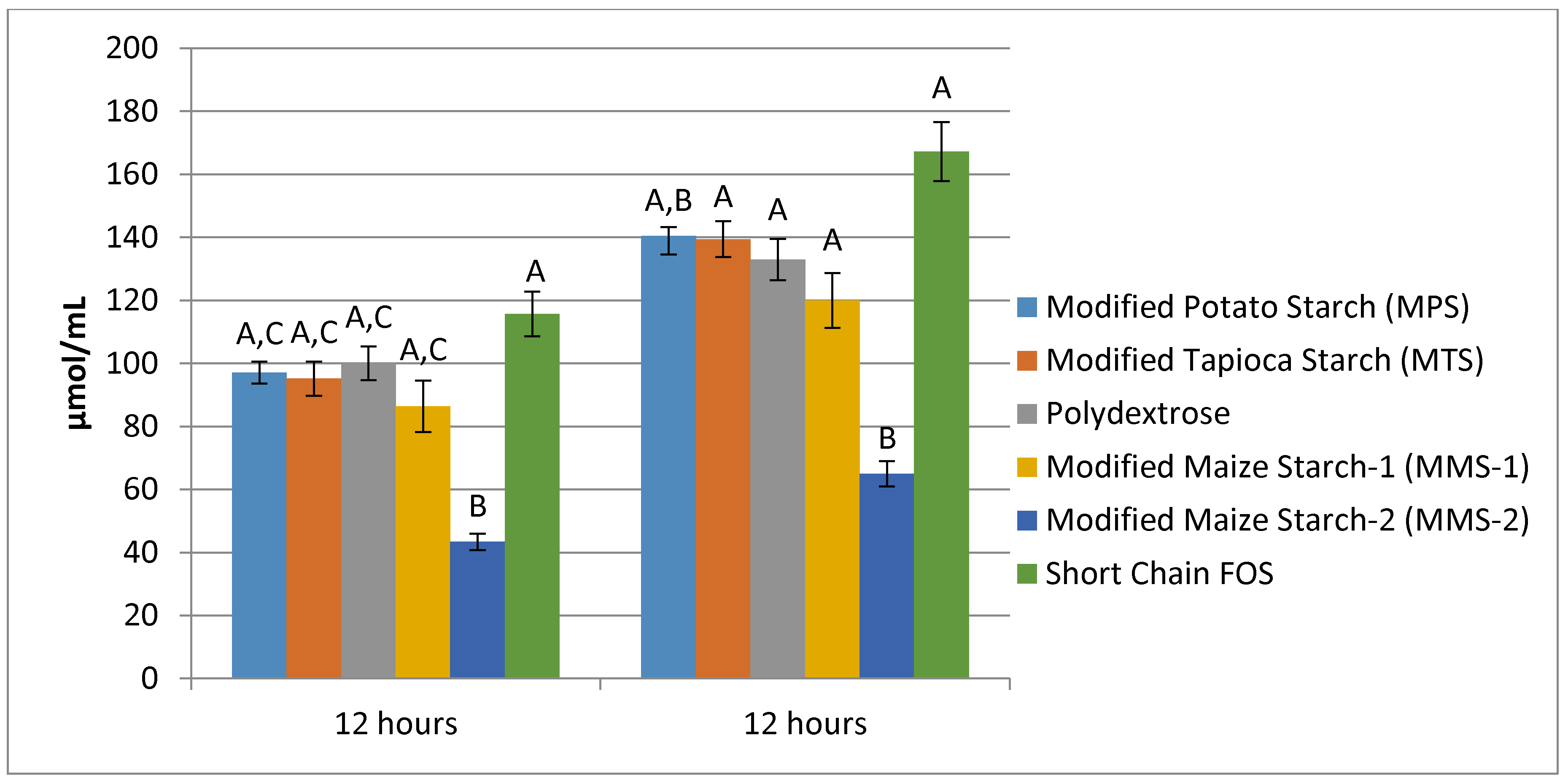
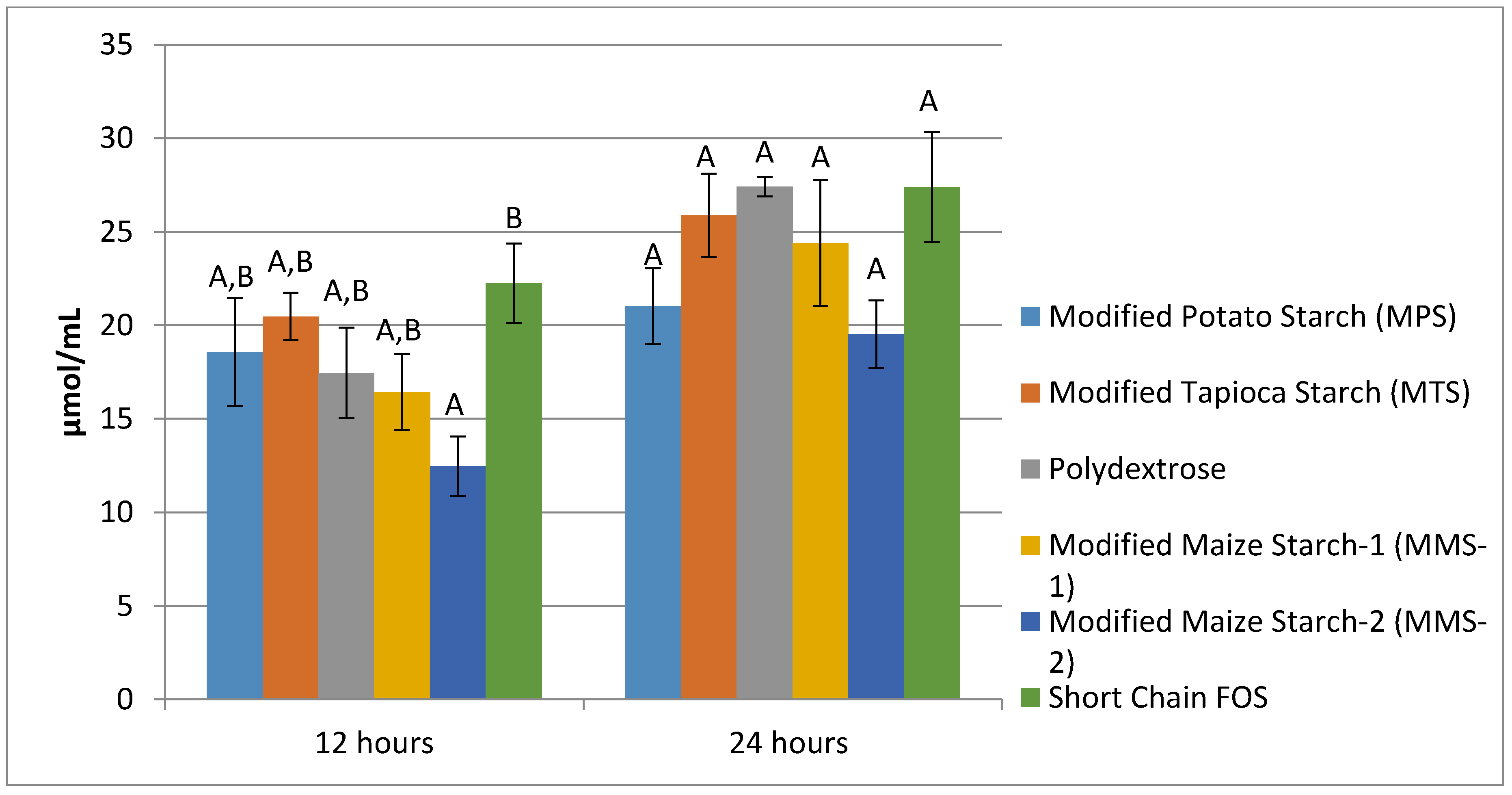
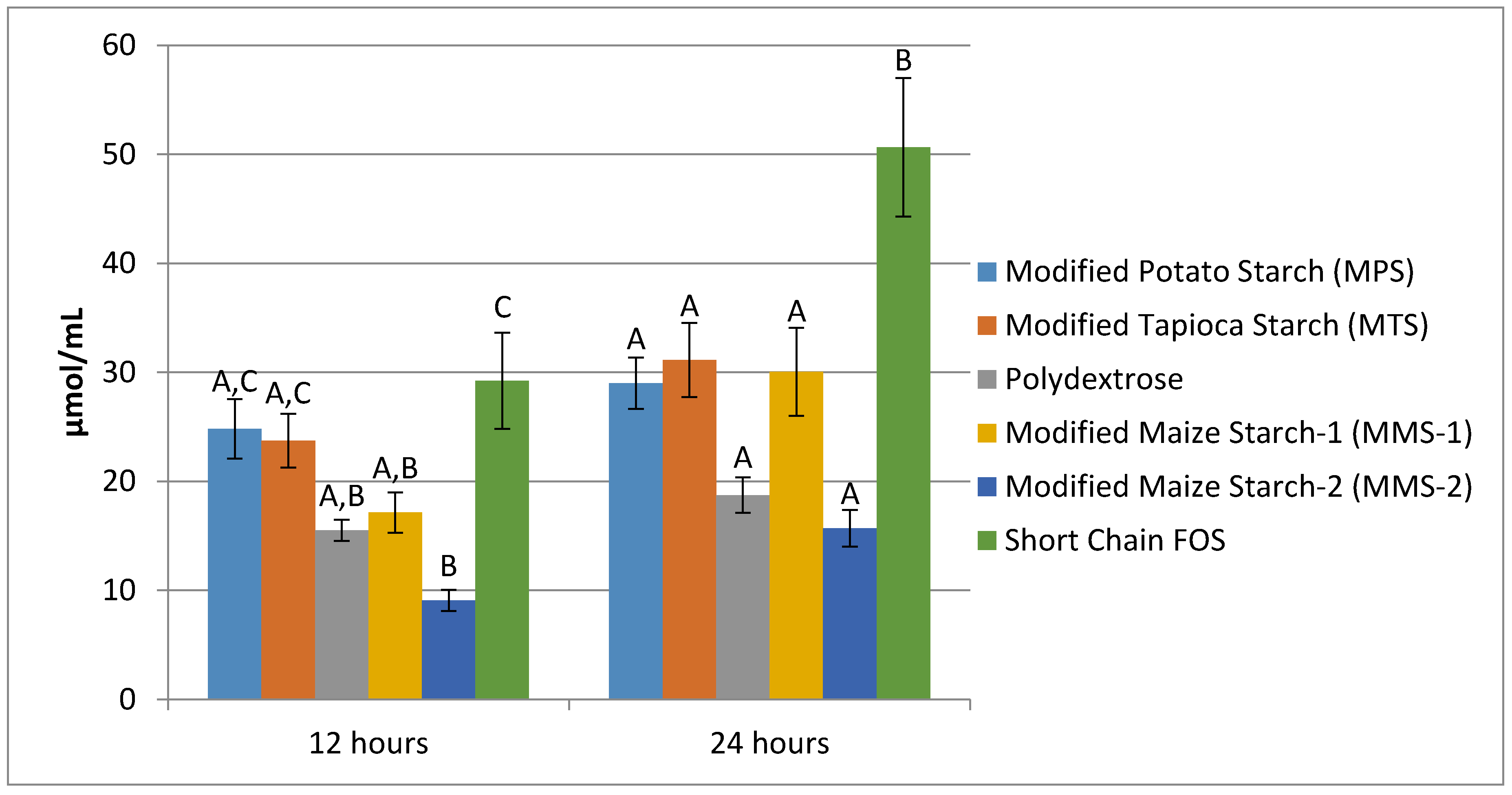
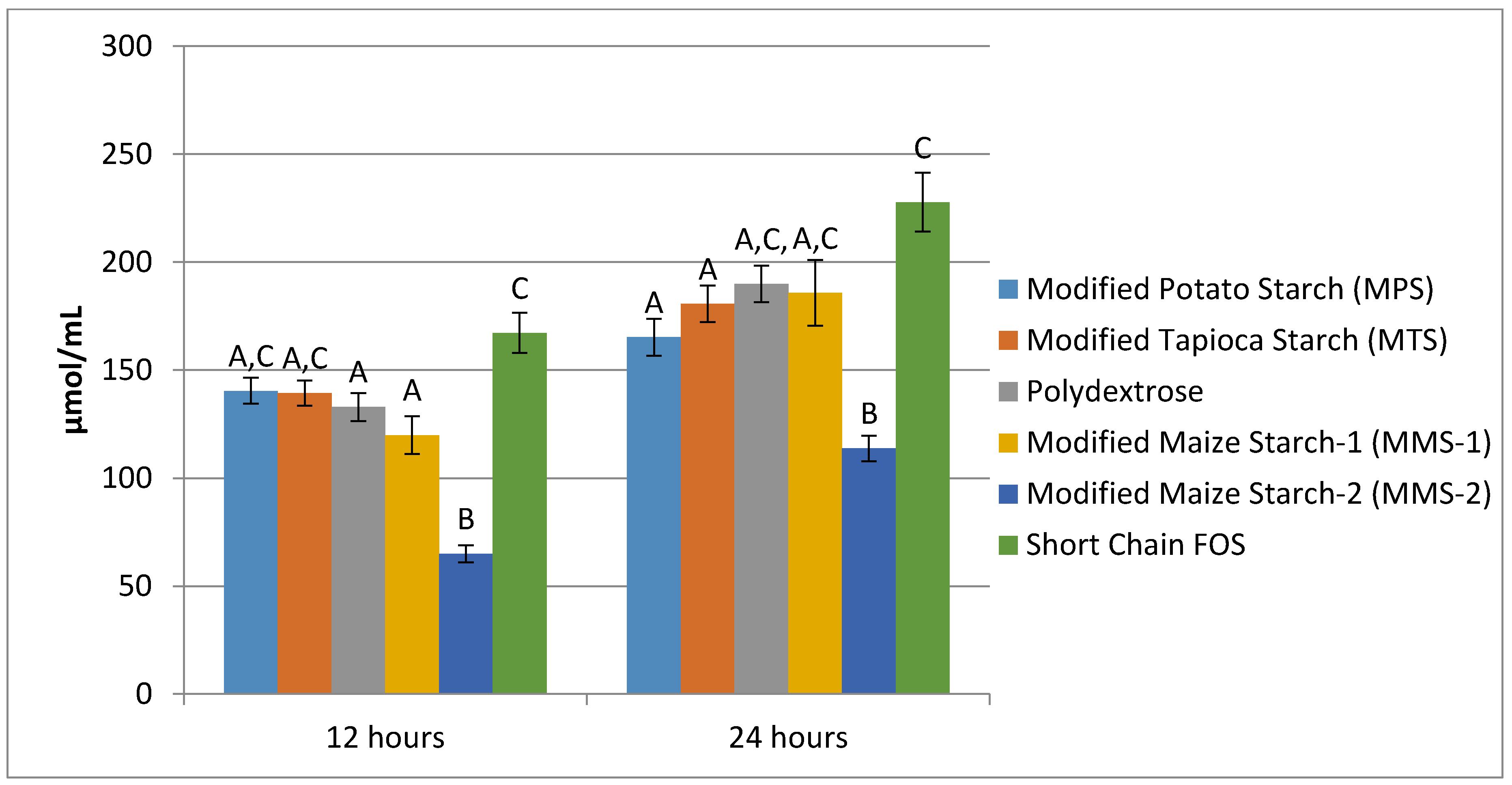
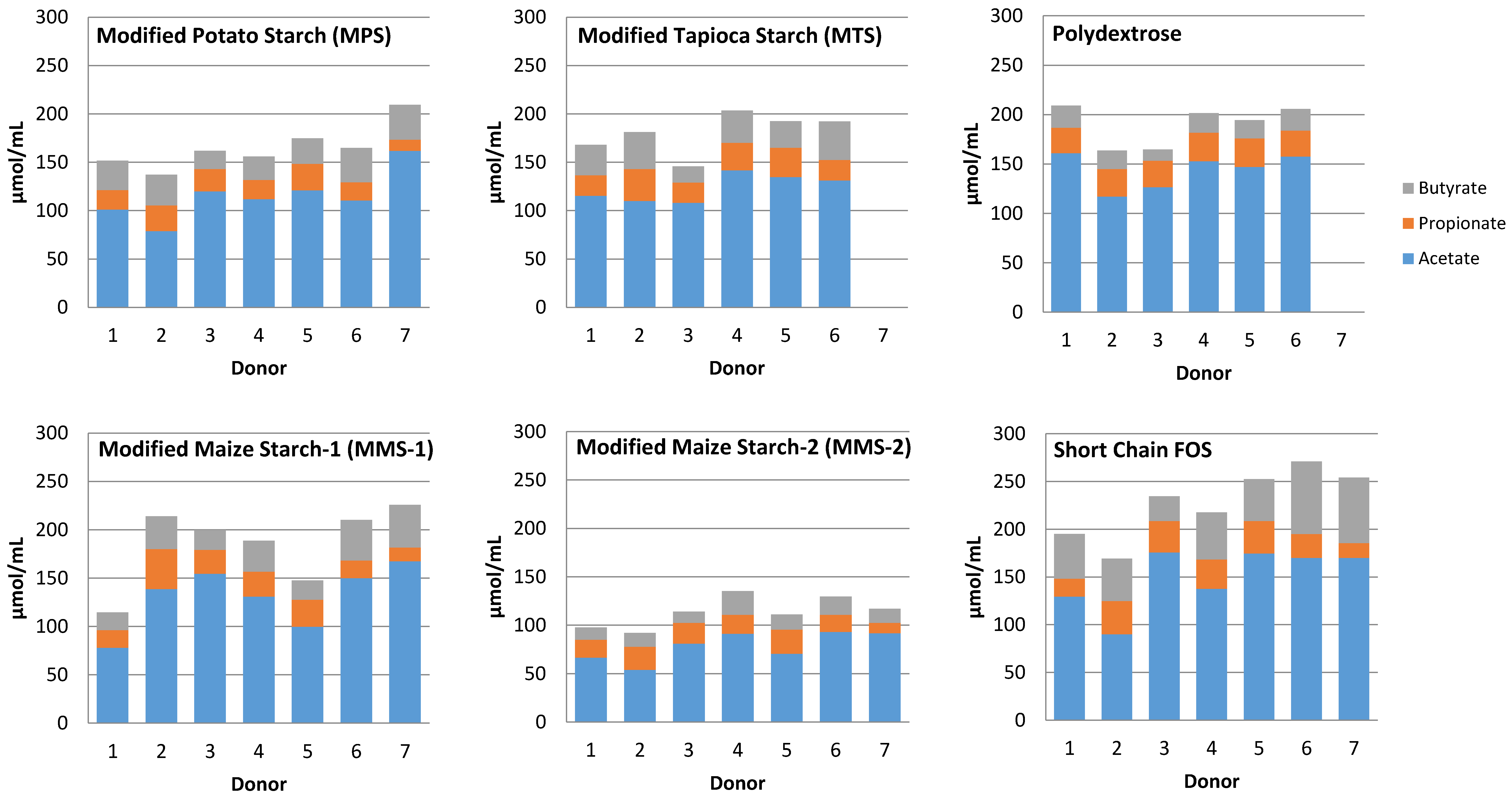
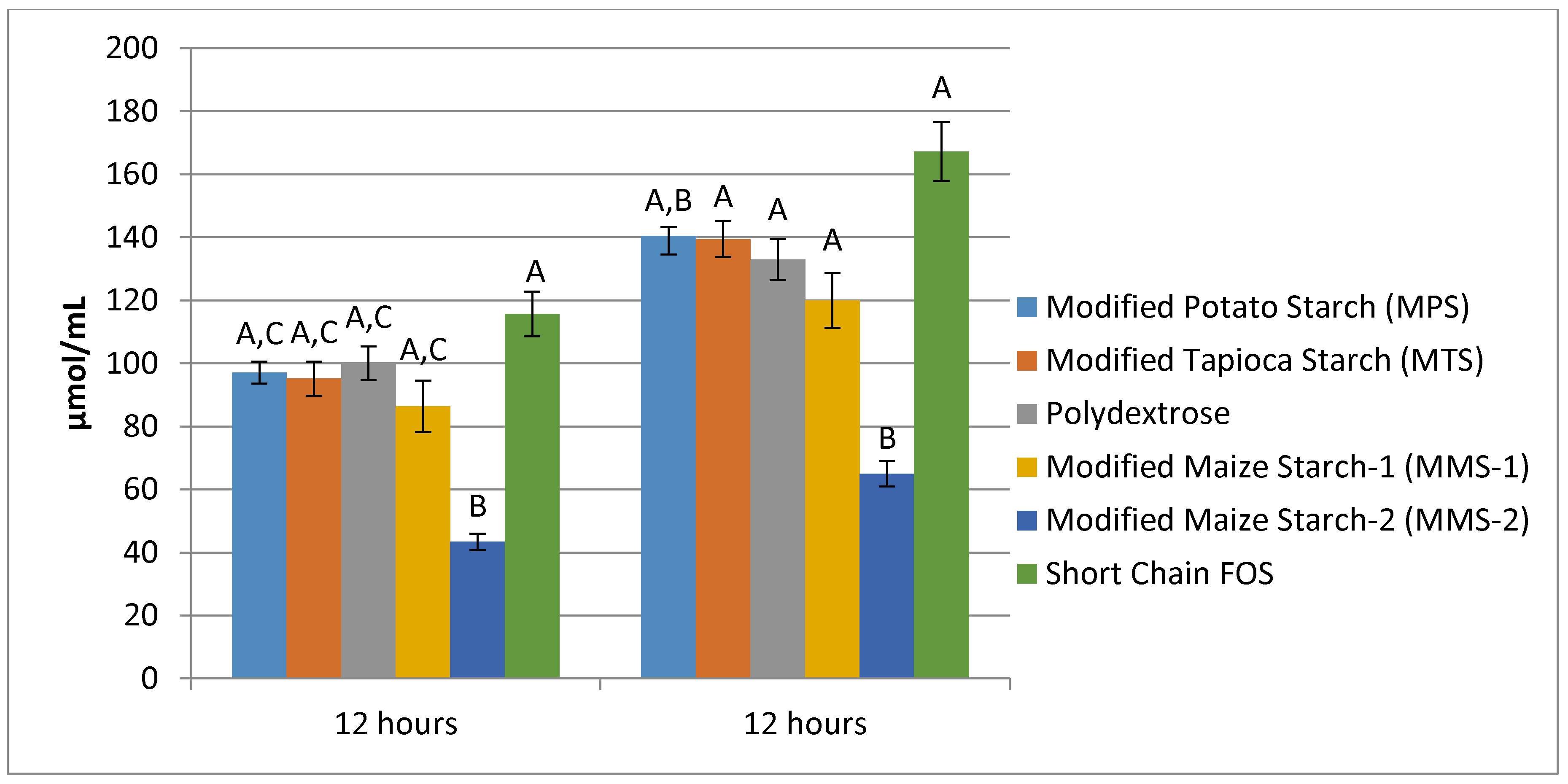
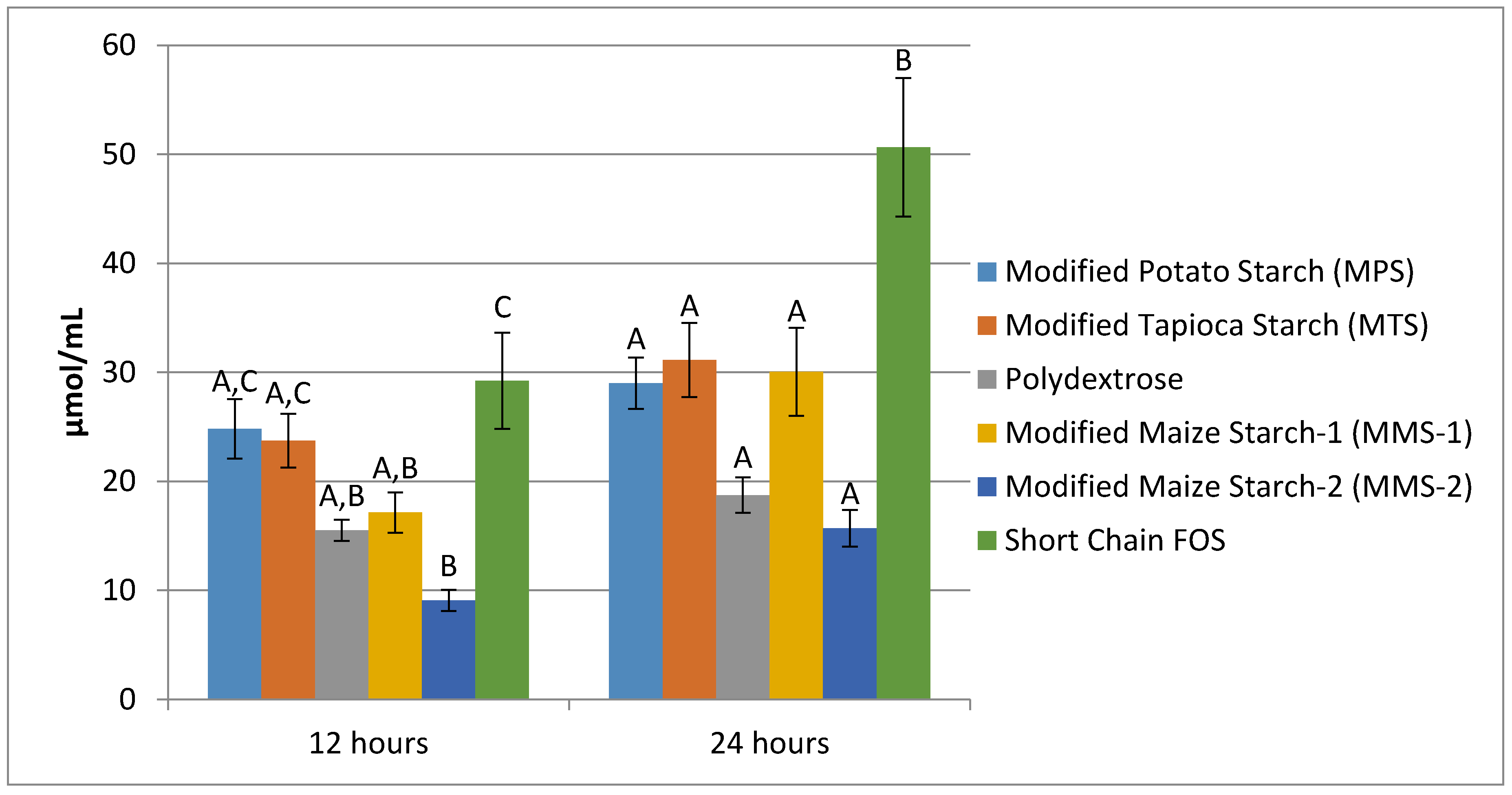
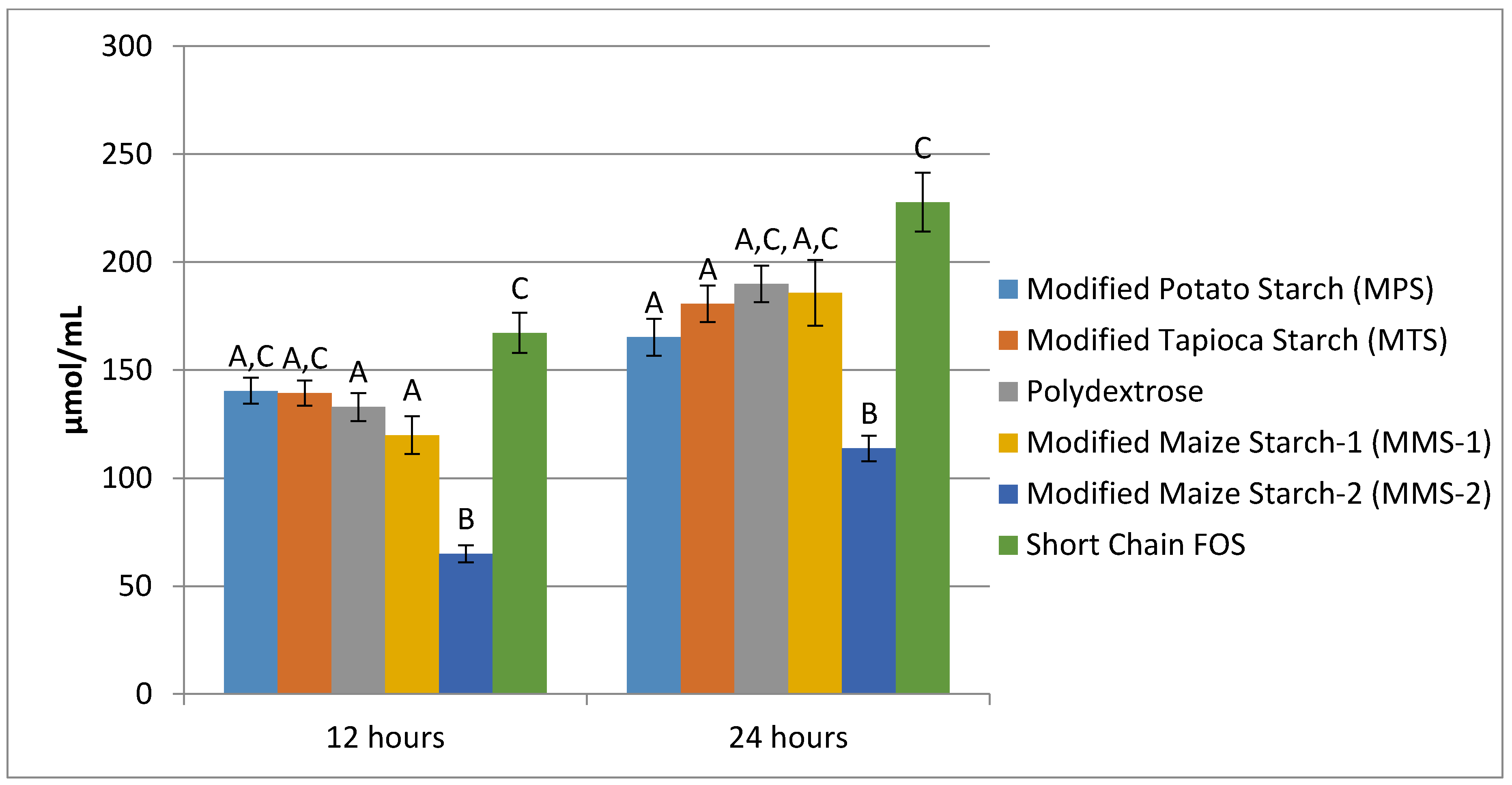
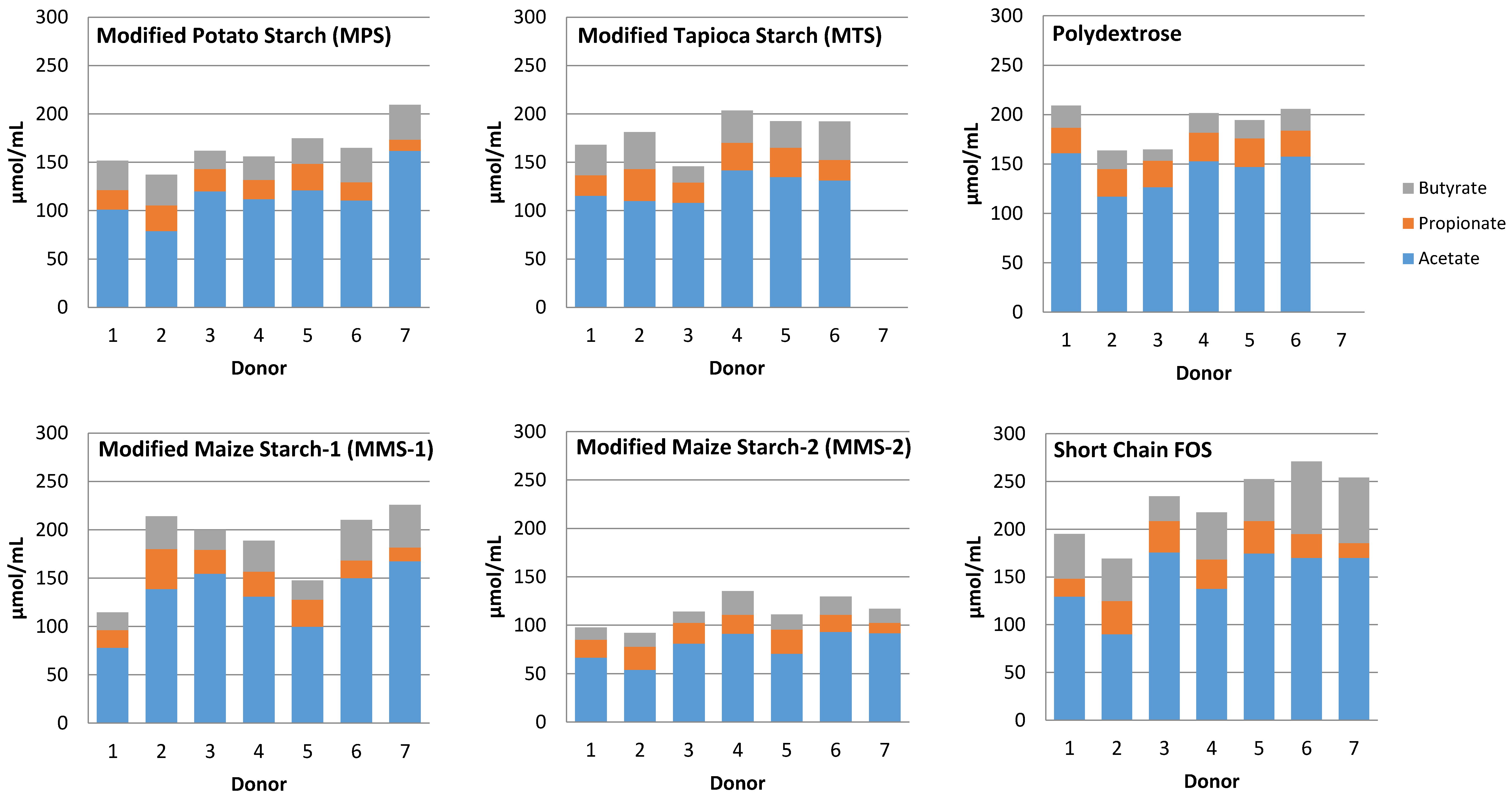
3.3. SCFA Production
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dahl, W.J.; Stewart, M.L. Position of the Academy of Nutrition and Dietetics: Health Implications of Dietary Fiber. J. Acad. Nutr. Diet. 2015, 115, 1861–1870. [Google Scholar] [CrossRef] [PubMed]
- King, D.E.; Mainous, A.G.; Lambourne, C.A. Trends in Dietary Fiber Intake in the United States, 1999–2008. J. Acad. Nutr. Diet. 2012, 112, 642–648. [Google Scholar] [CrossRef] [PubMed]
- United States Food and Drug Administration. Scientific Evaluation of the Evidence on the Beneficial Physiological Effects of Isolated or Synthetic Non-DiGestible Carbohydrates Submitted as a Citizen Petition (21 CFR 10.30): Guidance for Industry; United States Food and Drug Administration: Silver Spring, MD, USA, 2016.
- Office of Nutrition and Food Labeling; Center for Food Safety and Applied Nutrition; United States Food and Drug Administration; US Department of Health and Human Services. Science Review of Isolated and Synthetic Non-Digestible Carbohydrates; United States Food and Drug Administration: Silver Spring, MD, USA, 2016.
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.-L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant starch: Promise for improving human health. Adv. Nutr. 2013, 4, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, S.; Nugent, A.P. Health effects of resistant starch. Nutr. Bull. 2017, 42, 10–41. [Google Scholar] [CrossRef]
- Homayouni, A.; Amini, A.; Keshtiban, A.K.; Mortazavian, A.M.; Esazadeh, K.; Pourmoradian, S. Resistant starch in food industry: A changing outlook for consumer and producer. Starch Stärke 2014, 66, 102–114. [Google Scholar] [CrossRef]
- Charalampopoulos, D.; Wang, R.; Pandiella, S.S.; Webb, C. Application of cereals and cereal components in functional foods: A review. Int. J. Food Microbiol. 2002, 79, 131–141. [Google Scholar] [CrossRef]
- Baixauli, R.; Salvador, A.; Martínez-Cervera, S.; Fiszman, S.M. Distinctive sensory features introduced by resistant starch in baked products. LWT Food Sci. Technol. 2008, 41, 1927–1933. [Google Scholar] [CrossRef]
- Wong, T.H.T.; Louie, J.C.Y. The relationship between resistant starch and glycemic control: A review on current evidence and possible mechanisms. Starch Stärke 2017, 69, 1600205. [Google Scholar] [CrossRef]
- Dodevska, M.S.; Sobajic, S.S.; Djordjevic, P.B.; Dimitrijevic-Sreckovic, V.S.; Spasojevic-Kalimanovska, V.V.; Djordjevic, B.I. Effects of total fibre or resistant starch-rich diets within lifestyle intervention in obese prediabetic adults. Eur. J. Nutr. 2016, 55, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Wanders, A.J.; van den Borne, J.J.G.C.; de Graaf, C.; Hulshof, T.; Jonathan, M.C.; Kristensen, M.; Mars, M.; Schols, H.A.; Feskens, E.J.M. Effects of dietary fibre on subjective appetite, energy intake and body weight: A systematic review of randomized controlled trials. Obes. Rev. 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.J.; Slavin, J.L. The Effect of Fiber on Satiety and Food Intake: A Systematic Review. J. Am. Coll. Nutr. 2013, 32, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Serra, J.; Azpiroz, F.; Malagelada, J. Intestinal gas dynamics and tolerance in humans. Gastroenterology 1998, 115, 542–550. [Google Scholar] [CrossRef]
- Cherrington, C.A.; Hinton, M.; Pearson, G.R.; Chopra, I. Short-chain organic acids at pH 5.0 kill Escherichia coli and Salmonella spp. without causing membrane perturbation. J. Appl. Bacteriol. 1991, 70, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Duncan, S.H.; McWilliam Leitch, E.C.; Child, M.W.; Flint, H.J. pH and peptide supply can radically alter bacterial populations and short-chain fatty acid ratios within microbial communities from the human colon. Appl. Environ. Microbiol. 2005, 71, 3692–3700. [Google Scholar] [CrossRef] [PubMed]
- Trinidad, T.P.; Wolever, T.M.; Thompson, L.U. Effect of acetate and propionate on calcium absorption from the rectum and distal colon of humans. Am. J. Clin. Nutr. 1996, 63, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Younes, H.; Demigné, C.; Rémésy, C. Acidic fermentation in the caecum increases absorption of calcium and magnesium in the large intestine of the rat. Br. J. Nutr. 1996, 75, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Lutz, T.; Scharrer, E. Effect of short-chain fatty acids on calcium absorption by the rat colon. Exp. Physiol. 1991, 76, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Livesey, G. Energy values of unavailable carbohydrate and diets: An inquiry and analysis. Am. J. Clin. Nutr. 1990, 51, 617–637. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- JECFA. Phosphated distarch phosphate: In Toxicological Evaluation of Certain Food Additives. In Proceedings of the 26th JECFA Sess, Rome, Italy, 19–28 April 1982. [Google Scholar]
- Coles, L.T.; Moughan, P.J.; Darragh, A.J. In vitro digestion and fermentation methods, including gas production techniques, as applied to nutritive evaluation of foods in the hindgut of humans and other simple-stomached animals. Anim. Feed Sci. Technol. 2005, 123–124, 421–444. [Google Scholar] [CrossRef]
- Schneider, S.M.; Girard-Pipau, F.; Anty, R.; van der Linde, E.G.M.; Philipsen-Geerling, B.J.; Knol, J.; Filippi, J.; Arab, K.; Hébuterne, X. Effects of total enteral nutrition supplemented with a multi-fibre mix on faecal short-chain fatty acids and microbiota. Clin. Nutr. 2006, 25, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Hospattankar, A.; Deng, P.; Swanson, K.; Slavin, J. Prebiotic Effects and Fermentation Kinetics of Wheat Dextrin and Partially Hydrolyzed Guar Gum in an In Vitro Batch ermentation System. Foods 2015, 4, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Esparza, J.; Swan, J.; Taussig, D.; Combs, J.; Slavin, J. In vitro analysis of partially hydrolyzed guar gum fermentation differences between six individuals. Food Funct. 2016, 7, 1833–1838. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Erickson, J.; Hess, J.; Gould, T.; Slavin, J. Prebiotic Dietary Fiber and Gut Health: Comparing the in vitro Fermentations of Beta-Glucan, Inulin and Xylooligosaccharide. Nutrients 2017, 9, 1361. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.Z.; Zou, D.W.; Li, Z.S.; Zou, X.P.; Zhu, A.Y.; Xu, G.M.; Yin, N.; Gong, Y.F.; Sun, Z.X.; Man, X.H. Study of visceral hypersensitivity in irritable bowel syndrome. Chin. J. Dig. Dis. 2004, 5, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Bonnema, A.L.; Kolberg, L.W.; Thomas, W.; Slavin, J.L. Gastrointestinal tolerance of chicory inulin products. J. Am. Diet. Assoc. 2010, 110, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Timm, D.A.; Stewart, M.L.; Hospattankar, A.; Slavin, J.L. Wheat Dextrin, Psyllium, and Inulin Produce Distinct Fermentation Patterns, Gas Volumes, and Short-Chain Fatty Acid Profiles In Vitro. J. Med. Food 2010, 13, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Thornton, J.R. High Colonic pH Promotes Colorectal Cancer. Lancet 1981, 317, 1081–1083. [Google Scholar] [CrossRef]
- Newmark, H.L.; Lupton, J.R. Determinants and consequences of colonic luminal pH: Implications for colon cancer. Nutr. Cancer 1990, 14, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R.; Harris, P.J. Studies on the role of specific dietary fibres in protection against colorectal cancer. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1996, 350, 173–184. [Google Scholar] [CrossRef]
- Venkataraman, A.; Sieber, J.R.; Schmidt, A.W.; Waldron, C.; Theis, K.R.; Schmidt, T.M. Variable responses of human microbiomes to dietary supplementation with resistant starch. Microbiome 2016, 4, 33. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbohydrate Source | Sample Tradename, Chemical Characterization (If Applicable) | Grams of Fiber/100 g | Grams of Starch/100 g | Grams of Sugar/100 g |
|---|---|---|---|---|
| Modified potato starch (MPS) | VERSAFIBETM 1490 dietary fiber, Distarch phosphate | 74.2 | 11.8 | 0 |
| Modified tapioca starch (MTS) | NOVELOSETM 3490 dietary fiber, Distarch phosphate | 79.9 | 7.9 | 0 |
| Modified maize starch (MMS-1) | VERSAFIBETM 2470 dietary fiber, Acid hydrolyzed and heat treated | 65 | 24.9 | 0 |
| Modified maize starch (MMS-2) | VERSAFIBETM 2480 dietary fiber, Distarch phosphate | 71 | 14 | 0 |
| Polydextrose | n/a | 87.5 | 0 | 4.5 |
| Short-chain fructooligosaccharide | NUTRAFLORA® soluble prebiotic fiber | 92.5 | 0 | 4.3 |
| Donor 1 | Donor 2 | Donor 3 | Donor 4 | Donor 5 | Donor 6 | Donor 7 | |
|---|---|---|---|---|---|---|---|
| Sex | Female | Male | Male | Male | Female | Male | Female |
| Age | 21 | 21 | 23 | 27 | 25 | 28 | 28 |
| BMI | 20.0 | 30.1 | 24.0 | 25.2 | 17.7 | 27.7 | 25.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erickson, J.M.; Carlson, J.L.; Stewart, M.L.; Slavin, J.L. Fermentability of Novel Type-4 Resistant Starches in In Vitro System. Foods 2018, 7, 18. https://doi.org/10.3390/foods7020018
Erickson JM, Carlson JL, Stewart ML, Slavin JL. Fermentability of Novel Type-4 Resistant Starches in In Vitro System. Foods. 2018; 7(2):18. https://doi.org/10.3390/foods7020018
Chicago/Turabian StyleErickson, Jennifer M., Justin L. Carlson, Maria L. Stewart, and Joanne L. Slavin. 2018. "Fermentability of Novel Type-4 Resistant Starches in In Vitro System" Foods 7, no. 2: 18. https://doi.org/10.3390/foods7020018
APA StyleErickson, J. M., Carlson, J. L., Stewart, M. L., & Slavin, J. L. (2018). Fermentability of Novel Type-4 Resistant Starches in In Vitro System. Foods, 7(2), 18. https://doi.org/10.3390/foods7020018