Optimum Thermal Processing for Extended Shelf-Life (ESL) Milk

Abstract
:1. Introduction
2. Heating Methods
2.1. Direct Heating
2.2. Indirect Heating
2.3. Nominal Temperature–Time Combinations
2.4. Alternative Direct Heating Systems
2.4.1. The Pure-Lac™ System
2.4.2. The Innovative Steam Injection (ISI) Process
2.5. The Millisecond Technologies (MST) Process
3. Microbiological Considerations
3.1. Microbiological Issues Related to the Heating Process
3.1.1. Psychrotrophic Spore-Formers
3.1.2. Paenibacillus
3.1.3. B. cereus
3.2. Post-Processing Contamination
3.3. Aseptic Packaging of ESL Milk
3.4. Storage Temperature
4. Optimizing the Flavor of ESL Milk
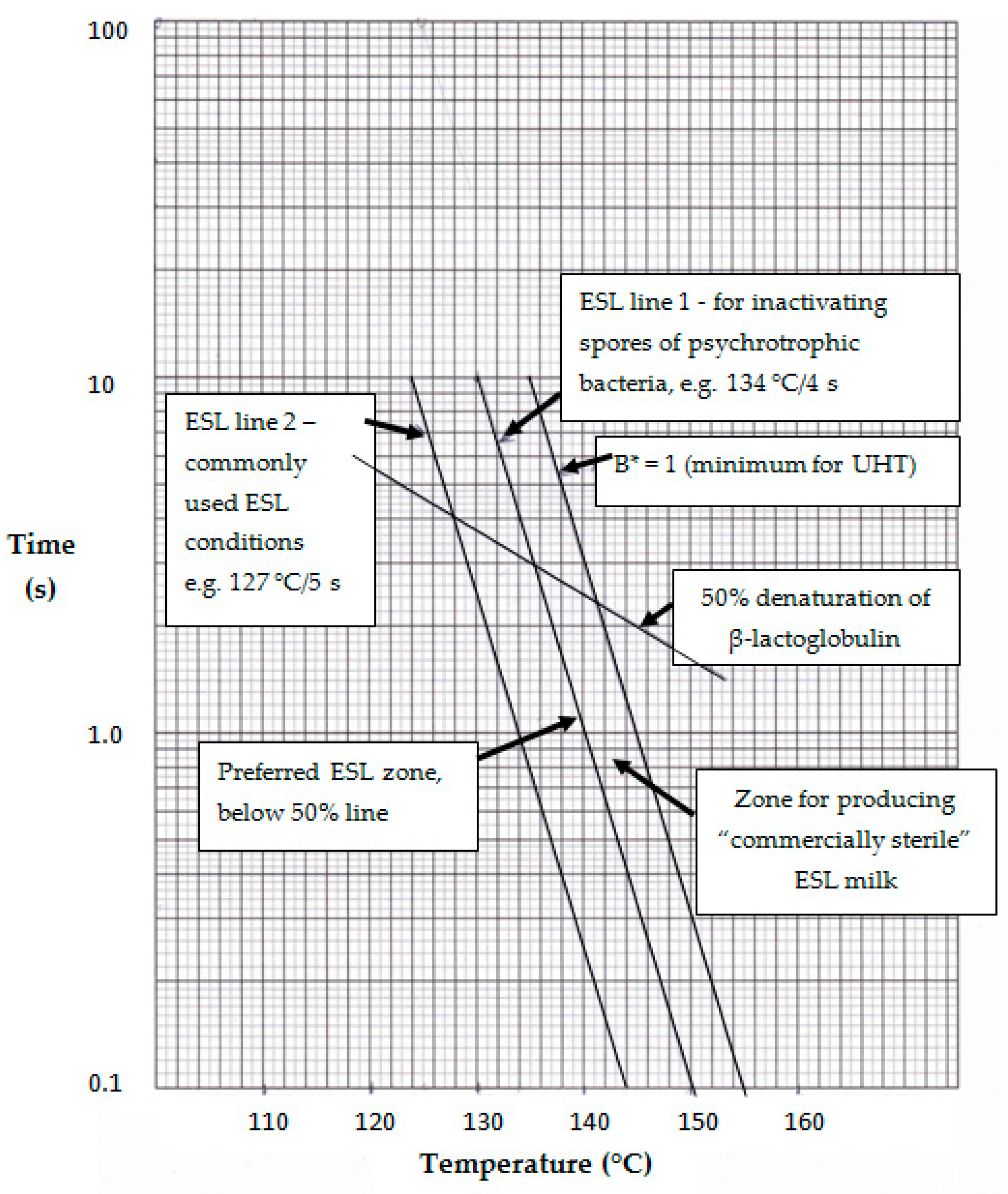
5. Optimizing ESL Heating Conditions
6. Assessment of Some Possible Temperature–Time Conditions for ESL Processing
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Rysstad, G.; Kolstad, J. Conference contribution: Extended shelf life milk-advances in technology. Int. J. Dairy Technol. 2006, 59, 85–96. [Google Scholar] [CrossRef]
- Schmidt, V.S.J.; Kaufmann, V.; Kulozik, U.; Scherer, S.; Wenning, M. Microbial biodiversity, quality and shelf life of microfiltered and pasteurized extended shelf life (ESL) milk from Germany, Austria and Switzerland. Int. J. Food Microbiol. 2012, 154, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Khoza, S. Effect of Extended Shelf Life Milk Processing on the Bacterial Composition Associated with the Nozzles of Filling Machines. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2015. [Google Scholar]
- Deeth, H.C.; Lewis, M.J. High Temperature Processing of Milk and Milk Products; Wiley Blackwell: Oxford, UK, 2017; ISBN 978-1-118-46050-4. [Google Scholar]
- Burton, H. Ultra High Temperature Processing of Milk and Milk Products; Elsevier Applied Science Publishers: New York, NY, USA, 1988. [Google Scholar]
- Lewis, M.; Heppell, N. Continuous Thermal Processing of Foods: Pasteurization and UHT Sterilization; Aspen Publishers: Gaithersburg, MD, USA, 2000; ISBN 0-8342-1259-5. [Google Scholar]
- Datta, N.; Elliott, A.J.; Perkins, M.L.; Deeth, H.C. Ultra-high-temperature (UHT) treatment of milk: Comparison of direct and indirect modes of heating. Aust. J. Dairy Technol. 2002, 57, 211–227. [Google Scholar]
- Fredsted, L.B.; Rysstad, G.; Eie, T. Pure-Lac™: The new milk with protected freshness and extended shelf life. In Heat Treatments and Alternative Methods; International Dairy Federation: Brussels, Belgium, 1996; Volume 9602, pp. 104–125. ISBN 92 9098 023 6. [Google Scholar]
- APV (SPX). Dairy Technology; SPX Corporation: Silkeborg, Denmark, 2008. [Google Scholar]
- Huijs, G.; van Asselt, A.; Verdurmen, R.; de Jong, P. High speed milk. Dairy Ind. Int. 2004, 69, 30–32. [Google Scholar]
- De Jong, P. Thermal processing of milk. In Advanced Dairy Science and Technology; Britz, T.J., Robinson, R.K., Eds.; Blackwell Publishers Ltd.: Oxford, UK, 2008; pp. 1–34. ISBN 978-1-4051-3618-1. [Google Scholar]
- Van Asselt, A.J.; Sweere, A.P.J.; Rollema, H.S.; de Jong, P. Extreme high-temperature treatment of milk with respect to plasmin inactivation. Int. Dairy J. 2008, 18, 531–538. [Google Scholar] [CrossRef]
- Arofikin, N.V. Liquid Product Pressure Treatment Method and Device. U.S. Patent 7,708,941, 4 May 2010. [Google Scholar]
- Myer, P.R.; Parker, K.R.; Kanach, A.T.; Zhu, T.; Morgan, M.T.; Applegate, B.M. The effect of a novel low temperature-short time (LTST) process to extend the shelf-life of fluid milk. SpringerPlus 2016, 5, 660. [Google Scholar] [CrossRef] [PubMed]
- Noma, S.; Shimoda, M.; Hayakawa, I. Inactivation of vegetative bacteria by rapid decompression treatment. J. Food Sci. 2002, 67, 3408–3411. [Google Scholar] [CrossRef]
- Hayakawa, I.; Furukawa, S.; Midzunaga, A.; Horiuchi, H.; Nakashima, T.; Fujio, Y.; Yano, Y.; Ishikura, T.; Sasaki, K. Mechanism of inactivation of heat-tolerant spores of Bacillus stearothermophilus IFO 12550 by rapid decompression. J. Food Sci. 1998, 63, 371–374. [Google Scholar] [CrossRef]
- Mayr, R.; Gutser, K.; Busse, M.; Seiler, H. Indigenous aerobic spore-formers in high heat treated (127 °C, 5 s) German ESL (Extended Shelf-life) milk. Milchwissenschaft 2004, 59, 143–146. [Google Scholar]
- Blake, M.R.; Weimer, B.C.; McMahon, D.J.; Savello, P.A. Sensory and microbial quality of milk processed for extended shelf life by direct steam injection. J. Food Prot. 1995, 58, 1007–1013. [Google Scholar] [CrossRef]
- Cromie, S.J.; Dommett, T.W.; Schmidt, D. Changes in the microflora of milk with different pasteurisation and storage conditions and aseptic packaging. Aust. J. Dairy Technol. 1989, 44, 74–77. [Google Scholar]
- Collins, E.B. Heat resistant psychrotrophic microorganisms. J. Dairy Sci. 1981, 64, 157–160. [Google Scholar] [CrossRef]
- Doll, E.V.; Scherer, S.; Wenning, M. Spoilage of microfiltered and pasteurized extended shelf life milk is mainly induced by psychrotolerant spore-forming bacteria that often originate from recontamination. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Fromm, H.I.; Boor, K. Characterization of pasteurized fluid milk shelf-life attributes. J. Food Sci. 2004, 69, 207–214. [Google Scholar] [CrossRef]
- Ranieri, M.L.; Boor, K.J. Short communication: Bacterial ecology of high-temperature, short-time pasteurized milk processed in the United States. J. Dairy Sci. 2009, 92, 4833–4840. [Google Scholar] [CrossRef] [PubMed]
- Mugadza, D.T.; Buys, E.M. Spoilage Potential of Bacillus spp. & Paenibacillus spp. in Extended Shelf Life (ESL) Milk. In Proceedings of the SASDT 48th AGM & Symposium: Innovation and Cost Optimization, Cape St Francis, Eastern Cape, South Africa, 14–16 April 2015; Available online: http://www.sasdt.co.za/wp-content/uploads/2015/04/Session-8-Speaker-26-Mugadza1.pdf (accessed on 1 October 2017).
- Martin, N.H.; Ranieri, M.L.; Murphy, S.C.; Ralyea, R.D.; Wiedmann, M.; Boor, K.J. Results from raw milk microbiological tests do not predict shelf-life performance of commercially pasteurized fluid milk. J. Dairy Sci. 2011, 94, 1211–1222. [Google Scholar] [CrossRef] [PubMed]
- Scheldeman, P.; Goossens, K.; Rodriguez-Diaz, M.; Pil, A.; Goris, J.; Herman, L.; De Vos, P.; Logan, N.A.; Heyndrickx, M. Paenibacillus lactis sp. nov. isolated from raw and heat-treated milk. Int. J. Syst. Evol. Microbiol. 2004, 54, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Bosshard, P.; Zbinden, R.; Altwegg, M. Paenibacillus turicensis sp. nov., a novel bacterium harbouring heterogeneities between 16S rRNA genes. Int. J. Syst. Evol. Microbiol. 2002, 52, 2241–2249. [Google Scholar] [CrossRef] [PubMed]
- Mugadza, D.T.; Buys, E.M. Source Tracking of Bacillus cereus in an Extended Shelf Life (ESL) Milk Processing Factory. 2017. Available online: http://sasdt.co.za/wp-content/uploads/2017/05/2017-Session-3-Speaker-3-Mugadza.pdf (accessed on 1 October 2017).
- European Food Safety Authority (EFSA). Bacillus cereus and other Bacillus spp. in foodstuffs. Opinion of the Scientific Panel on Biological Hazards on Bacillus cereus and other Bacillus spp. in foodstuffs. EFSA J. 2005, 175, 1–48. [Google Scholar] [CrossRef]
- McKillip, J.L. Prevalence and expression of enterotoxins in Bacillus cereus and other Bacillus spp., a literature review. Antonie Van Leeuwenhoek 2000, 77, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Cressey, P.; King, N.; Soboleva, T. Risk Profile: Bacillus cereus in Dairy Products. New Zealand Ministry for Primary Industries (MPI) Technical Paper, May 2016, No. 2016/58; ISBN 978-1-77665-373-7. Available online: www.mpi.govt.nz/dmsdocument/14149 (accessed on 1 October 2017).
- International Dairy Federation. Bacillus cereus in Milk and Dairy Products; International Dairy Federation: Brussels, Belgium, 1992; Volume 275. [Google Scholar]
- International Dairy Federation. Bacillus cereus in Milk and Dairy Products; International Dairy Federation: Brussels, Belgium, 2016. [Google Scholar]
- Becker, H.; Schaller, G.; von Wiese, W.; Terplan, G. Bacillus cereus in infant foods and dried milk products. Int. J. Food Microbiol. 1994, 23, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Larsen, H.D.; Jørgensen, K. Growth of Bacillus cereus in pasteurized milk products. Int. J. Food Microbiol. 1999, 46, 173–176. [Google Scholar] [CrossRef]
- Markland, S.M.; Farkas, D.F.; Kniel, K.E.; Hoover, D.G. Pathogenic psychrotolerant spore-formers: An emerging challenge for low-temperature storage of minimally processed foods. Foodborne Pathog. Dis. 2013, 10, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Ivy, R.A.; Ranieri, M.L.; Martin, N.H.; den Bakker, H.C.; Xavier, B.M.; Wiedmann, M.; Boor, K.J. Identification and characterization of psychrotolerant spore-formers associated with fluid milk production and processing. Appl. Environ. Microbiol. 2012, 78, 1853–1864. [Google Scholar] [CrossRef] [PubMed]
- Raju, V.V.R.; Chetty, M.S.; Kumar, M.K. Aerobic psychrotrophic spore forming bacteria in heat treated milk. Cheiron 1989, 18, 15–18. [Google Scholar]
- Van Netten, P.; van de Moosdijk, A.; van Hoensel, P.; Mossel, D.A.A.; Perales, I. Psychrotrophic strains of Bacillus cereus producing enterotoxin. J. Appl. Bact. 1990, 69, 73–79. [Google Scholar] [CrossRef]
- Champagne, C.P.; Laing, R.R.; Roy, D.; Mafu, A.A. Psychrotrophs in dairy products: Their effect and their control. Crit. Rev. Food Sci. Nutr. 1994, 34, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.F. Milk and dairy products. In Food Microbiology: Fundamentals and Frontiers; Doyle, M.P., Beuchat, L.R., Montville, T.J., Eds.; ASM Press: Washington, DC, USA, 1997. [Google Scholar]
- Griffiths, M.W.; Phillips, J.D. Strategies to control the outgrowth of spores of psychrotrophic Bacillus spp. in dairy-products. 1. Use of naturally-occurring materials. Milchwissenschaft 1990, 45, 621–626. [Google Scholar]
- Franklin, J.G. Some bacteriological problems in the market milk industry in the UK. J. Soc. Dairy Technol. 1969, 22, 100–112. [Google Scholar] [CrossRef]
- Coghill, D.; Juffs, H.S. Incidence of psychrotrophic spore-forming bacteria in pasteurized milk and cream products and effect of temperature on their growth. Aust. J. Dairy Technol. 1979, 34, 150–153. [Google Scholar]
- Ahmed, A.A.H.; Moustafa, M.K.; Marth, E.H. Incidence of Bacillus cereus in milk and some milk products. J. Food Prot. 1983, 46, 126–128. [Google Scholar] [CrossRef]
- Rangasamy, P.N.; Iyer, M.; Roginsky, H. Isolation and characterisation of Bacillus cereus in milk and dairy products manufactured in Victoria. Aust. J. Dairy Technol. 1993, 48, 93–95. [Google Scholar]
- Stewart, D.B. Factors influencing the incidence of B. cereus spores in milk. J. Soc. Dairy Technol. 1975, 28, 80–85. [Google Scholar] [CrossRef]
- Meer, R.R.; Baker, J.; Bodyfelt, F.W.; Griffiths, M.W. Psychrotrophic Bacillus spp. in fluid milk-products-a review. J. Food Prot. 1991, 54, 969–979. [Google Scholar] [CrossRef]
- Te Giffel, M.; Beumer, R.R.; Bonestroo, M.H.; Rombouts, F.M. Incidence and characterization of Bacillus cereus in two dairy processing plants. Neth. Milk Dairy J. 1996, 50, 479–492. [Google Scholar]
- Salustiano, V.C.; Andrade, N.J.; Soares, N.F.F.; Lima, J.C.; Bernardes, P.C.; Luiz, L.M.P.; Fernandes, P.E. Contamination of milk with Bacillus cereus by post-pasteurization surface exposure as evaluated by automated ribotyping. Food Control 2009, 20, 439–442. [Google Scholar] [CrossRef]
- Mugadza, D.T.; Buys, E.M. Diversity of Bacillus cereus strains in extended shelf life. Int. Dairy J. 2017, 73, 144–150. [Google Scholar] [CrossRef]
- Svensson, B.; Ekelund, K.; Ogura, H.; Christiansson, A. Characterisation of Bacillus cereus isolated from milk silo tanks at eight different dairy plants. Int. Dairy J. 2004, 14, 17–27. [Google Scholar] [CrossRef]
- Te Giffel, M.C.; Beumer, R.R.; Granum, P.E.; Rombouts, F.M. Isolation and characterisation of Bacillus cereus from pasteurised milk in household refrigerators in The Netherlands. Int. J. Food Microbiol. 1997, 34, 307–318. [Google Scholar] [CrossRef]
- Mugadza, D.T.; Buys, E.M. Risk Characterisation of Bacillus cereus in Extended Shelf Life (ESL) Milk. In Proceedings of the 21st SAAFoST International Congress and Exhibition, Durban, South Africa, 7–9 September 2015. [Google Scholar]
- Bartoszewiez, M.; Hansen, B.M.; Swiecicka, I. The members of the Bacillus cereus group are commonly present contaminants of fresh and heat-treated milk. Food Microbiol. 2008, 25, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Slaghuis, B.A.; Te Giffel, M.; Beumer, R.R.; Andre, G. Effect of pasturing on the incidence of Bacillus cereus spores in raw milk. Int. Dairy J. 1997, 7, 201–205. [Google Scholar] [CrossRef]
- Notermans, S.; Dufrenne, J.; Teunis, P.; Beumer, R.; Te Giffel, M.; Peeters Weem, P. A risk assessment study of Bacillus cereus present in pasteurized milk. Food Microbiol. 1997, 14, 143–151. [Google Scholar] [CrossRef]
- Agata, N.; Ohta, M.; Yokoyama, K. Production of Bacillus cereus emetic toxin (cereulide) in various foods. Int. J. Food Microbiol. 2002, 73, 23–27. [Google Scholar] [CrossRef]
- Jooste, P.J.; Anelich, L.E.C.M. Safety and quality of dairy products. In Advanced Dairy Science and Technology; Britz, T.J., Robinson, R.K., Eds.; Blackwell Publishing Professional: Oxford, UK, 2008; pp. 153–182. ISBN 9780470697634. [Google Scholar]
- Lewis, M.J. Microbiological issues associated with heat-treated milks. Int. J. Dairy Technol. 1999, 52, 121–125. [Google Scholar] [CrossRef]
- Rowan, N.J.; Anderson, J.G. Diarrhoeal enterotoxin production by psychrotrophic Bacillus cereus present in reconstituted milk-based infant formulae (MIF). Lett. Appl. Microbiol. 1998, 26, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Juffs, H.; Deeth, H.C. Scientific Evaluation of Pasteurisation for Pathogen Reduction in Milk and Milk Products; FSANZ: Canberra, Australia, 2007; ISBN 978-0-642-34555-4. [Google Scholar]
- Te Giffel, M.; Beumer, R.R.; Slaghuis, B.A.; Rombouts, F.M. Occurrence and characterization of (psychrotrophic) Bacillus cereus on farms in The Netherlands. Neth. Milk Dairy J. 1995, 49, 125–138. [Google Scholar]
- Jensen, I.; Moir, C.J. Bacillus cereus and other Bacillus species. In Foodborne Microorganisms of Public Health Significance, 6th ed.; Hocking, A.D., Ed.; Australian Institute of Food Science and Technology Inc., NSW Branch, Food Microbiology Group: Waterloo, Australia, 2003; pp. 445–478. ISBN 0957890710. [Google Scholar]
- Dufeu, J.; Leesment, H. Growth and resistance characteristics of some psychrotrophic spore-formers isolated from raw milk. In Proceedings of the XIX International Dairy Congress, New Dehli, India, 2–6 December 1974; pp. 535–536. [Google Scholar]
- Dufrenne, J.; Bijwaard, M.; Te Giffel, M.; Beumer, R.; Notermans, S. Characteristics of some psychrotrophic Bacillus cereus isolates. Int. J. Food Microbiol. 1995, 27, 175–183. [Google Scholar] [CrossRef]
- Odumeru, J.A.; Toner, A.K.; Muckle, C.A.; Griffiths, M.W.; Lynch, J.A. Detection of Bacillus cereus diarrheal enterotoxin in raw and pasteurized milk. J. Food Prot. 1997, 60, 1391–1393. [Google Scholar] [CrossRef]
- Overcast, W.W.; Atmaram, K. The role of Bacillus cereus in sweet curdling of fluid milk. J. Milk Food Technol. 1974, 37, 233–236. [Google Scholar] [CrossRef]
- Stone, J.M.; Rowlands, A. Broken or bitty cream in raw and pasteurised milk. J. Dairy Res. 1952, 19, 52–62. [Google Scholar] [CrossRef]
- Stadhouders, J.; Hup, G. Effect of milk storage before pasteurization on the formation of flocs bitty cream by Bacillus cereus in pasteurized milk. Zuivelzicht 1980, 72, 178–180. [Google Scholar]
- Mikolajcik, E.M. Thermodestruction of Bacillus spores in milk. J. Milk Food Technol. 1970, 33, 77–82. [Google Scholar] [CrossRef]
- Franklin, J.G. Spores in milk-problems associated with UHT processing. J. Appl. Bact. 1970, 33, 180–191. [Google Scholar] [CrossRef]
- Vyletelova, M.; Svec, P.; Pacova, Z.; Sedlacek, I.; Roubal, P. Occurrence of Bacillus cereus and Bacillus licheniformis strains in the course of UHT milk production. Czech J. Anim. Sci. 2002, 47, 200–205. [Google Scholar]
- Bergere, J.-L.; Cerf, O. Heat resistance of Bacillus cereus spores. In Bacillus cereus in Milk and Milk Products; International Dairy Federation: Brussels, Belgium, 1992; Volume 275, pp. 23–25. [Google Scholar]
- Bradshaw, J.G.; Peeler, J.T.; Twedt, R.M. Heat-resistance of ileal loop reactive Bacillus cereus strains isolated from commercially canned food. Appl. Microbiol. 1975, 30, 943–945. [Google Scholar] [PubMed]
- Stoeckel, M.; Westermann, A.C.; Atamer, Z.; Hinrichs, J. Thermal inactivation of Bacillus cereus spores in infant formula under shear conditions. Dairy Sci. Technol. 2013, 93, 163–175. [Google Scholar] [CrossRef]
- Van Asselt, E.D.; Zwietering, M.H. A systematic approach to determine global thermal inactivation parameters for various food pathogens. Int. J. Food Microbiol. 2006, 107, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, J.; Atamer, Z. Sterilization of milk and other products. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Fox, P.F., Eds.; McSweeney Academic Press: San Diego, CA, USA, 2011; Volume 2, pp. 714–724. ISBN 978-0-12-374402-9. [Google Scholar]
- Van Asselt, A.J.; Te Giffel, M.C. Hygienic practices in liquid milk dairies. In Market Milks–Processing and Quality Management; Tamime, A.Y., Ed.; Blackwell Publishing: Oxford, UK, 2009; pp. 48–71. [Google Scholar]
- Simmonds, P.; Mossel, B.L.; Intaraphan, T.; Deeth, H.C. The heat resistance of Bacillus spores when adhered to stainless steel and its relationship to spore hydrophobicity. J. Food Prot. 2003, 66, 2070–2075. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, J.; Kessler, H. Heat resistance of Bacillus cereus spores located between seals and seal surfaces. J. Food Prot. 1995, 58, 1206–1210. [Google Scholar] [CrossRef]
- Bergere, J.-L. Spore formation and germination of Bacillus cereus: The spore cycle. In Bacillus cereus in Milk and Milk Products; International Dairy Federation: Brussels, Belgium, 1992; Volume 275, pp. 9–14. [Google Scholar]
- Stadhouders, J.; Hup, G.; Langeveld, L.P.M. Some observations on the germination, heat resistance and outgrowth of fast-germinating and slow-germinating spores of Bacillus cereus in pasteurized milk. Neth. Milk Dairy J. 1980, 34, 215–228. [Google Scholar]
- Guirguis, A.H.; Griffiths, M.W.; Muir, D.D. Spore-forming bacteria in milk. 1. Optimization of heat-treatment for activation of spores of Bacillus species. Milchwissenschaft 1983, 38, 641–644. [Google Scholar]
- Wilkinson, G.; Davies, F.L. Germination of spores of Bacillus cereus in milk and milk dialysates: Effect of heat treatment. J. Appl. Bact. 1973, 36, 485–496. [Google Scholar] [CrossRef]
- Labots, H.; Hup, G. Bacillus cereus in raw and pasteurized milk. II. The occurrence of slow and fast germinating spores in milk and their significance in the enumeration of B. cereus spores. Neth. Milk Dairy J. 1964, 18, 167–176. [Google Scholar]
- Anderson Borge, G.I.; Skeie, M.; Sorhaug, T.; Langsrud, T.; Granum, P.E. Growth and toxin profiles of Bacillus cereus isolated from different food sources. Int. J. Food Microbiol. 2001, 69, 237–246. [Google Scholar] [CrossRef]
- Griffiths, M.W.; Phillips, J.D. Modeling the relation between bacterial-growth and storage-temperature in pasteurized milks of varying hygienic quality. J. Soc. Dairy Technol. 1988, 41, 96–102. [Google Scholar] [CrossRef]
- Mayr, R.; Gutser, K.; Busse, M.; Seiler, H. Gram-positive non-spore-forming bacteria are frequent spoilage organisms of German retail ESL Extended Shelf-life milk. Milchwissenschaft 2004, 59, 262–266. [Google Scholar]
- Mugadza, D.; Buys, E. Diversity of Spore Former and Non-Spore Former Bacteria in Extended Shelf-Life (ESL) Milk; South African Society of Dairy Technology Dairy Symposium: Capetown, South Africa, 2014. [Google Scholar]
- Griffiths, M.W.; Phillips, J.D. The application of the preincubation test in commercial dairies. Aust. J. Dairy Technol. 1986, 41, 71–79. [Google Scholar]
- Craven, H.M.; Macauley, B.J. Micro-organisms in pasteurised milk after refrigerated storage. 3. Effect of milk processors. Aust. J. Dairy Technol. 1992, 47, 50–55. [Google Scholar]
- Brody, A.L. Aseptic and extended-shelf-life packaging. Food Technol. 2006, 60, 66–68. [Google Scholar]
- Brody, A.L. The when and why of aseptic packaging. Food Technol. 2000, 54, 101–102. [Google Scholar]
- Mayer, H.K.; Raba, B.; Meier, J.; Schmid, A. RP-HPLC analysis of furosine and acid-soluble β-lactoglobulin to assess the heat load of extended shelf life milk samples in Austria. Dairy Sci. Technol. 2010, 90, 413–428. [Google Scholar] [CrossRef]
- Gaafar, A.M.M. Investigation into the Cooked Flavor in Heat-Treated Milk. Ph.D. Thesis, University of Reading, Reading, UK, 1987. [Google Scholar]
- Oliveira, L.N.; Marinho, V.T.; Zamagno, M.V.; Lauro, M.A.; Barboza, J.A.N.; da Silva, P.H.F. Assessment of whey protein nitrogen index as an indicator of heat treatment for UHT milk and milk powder. J. Candido Tostes Dairy Inst. 2015, 70, 78–84. [Google Scholar] [CrossRef]
- Anema, S.G.; Lloyd, R.J. Analysis of whey protein denaturation: A comparative study of alternative methods. Milchwissenschaft 1999, 54, 206–210. [Google Scholar]
- Lorenzen, P.C.; Clawin-Raedecker, I.; Einhoff, K.; Hammer, P.; Hartmann, R.; Hoffmann, W.; Martin, D.; Molkentin, J.; Walte, H.G.; Devrese, M. A survey of the quality of extended shelf life (ESL) milk in relation to HTST and UHT milk. Int. J. Dairy Technol. 2011, 64, 166–178. [Google Scholar] [CrossRef]
- Gallmann, P.U. Possible quality improvements/benefits of extended shelf life (ESL) milk. In Proceedings of the Conference on Extended Shelf-Life Milk; International Dairy Federation: Brussels, Belgium, 2000. [Google Scholar]
- Kaufmann, V.; Scherer, S.; Kulozik, U. Procedures for the prolongation of the storability of consumable milk and its material changes: ESL milk. J. Verbraucherschutz Lebensm.-J. Consum. Prot. Food Saf. 2010, 5, 59–64. [Google Scholar] [CrossRef]
- Vatne, K.B.; Castberg, H.B. Processing and packaging aspects of extended shelf life products. Aust. J. Dairy Technol. 1991, 46, 98–100. [Google Scholar]
- Kaufmann, V.; Kulozik, U. Processing factors influencing quality and stability of extended shelf life (ESL) milk. DMZ Lebensm. Milchmirtschaft 2008, 129, 28–30. [Google Scholar] [CrossRef]
- MGT. ESL Systems. Extended Shelf Life Milk (Brochure). Available online: http://www.mgt.co.il/sites/MGT_site/UserContent/files/MGT_ESL_brochure_V9.pdf (accessed on 1 October 2017).
- Manji, B. Regulatory perspectives to ESL products: North American situation. In Proceedings of the Conference on Extended Shelf-Life Milk; International Dairy Federation: Brussels, Belgium, 2000. [Google Scholar]
- Ranjith, H.M.P. High temperature pasteurization. In Proceedings of the Conference on Extended Shelf-Life Milk; International Dairy Federation: Brussels, Belgium, 2000. [Google Scholar]
- Lyster, R.L.J. The denaturation of α-lactalbumin and β-lactoglobulin in heated milk. J. Dairy Res. 1970, 37, 233–243. [Google Scholar] [CrossRef]
- Crudden, A.; Fox, F.; Kelly, A.L. Factors affecting the hydrolytic action of plasmin in milk. Int. Dairy J. 2005, 15, 305–313. [Google Scholar] [CrossRef]
- Browning, E.; Lewis, M.; MacDougall, D. Predicting safety and quality parameters for UHT-processed milks. Int. J. Dairy Technol. 2001, 54, 111–120. [Google Scholar] [CrossRef]
- Tran, H.; Datta, N.; Lewis, M.J.; Deeth, H.C. Predictions of some product parameters based on the processing conditions of ultra-high-temperature milk plants. Int. Dairy J. 2008, 18, 939–944. [Google Scholar] [CrossRef]
- Kessler, H.G. Food Engineering and Dairy Technology; Verlag A. Kessler: Freising, Germany, 1981. [Google Scholar]
- Chavan, R.S.; Chavan, S.R.; Khedkar, C.D.; Jana, A.H. UHT milk processing and effect of plasmin activity on shelf-life: A review. Comp. Rev. Food Sci. Food Saf. 2011, 10, 251–268. [Google Scholar] [CrossRef]

{kind=link}
| Heating Conditions (°C/s) | B* | β-Lactoglobulin Denaturation (%) 2 | Comments |
|---|---|---|---|
| 120/9 | 0.03 | 61 | B* too low to inactivate spores of psychrotrophic bacteria; β-Lg denaturation too high |
| 127/5 | 0.09 | 55 | Representative of commonly used conditions for ESL milk; B* too low to inactivate spores of psychrotrophic bacteria; β-Lg denaturation marginal |
| 134/4 | 0.32 | 56 | Conditions sufficient for inactivating spores of psychrotrophic bacteria; β-Lg denaturation marginal |
| 138/2 | 0.40 | 45 | Minimum conditions for ESL in USA; meets proposed criteria |
| 140/1 | 0.32 | 34 | Meets proposed criteria; excellent conditions if short holding time can be achieved |
| 145/0.3 | 0.32 | 24 | Meets proposed criteria; excellent conditions if short holding time can be achieved |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deeth, H. Optimum Thermal Processing for Extended Shelf-Life (ESL) Milk. Foods 2017, 6, 102. https://doi.org/10.3390/foods6110102
Deeth H. Optimum Thermal Processing for Extended Shelf-Life (ESL) Milk. Foods. 2017; 6(11):102. https://doi.org/10.3390/foods6110102
Chicago/Turabian StyleDeeth, Hilton. 2017. "Optimum Thermal Processing for Extended Shelf-Life (ESL) Milk" Foods 6, no. 11: 102. https://doi.org/10.3390/foods6110102
APA StyleDeeth, H. (2017). Optimum Thermal Processing for Extended Shelf-Life (ESL) Milk. Foods, 6(11), 102. https://doi.org/10.3390/foods6110102