
Fermentation-Assisted Extraction of Isothiocyanates from Brassica Vegetable Using Box-Behnken Experimental Design



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Their Preparation
2.2. Culture
2.3. Inoculum Preparation
2.4. De-Sulphatation of GLS
2.5. Identification and Quantification of GLS
2.6. Estimation of Total Isothiocyanates Content
2.7. Box-Behnken Design Experiments
2.8. Optimisation of LAB Fermentation for Maximum ITCs Yield
2.9. Viable Cell Counts
2.10. Statistical Analysis
3. Results
3.1. Identification of GLS in Irish York Cabbage
3.2. Effect of Fermentation on GLS
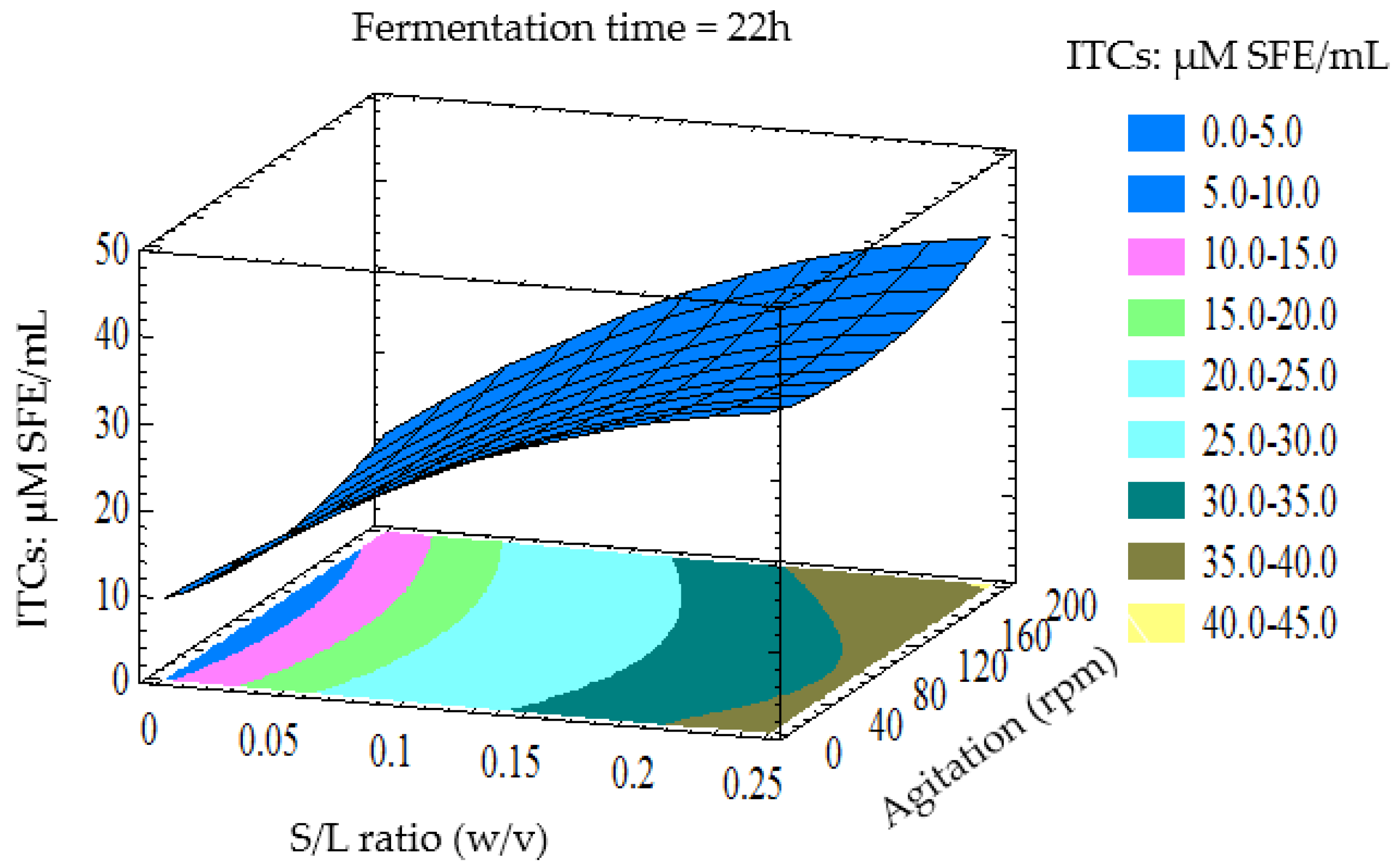
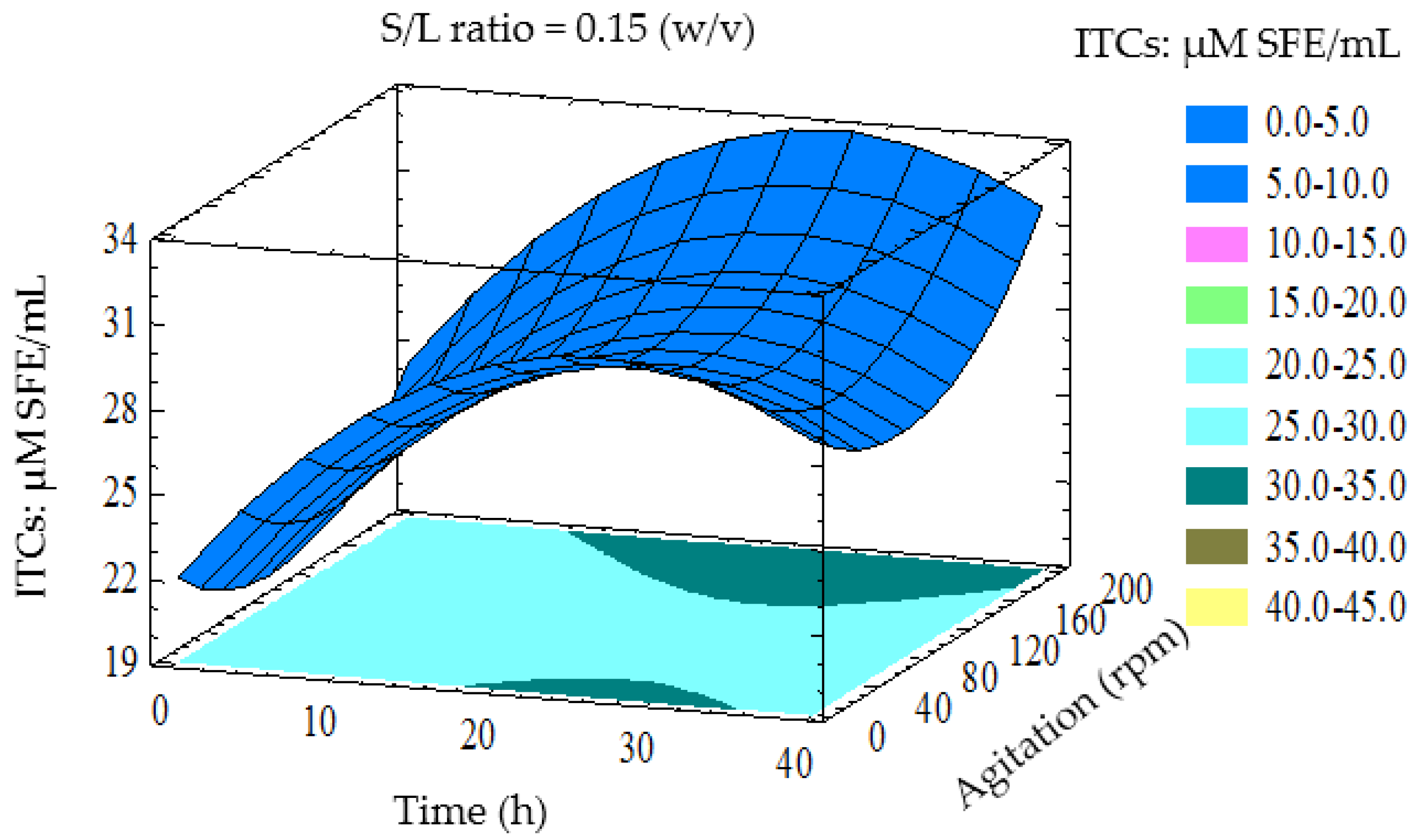
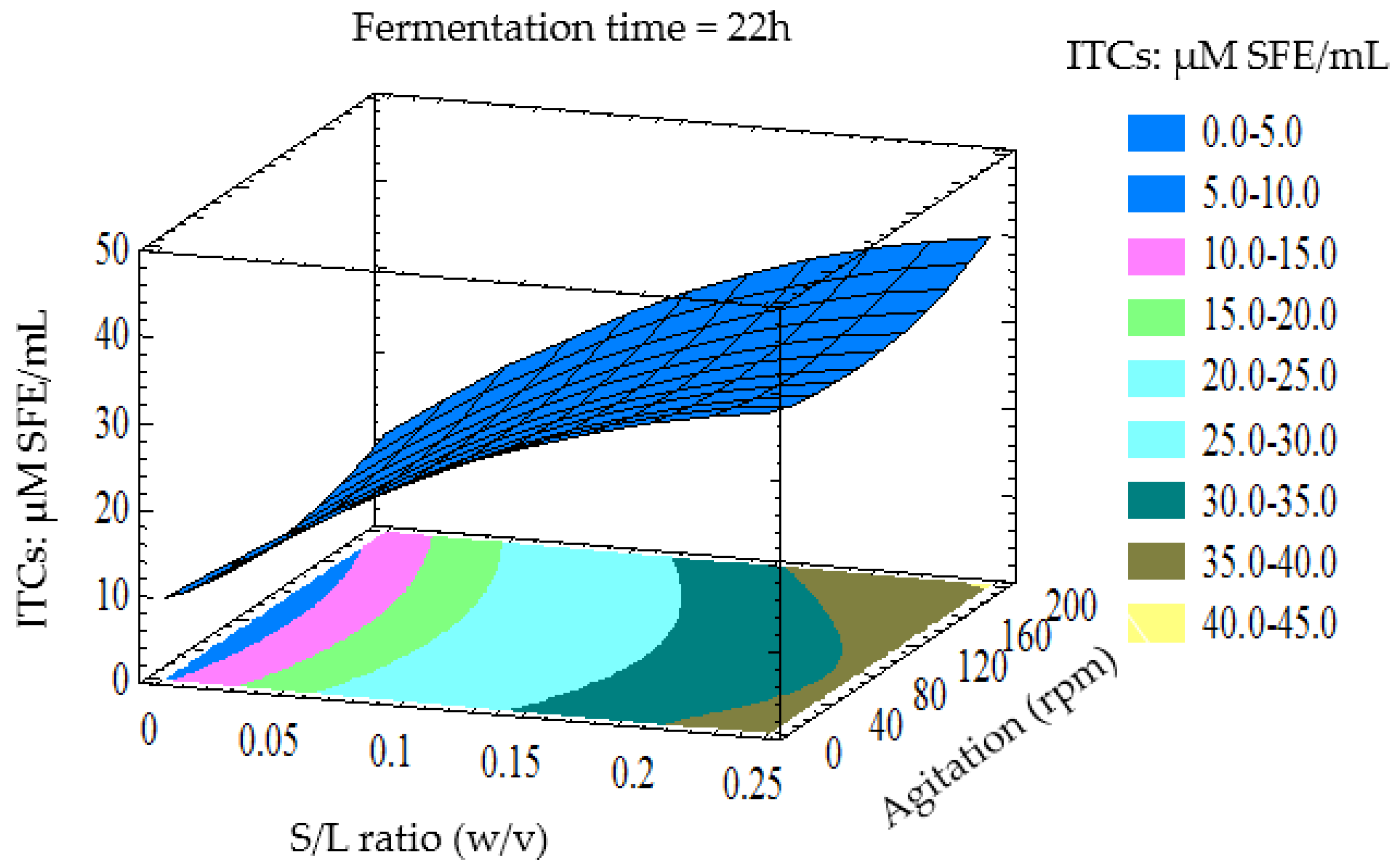
3.3. Optimisation of LAB Fermentation for High ITCs Yield
3.4. Verification of Predictive Model
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dufour, V.; Stahl, M.; Baysse, C. The antibacterial properties of isothiocyanates. Microbiology 2015, 161, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Kadir, N.H.; David, R.; Rossiter, J.T.; Gooderham, N.J. The selective cytotoxicity of the alkenyl glucosinolate hydrolysis products and their presence in brassica vegetables. Toxicology 2015, 334, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.K.; Lund, E.K.; Parker, M.L.; Clarke, R.G.; Johnson, I.T. Allyl-isothiocyanate causes mitotic block, loss of cell adhesion and disrupted cytoskeletal structure in ht29 cells. Carcinogenesis 2004, 25, 1409–1415. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Srivastava, S.K.; Lew, K.L.; Zeng, Y.; Hershberger, P.; Johnson, C.S.; Trump, D.L.; Singh, S.V. Allyl isothiocyanate, a constituent of cruciferous vegetables, inhibits proliferation of human prostate cancer cells by causing g2/m arrest and inducing apoptosis. Carcinogenesis 2003, 24, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Okulicz, M. Multidirectional time-dependent effect of sinigrin and allyl isothiocyanate on metabolic parameters in rats. Plant Foods Hum. Nutr. 2010, 65, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Hong, E.; Kim, G.H. Evaluation of antibacterial activity of 3-butenyl, 4-pentenyl, 2-phenylethyl, and benzyl isothiocyanate in brassica vegetables. J. Food Sci. 2010, 75, M412–M416. [Google Scholar] [CrossRef] [PubMed]
- Kurt, S.; Günes, U.; Soylu, E.M. In Vitro and In Vivo antifungal activity of synthetic pure isothiocyanates against sclerotinia sclerotiorum. Pest Manag. Sci. 2011, 67, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Manyes, L.; Luciano, F.; Mañes, J.; Meca, G. In Vitro antifungal activity of allyl isothiocyanate (aitc) against aspergillus parasiticus and penicillium expansum and evaluation of the aitc estimated daily intake. Food Chem. Toxicol. 2015, 83, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Langer, P.; Stolc, V. Goitrogenic activity of allylisothiocyanate—A widespread natural mustard oil. Endocrinology 1965, 76, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Talalay, P.; De Long, M.J.; Prochaska, H.J. Identification of a common chemical signal regulating the induction of enzymes that protect against chemical carcinogenesis. Proc. Natl. Acad. Sci. USA 1988, 85, 8261–8265. [Google Scholar] [CrossRef] [PubMed]
- Ippoushi, K.; Takeuchi, A.; Azuma, K. Sinigrin suppresses nitric oxide production in rats administered intraperitoneally with lipopolysaccharide. Food Chem. 2010, 120, 1119–1121. [Google Scholar] [CrossRef]
- Aguilar, F.; Dusemund, B.; Galtier, P.; Gilbert, J.; Gott, D.; Grilli, S.; Gürtler, R.; König, J.; Lambré, C.; Larsen, J. Efsa panel on food additives and nutrient sources added to food (ans): Scientific opinion on the safety of allyl isothiocyanate for the proposed uses as a food additive. EFSA J. 2010, 8, 1943–1983. [Google Scholar]
- Banerjee, A.; Penna, S.; Variyar, P.S. Allyl isothiocyanate enhances shelf life of minimally processed shredded cabbage. Food Chem. 2015, 183, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [PubMed]
- Nugrahedi, P.Y.; Verkerk, R.; Widianarko, B.; Dekker, M. A mechanistic perspective on process-induced changes in glucosinolate content in brassica vegetables: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 823–838. [Google Scholar] [CrossRef] [PubMed]
- Peñas, E.; Pihlava, J.-M.; Vidal-Valverde, C.; Frías, J. Influence of fermentation conditions of Brassica oleracea L. var. Capitata on the volatile glucosinolate hydrolysis compounds of sauerkrauts. LWT-Food Sci. Technol. 2012, 48, 16–23. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Meehan, J.T., IV. Use of isothiocyanates for suppression of palmer amaranth (Amaranthus palmeri), pitted morningglory (Ipomoea lacunosa), and yellow nutsedge (Cyperus esculentus). Weed Sci. 2005, 53, 884–890. [Google Scholar] [CrossRef]
- Guo, Q.; Guo, L.; Wang, Z.; Zhuang, Y.; Gu, Z. Response surface optimization and identification of isothiocyanates produced from broccoli sprouts. Food Chem. 2013, 141, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lee, W.; Lee, W.J.; Auh, J.H.; Kim, S.S.; Yoon, J. Extraction of allyl isothiocyanate from wasabi (Wasabia japonica matsum) using supercritical carbon dioxide. Food Sci. Biotechnol. 2010, 19, 405–410. [Google Scholar] [CrossRef]
- Parniakov, O.; Roselló-Soto, E.; Barba, F.J.; Grimi, N.; Lebovka, N.; Vorobiev, E. New approaches for the effective valorization of papaya seeds: Extraction of proteins, phenolic compounds, carbohydrates, and isothiocyanates assisted by pulsed electric energy. Food Res. Int. 2015, 77, 711–717. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Peñas, E.; Frias, J.; Ciska, E.; Honke, J.; Piskula, M.; Kozlowska, H.; Vidal-Valverde, C. Influence of fermentation conditions on glucosinolates, ascorbigen, and ascorbic acid content in white cabbage (Brassica oleracea var. Capitata cv. Taler) cultivated in different seasons. J. Food Sci. 2009, 74, C62–C67. [Google Scholar] [PubMed]
- Palani, K.; Harbaum-Piayda, B.; Meske, D.; Keppler, J.K.; Bockelmann, W.; Heller, K.J.; Schwarz, K. Influence of fermentation on glucosinolates and glucobrassicin degradation products in sauerkraut. Food Chem. 2016, 190, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cho, C.-G.; Posner, G.H.; Talalay, P. Spectroscopic quantitation of organic isothiocyanates by cyclocondensation with vicinal dithiols. Anal. Biochem. 1992, 205, 100–107. [Google Scholar] [CrossRef]
- Kusznierewicz, B.; Bartoszek, A.; Wolska, L.; Drzewiecki, J.; Gorinstein, S.; Namieśnik, J. Partial characterization of white cabbages (Brassica oleracea var. Capitata F. Alba) from different regions by glucosinolates, bioactive compounds, total antioxidant activities and proteins. LWT-Food Sci. Technol. 2008, 41, 1–9. [Google Scholar]
- Ciska, E.; Pathak, D.R. Glucosinolate derivatives in stored fermented cabbage. J. Agric. Food Chem. 2004, 52, 7938–7943. [Google Scholar] [CrossRef] [PubMed]
- Cartea, M.E.; Velasco, P. Glucosinolates in brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Aires, A.; Fernandes, C.; Carvalho, R.; Bennett, R.N.; Saavedra, M.J.; Rosa, E.A. Seasonal effects on bioactive compounds and antioxidant capacity of six economically important brassica vegetables. Molecules 2011, 16, 6816–6832. [Google Scholar] [CrossRef] [PubMed]
- Ludikhuyze, L.; Ooms, V.; Weemaes, C.; Hendrickx, M. Kinetic study of the irreversible thermal and pressure inactivation of myrosinase from broccoli (Brassica oleracea L. cv. Italica). J. Agric. Food Chem. 1999, 47, 1794–1800. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.-C.; Wei, Q.-K. Myrosinase activity and total glucosinolate content of cruciferous vegetables, and some properties of cabbage myrosinase in Taiwan. J. Sci. Food Agric. 1993, 61, 471–475. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Gupta, S.; Abu-Ghannam, N. Kinetic evaluation of colour, texture, polyphenols and antioxidant capacity of irish york cabbage after blanching treatment. Food Chem. 2012, 131, 63–72. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Gupta, S.; Abu-Ghannam, N. Optimisation of lactic acid fermentation of york cabbage for the development of potential probiotic products. Int. J. Food Sci. Technol. 2012, 47, 1605–1612. [Google Scholar] [CrossRef]
- Pirzadah, T.; Garg, S.; Singh, J.; Vyas, A.; Kumar, M.; Gaur, N.; Bala, M.; Rehman, R.; Varma, A.; Kumar, V.; et al. Characterization of actinomycetes and trichoderma spp. for cellulase production utilizing crude substrates by response surface methodology. SpringerPlus 2014, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tolonen, M.; Taipale, M.; Viander, B.; Pihlava, J.-M.; Korhonen, H.; Ryhänen, E.-L. Plant-derived biomolecules in fermented cabbage. J. Agric. Food Chem. 2002, 50, 6798–6803. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Independent Variables | Coded Symbols | Coded Variable Level | ||
|---|---|---|---|---|
| –1 | 0 | +1 | ||
| S/L ratio (w/v) | X1 | 0.05 | 0.15 | 0.25 |
| Fermentation time (h) | X2 | 8 | 22 | 36 |
| Agitation rate (rpm) | X3 | 0 | 100 | 200 |
| Individual GLS | GLS Content (μg/mL) |
|---|---|
| Aliphatic GLS | |
| Sinigrin | 1.96 ± 0.025 |
| Gluconapin | 1.78 ± 0.075 |
| Glucoiberin | 3.08 ± 0.070 |
| Glucoraphanin | 3.92 ± 0.178 |
| Indol GLS | |
| Glucobrassicin | 2.42 ± 0.047 |
| Neoglucobrassicin | 0.63 ± 0.085 |
| 4-Methoxyglucobrassicin | 0.27 ± 0.038 |
| Source | Sum of Squares | Df | Mean Square | F-Ratio | p-Value |
|---|---|---|---|---|---|
| A: S/L ratio | 764.2 | 1 | 764.2 | 2496.9 | 0.0000 |
| B: Time | 24.2 | 1 | 24.2 | 79.0 | 0.0009 |
| C: Agitation | 15.6 | 1 | 15.6 | 50.9 | 0.0020 |
| AA | 41.9 | 1 | 41.9 | 137.0 | 0.0003 |
| AB | 42.4 | 1 | 42.4 | 138.5 | 0.0003 |
| AC | 0.34 | 1 | 0.34 | 1.12 | 0.3500 |
| BB | 26.3 | 1 | 26.3 | 86.1 | 0.0008 |
| BC | 0.05 | 1 | 0.05 | 0.17 | 0.7051 |
| CC | 40.2 | 1 | 40.2 | 131.5 | 0.0003 |
| Lack-of-fit | 17.1 | 3 | 5.69 | 18.6 | 0.0082 |
| Pure error | 1.22 | 4 | 0.31 | ||
| Total (corr.) | 969.7 | 16 |
| Row | X1 | X2 | X3 | Total ITCs Content | |
|---|---|---|---|---|---|
| (w/v) | (h) | (rpm) | Exp | Predicted | |
| 1 | 0.25 | 36 | 100 | 36.8 | 38.1 |
| 2 | 0.25 | 22 | 200 | 39.7 | 40.4 |
| 3 | 0.15 | 8 | 200 | 28.9 | 29.2 |
| 4 | 0.15 | 22 | 100 | 28.7 | 29.0 |
| 5 | 0.15 | 8 | 0 | 24.6 | 26.6 |
| 6 | 0.05 | 22 | 200 | 19.2 | 20.3 |
| 7 | 0.25 | 22 | 0 | 38.1 | 37.0 |
| 8 | 0.15 | 22 | 100 | 28.3 | 29.0 |
| 9 | 0.15 | 22 | 100 | 29.2 | 29.0 |
| 10 | 0.05 | 8 | 100 | 16.4 | 15.1 |
| 11 | 0.15 | 22 | 100 | 29.1 | 29.0 |
| 12 | 0.15 | 36 | 0 | 30.1 | 29.8 |
| 13 | 0.25 | 8 | 100 | 29.1 | 28.1 |
| 14 | 0.05 | 36 | 100 | 11.1 | 12.1 |
| 15 | 0.15 | 36 | 200 | 34.9 | 32.8 |
| 16 | 0.05 | 22 | 0 | 18.8 | 18.1 |
| 17 | 0.15 | 22 | 100 | 29.8 | 29.0 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaiswal, A.K.; Abu-Ghannam, N. Fermentation-Assisted Extraction of Isothiocyanates from Brassica Vegetable Using Box-Behnken Experimental Design. Foods 2016, 5, 75. https://doi.org/10.3390/foods5040075
Jaiswal AK, Abu-Ghannam N. Fermentation-Assisted Extraction of Isothiocyanates from Brassica Vegetable Using Box-Behnken Experimental Design. Foods. 2016; 5(4):75. https://doi.org/10.3390/foods5040075
Chicago/Turabian StyleJaiswal, Amit K., and Nissreen Abu-Ghannam. 2016. "Fermentation-Assisted Extraction of Isothiocyanates from Brassica Vegetable Using Box-Behnken Experimental Design" Foods 5, no. 4: 75. https://doi.org/10.3390/foods5040075
APA StyleJaiswal, A. K., & Abu-Ghannam, N. (2016). Fermentation-Assisted Extraction of Isothiocyanates from Brassica Vegetable Using Box-Behnken Experimental Design. Foods, 5(4), 75. https://doi.org/10.3390/foods5040075