
The Effect of Lactic Acid Bacteria Fermentation on the Anti-Diabetic Activity of Pumpkin Puree

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
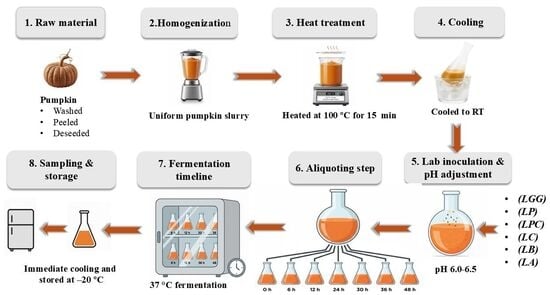
2.2. Preparation of Pumpkin Puree and Inoculated Fermentation
2.3. Determination of Viable LAB Counts
2.4. Physicochemical Analyses
2.4.1. pH Measurement
2.4.2. Total Titratable Acidity
2.4.3. Determination of Reducing Sugar
2.4.4. Determination of Total Soluble Sugars
2.5. Determination of Bioactive Compounds
2.5.1. Total Phenolic Content (TPC)
2.5.2. HPLC Analysis of Phenolic Compounds
2.6. Antioxidant Activities
2.6.1. DPPH Radical Scavenging Activity
2.6.2. ABTS Radical Scavenging Activity
2.7. Enzyme Inhibition Assays
2.7.1. α-Glucosidase Inhibition Assay
2.7.2. α-Amylase Inhibition Assay
2.8. Cell Culture and Glucose Uptake Measurement
2.9. Statistical Analysis
3. Results and Discussion
3.1. Phsiochemical Parameters and Viable Counts
3.2. Total Soluble Sugars
3.3. Bioactive Compounds
3.4. Antioxidant properties
3.5. α-Glucosidase and α-Amylase Inhibition
3.6. Cell Culture Glucose Uptake
3.7. Heat Map
3.8. Pearson’s Correlation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- Koh, W.; Uthumporn, U.; Rosma, A.; Irfan, A.; Park, Y. Optimization of a fermented pumpkin-based beverage to improve Lactobacillus mali survival and α-glucosidase inhibitory activity: A response surface methodology approach. Food Sci. Hum. Wellness 2018, 7, 57–70. [Google Scholar] [CrossRef]
- Bengtsson, J.; Rieckmann, A.; Carstensen, B.; Svensson, J.; Jørgensen, M.E.; Rod, N.H. Trajectories of childhood adversity and type 1 diabetes: A nationwide study of one million children. Diabetes Care 2021, 44, 740–747. [Google Scholar] [CrossRef]
- Dayan, C.M.; Besser, R.E.J.; Oram, R.A.; Hagopian, W.; Vatish, M.; Bendor-Samuel, O.; Snape, M.D.; Todd, J.A. Preventing type 1 diabetes in childhood. Science 2021, 373, 506–510. [Google Scholar] [CrossRef]
- Isara, R.D.S.; Gunathilaka, M.D.T.L. Pumpkin seeds and leaves as an alternative medicine for the treatment of hyperglycemia. Univ. Colombo Rev. 2023, 4, 31–47. [Google Scholar] [CrossRef]
- Kooti, W.; Farokhipour, M.; Asadzadeh, Z.; Ashtary-Larky, D.; Asadi-Samani, M. The role of medicinal plants in the treatment of diabetes: A systematic review. Electron. Physician 2016, 8, 1832–1842. [Google Scholar] [CrossRef] [PubMed]
- Kulczyński, B.; Gramza-Michałowska, A. The profile of secondary metabolites and other bioactive compounds in Cucurbita pepo L. and Cucurbita moschata pumpkin cultivars. Molecules 2019, 24, 2945. [Google Scholar] [CrossRef]
- Frattaruolo, L.; Durante, M.; Cappello, M.S.; Montefusco, A.; Mita, G.; Cappello, A.R.; Lenucci, M.S. The ability of supercritical CO2 carrot and pumpkin extracts to counteract inflammation and oxidative stress in RAW 264.7 macrophages stimulated with LPS or MDA-MB-231 cell-conditioned media. Food Funct. 2023, 14, 10083–10096. [Google Scholar] [CrossRef]
- Gebhardt, S.E.; Lemar, L.E.; Haytowitz, D.B.; Pehrsson, P.R.; Nickle, M.S.; Showell, B.A.; Thomas, R.G.; Exler, J.; Holden, J.M. USDA National Nutrient Database for Standard Reference, Release 21; United States Department of Agriculture (USDA), Agricultural Research Service, Beltsville Human Nutrition Research Center: Beltsville, MD, USA, 2008. [Google Scholar]
- Zhou, C.-L.; Liu, W.; Zhao, J.; Yuan, C.; Song, Y.; Chen, D.; Ni, Y.-Y.; Li, Q.-H. The effect of high hydrostatic pressure on the microbiological quality and physical–chemical characteristics of Pumpkin (Cucurbita maxima Duch.) during refrigerated storage. Innov. Food Sci. Emerg. Technol. 2014, 21, 24–34. [Google Scholar] [CrossRef]
- Sedigheh, A.; Jamal, M.S.; Mahbubeh, S.; Somayeh, K.; Mahmoud, R.; Azadeh, A.; Fatemeh, S. Hypoglycaemic and hypolipidemic effects of pumpkin (Cucurbita pepo L.) on alloxan-induced diabetic rats. Afr. J. Pharm. Pharmacol. 2011, 5, 2620–2626. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Radziejewska-Kubzdela, E.; Kidoń, M.; Kowiel, A.; Waszkowiak, K.; Szymandera-Buszka, K.; Bednarek, M.; Kuligowski, M.; Kobus-Cisowska, J.; Mierzwa, D. The effect of ultrasound and lactic acid fermentation on the selected quality parameters and bioactive compounds content in fermented pumpkin (Cucurbita pepo L.). Molecules 2024, 29, 5586. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Vincentini, O.; Cantatore, V.; Cavoski, I.; Gobbetti, M. Fermented Portulaca oleracea L. juice: A novel functional beverage with potential ameliorating effects on the intestinal inflammation and epithelial injury. Nutrients 2019, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, Y.; Li, F.; Jiao, X.; Ma, D.; Zhang, L.; Yang, B.; Zhao, J.; Han, J.; Li, Q. Effects of lactic acid bacteria fermentation on chemical compounds, antioxidant capacities and hypoglycemic properties of pumpkin juice. Food Biosci. 2022, 50, 102126. [Google Scholar] [CrossRef]
- Sammulia, S.; Suhaera, S.; Prayoga, D.; Pitriani, P.; Ramadhania, Z.; Fristiohady, A.; Milanda, T. Fermentation-induced changes in phytochemical composition and pharmacological activities of zingiberaceae plants: Insight from in vitro and in vivo studies. Drug Des. Dev. Ther. 2026, 20, 604038. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Ndeye, A.N.; Bassama, J.; Dieng, M.; Touré-Kané, N.C.; Montet, D. Optimization scale pasteurization of baobab juice using Response Surface Methodology (RSM). Food Nutr. Sci. 2020, 11, 113–122. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Quan, Q.; Liu, W.; Guo, J.; Ye, M.; Zhang, J. Effect of six lactic acid bacteria strains on physicochemical characteristics, antioxidant activities and sensory properties of fermented orange juices. Foods 2022, 11, 1920. [Google Scholar] [CrossRef]
- Lamuela-Raventós, R.M. Folin-Ciocalteu method for the measurement of total phenolic content and antioxidant capacity. In Measurement of Antioxidant Activity & Capacity: Recent Trends and Applications; Apak, R., Çapanoğlu, E., Shahidi, F., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 107–115. [Google Scholar] [CrossRef]
- Liu, Y.; Sheng, J.; Li, J.; Zhang, P.; Tang, F.; Shan, C. Influence of lactic acid bacteria on physicochemical indexes, sensory and flavor characteristics of fermented sea buckthorn juice. Food Biosci. 2022, 46, 101519. [Google Scholar] [CrossRef]
- Yue, Q.; Wang, Z.; Yu, F.; Tang, X.; Su, L.; Zhang, S.; Sun, X.; Li, K.; Zhao, C.; Zhao, L. Changes in metabolite profiles and antioxidant and hypoglycemic activities of Laminaria japonica after fermentation. LWT—Food Sci. Technol. 2022, 158, 113122. [Google Scholar] [CrossRef]
- Huang, C.; Somwar, R.; Patel, N.; Niu, W.; Torok, D.; Klip, A. Sustained exposure of L6 myotubes to high glucose and insulin decreases insulin-stimulated GLUT4 translocation but upregulates GLUT4 activity. Diabetes 2002, 51, 2090–2098. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Zou, C.; Wang, Y.; Shen, Z. 2-NBDG as a fluorescent indicator for direct glucose uptake measurement. J. Biochem. Biophys. Methods 2005, 64, 207–215. [Google Scholar] [CrossRef]
- Tkacz, K.; Chmielewska, J.; Turkiewicz, I.P.; Nowicka, P.; Wojdyło, A. Dynamics of changes in organic acids, sugars and phenolic compounds and antioxidant activity of sea buckthorn and sea buckthorn-apple juices during Malolactic fermentation. Food Chem. 2020, 332, 127382. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hu, Y.; Xie, Y.; Cui, M.; Jiang, J.; Li, Y.; Guo, J.; Xiang, M.; Tian, Z.; Niu, J.; et al. A comprehensive review of lactic acid bacteria-fermented plant-based beverages: Substrate types, nutritional quality, probiotic characteristics, metabolic pathways and future development trends. Crit. Rev. Food Sci. Nutr. 2025, 1–29. [Google Scholar] [CrossRef]
- Markkinen, N.; Laaksonen, O.; Nahku, R.; Kuldjärv, R.; Yang, B. Impact of lactic acid fermentation on acids, sugars, and phenolic compounds in black chokeberry and sea buckthorn juices. Food Chem. 2019, 286, 204–215. [Google Scholar] [CrossRef]
- Borowska, M.; Buttimer, C.; Bottacini, F.; Arendt, E.K.; Coffey, A. Comparative genomic analysis of selected lactic acid bacteria and phenotypic association of the key genes involved in fructan and ribose utilisation. Discov. Bact. 2025, 2, 5. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, H.; Lei, H. Phenolics profile, antioxidant activity and flavor volatiles of pear juice: Influence of lactic acid fermentation using three lactobacillus strains in monoculture and binary mixture. Foods 2022, 11, 11. [Google Scholar] [CrossRef]
- Tamang, J.P.; Shin, D.-H.; Jung, S.-J.; Chae, S.-W. Functional properties of microorganisms in fermented foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef]
- Hutkins, R.W. Microbiology and Technology of Fermented Foods; Blackwell Publishing: Oxford, UK, 2007; pp. 1–473. [Google Scholar] [CrossRef]
- Adams, G.G.; Imran, S.; Wang, S.; Mohammad, A.; Kok, M.S.; Gray, D.A.; Channell, G.A.; Harding, S.E. The hypoglycemic effect of pumpkin seeds, trigonelline (TRG), nicotinic acid (NA), and D-chiro-inositol (DCI) in controlling glycemic levels in diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2014, 54, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Wang, X.; Zhang, M.; Ren, Y.; Yue, T.; Gao, Z. Effect of mixed lactobacillus on the physicochemical properties of cloudy apple juice with the addition of polyphenols-concentrated solution. Food Biosci. 2021, 41, 101049. [Google Scholar] [CrossRef]
- De Montijo-Prieto, S.; Razola-Díaz, M.d.C.; Barbieri, F.; Tabanelli, G.; Gardini, F.; Jiménez-Valera, M.; Ruiz-Bravo, A.; Verardo, V.; Gómez-Caravaca, A.M. Impact of lactic acid bacteria fermentation on phenolic compounds and antioxidant activity of avocado leaf extracts. Antioxidants 2023, 12, 298. [Google Scholar] [CrossRef]
- Yang, F.; Chen, C.; Ni, D.; Yang, Y.; Tian, J.; Li, Y.; Chen, S.; Ye, X.; Wang, L. Effects of fermentation on bioactivity and the composition of polyphenols contained in polyphenol-rich foods: A review. Foods 2023, 12, 3315. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Tang, F.; Cai, W.; Peng, B.; Zhang, P.; Shan, C. Effect of fermentation by lactic acid bacteria on the phenolic composition, antioxidant activity, and flavor substances of jujube–wolfberry composite juice. LWT—Food Sci. Technol. 2023, 184, 114884. [Google Scholar] [CrossRef]
- Zhao, Y.-S.; Eweys, A.S.; Zhang, J.-Y.; Zhu, Y.; Bai, J.; Darwesh, O.M.; Zhang, H.-B.; Xiao, X. Fermentation affects the antioxidant activity of plant-based food material through the release and production of bioactive components. Antioxidants 2021, 10, 2004. [Google Scholar] [CrossRef]
- Dissanayake, I.H.; Tabassum, W.; Alsherbiny, M.; Chang, D.; Li, C.G.; Bhuyan, D.J. Lactic acid bacterial fermentation as a biotransformation strategy to enhance the bioavailability of phenolic antioxidants in fruits and vegetables: A comprehensive review. Food Res. Int. 2025, 209, 116283. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Saranyadevi, S.; Thirumalaisamy, S.K.; Durage, T.T.D.; Jaiswal, S.G.; Kavitake, D.; Wei, S. Phenolic acids in fermented foods: Microbial biotransformation, antioxidant mechanisms, and functional health implications. Front. Mol. Biosci. 2025, 12, 1678673. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects–A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Farazi, M.; Houghton, M.J.; Nicolotti, L.; Murray, M.; Cardoso, B.R.; Williamson, G. Inhibition of human starch digesting enzymes and intestinal glucose transport by walnut polyphenols. Food Res. Int. 2024, 189, 114572. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hong, J.; Wang, L.; Cai, C.; Mo, H.; Wang, J.; Fang, X.; Liao, Z. Effect of lactic acid bacteria fermentation on plant-based products. Fermentation 2024, 10, 48. [Google Scholar] [CrossRef]
- Mechchate, H.; Es-Safi, I.; Louba, A.; Alqahtani, A.S.; Nasr, F.A.; Noman, O.M.; Farooq, M.; Alharbi, M.S.; Alqahtani, A.; Bari, A.; et al. In vitro alpha-amylase and alpha-glucosidase inhibitory activity and in vivo antidiabetic activity of withania frutescens L. Foliar extract. Molecules 2021, 26, 293. [Google Scholar] [CrossRef]
- Gomes-Lobo, C.; Pérez-Correa, J.; Franco, W.; Pérez-Jiménez, J. Modulation of phenolic compounds in murta (Ugni molinae Turcz) juice by lactic fermentation in monoculture and coculture: Identification and quantification of extractable and non-extractable fractions by HPLC-QTOF-MS. Food Chem. 2025, 497, 146954. [Google Scholar] [CrossRef]
- Poquet, L.; Clifford, M.N.; Williamson, G. Bioavailability of flavanols and phenolic acids. In Plant Phenolics and Human Health; Fraga, C.G., Ed.; Wiley-Blackwell: Oxford, UK, 2009; pp. 51–89. [Google Scholar] [CrossRef]
- Paramithiotis, S. The future of lactic acid fermentation of fruits and vegetables. In Lactic Acid Fermentation of Fruits and Vegetables; Paramithiotis, S., Ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 283–290. [Google Scholar] [CrossRef]
- Oh, B.-T.; Jeong, S.-Y.; Velmurugan, P.; Park, J.-H.; Jeong, D.-Y. Probiotic-mediated blueberry (Vaccinium corymbosum L.) fruit fermentation to yield functionalized products for augmented antibacterial and antioxidant activity. J. Biosci. Bioeng. 2017, 124, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Frediansyah, A.; Romadhoni, F.; Suryani; Nurhayati, R.; Wibowo, A.T. Fermentation of jamaican cherries juice using lactobacillus plantarum elevates antioxidant potential and inhibitory activity against Type II diabetes-related enzymes. Molecules 2021, 26, 2868. [Google Scholar] [CrossRef]
- Schmidt, J.; Hotz, H.G.; Foitzik, T.; Ryschich, E.; Buhr, H.J.; Warshaw, A.L.; Herfarth, C.; Klar, E. Intravenous contrast medium aggravates the impairment of pancreatic microcirculation in necrotizing pancreatitis in the rat. Ann. Surg. 1995, 221, 257–264. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Tybussek, T.; Herfellner, T.; Schneider, F.; Schweiggert-Weisz, U.; Eisner, P. Radical scavenging mechanisms of phenolic compounds: A quantitative structure-property relationship (QSPR) study. Front. Nutr. 2022, 9, 882458. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compounds | LP | LB | LPC | LGG | LC | LA | Control |
|---|---|---|---|---|---|---|---|
| Gallic acid | 1996.32 ± 7.27a | 1356.60 ± 8.03d | 1728.69 ± 21.85b | 1657.57 ± 7.66c | 1647.63 ± 1.99c | 1296.90 ± 1.85d | 1143.94 ± 4.60e |
| 3,4-Dihydroxybenzoic acid | 1855.24 ± 5.03a | 1325.58 ± 0.78e | 1751.44 ± 16.75b | 1684.29 ± 6.45c | 1648.10 ± 2.67c | 1396.69 ± 1.64d | 1244.75 ± 5.78f |
| 4-Hydroxybenzoic acid | 1780.47 ± 19.78a | 1391.65 ± 1.55d | 1707.10 ± 3.04a | 1610.55 ± 9.63b | 1594.41 ± 2.10b | 1494.47 ± 4.51c | 1306.85 ± 4.00e |
| Catechin | 175.59 ± 4.54a | 137.61 ± 2.20c | 169.01 ± 0.98a | 162.46 ± 2.48b | 155.08 ± 2.00b | 144.04 ± 1.57c | 129.54 ± 3.56d |
| Caffeic acid | 1894.16 ± 4.13a | 1357.39 ± 9.00e | 1745.71 ± 32.29bc | 1709.05 ± 6.06bc | 1695.54 ± 2.66c | 1455.52 ± 3.02d | 1324.66 ± 2.09e |
| Syringic acid | 1860.77 ± 5.05a | 1507.14 ± 4.38d | 1745.08 ± 10.41bc | 1723.50 ± 2.40c | 1713.91 ± 2.09c | 1545.52 ± 2.90d | 1445.60 ± 7.65d |
| Vanillin | 1792.07 ± 7.24a | 1456.51 ± 11.71d | 1734.33 ± 4.19a | 1636.81 ± 3.84b | 1626.15 ± 2.64b | 1548.55 ± 2.57c | 1361.59 ± 9.95e |
| p-Hydroxycinnamic acid | 1645.75 ± 5.02a | 1352.84 ± 3.7d | 1604.23 ± 2.95a | 1595.97 ± 3.41ab | 1541.00 ± 3.70bc | 1494.68 ± 2.50c | 1301.06 ± 1.67d |
| Salicylic acid | 176.21 ± 3.15a | 126.51 ± 3.14d | 162.55 ± 0.66b | 151.10 ± 2.03c | 146.09 ± 2.66c | 132.55 ± 2.37d | 118.40 ± 2.92e |
| Vitexin | 181.88 ± 2.91a | 133.06 ± 1.65ef | 158.22 ± 1.47b | 147.69 ± 1.97c | 142.99 ± 2.08cd | 139.73 ± 0.90de | 128.68 ± 2.48f |
| Trans-ferulic acid | 3894.29 ± 1.25a | 2999.29 ± 4.71d | 3732.22 ± 14.20a | 3581.24 ± 9.99a | 3215.39 ± 13.24c | 3425.91 ± 13.61b | 2506.20 ± 15.25e |
| Quercetin-3-β-D-glucoside | 182.66 ± 3.22a | 123.15 ± 2.62e | 162.89 ± 2.62b | 149.61 ± 2.49c | 148.87 ± 1.46c | 134.81 ± 4.40d | 111.65 ± 3.40f |
| Luteoloside | 174.99 ± 1.90a | 128.24 ± 2.47c | 167.32 ± 1.42a | 156.63 ± 1.52b | 148.69 ± 2.22b | 134.89 ± 3.12c | 119.70 ± 1.21d |
| (+)-Dihydroquercetin | 170.72 ± 2.58a | 136.34 ± 4.15c | 165.76 ± 2.67a | 164.74 ± 2.56a | 155.80 ± 2.07b | 152.29 ± 3.87b | 130.40 ± 3.45c |
| Benzoic acid | 1764.91 ± 3.98a | 1484.82 ± 17.04c | 1746.65 ± 35.11a | 1707.20 ± 4.43a | 1695.58 ± 2.51a | 1576.46 ± 11.00b | 1444.33 ± 5.40c |
| Kaempferol-3-O-glucoside | 168.52 ± 0.64a | 137.52 ± 1.53cd | 165.06 ± 2.28a | 159.10 ± 1.90ab | 154.16 ± 2.69b | 144.78 ± 4.84c | 129.01 ± 1.89d |
| Daidzein | 181.99 ± 2.81a | 147.53 ± 1.43c | 177.50 ± 1.84a | 163.72 ± 1.69b | 164.81 ± 1.50b | 156.89 ± 1.30b | 136.78 ± 2.72d |
| Luteolin | 172.33 ± 1.65a | 135.85 ± 2.73d | 165.81 ± 1.59ab | 157.50 ± 1.98bc | 154.34 ± 2.06bcd | 146.70 ± 2.57d | 128.45 ± 2.34e |
| Quercetin | 166.36 ± 2.41a | 148.06 ± 2.46bc | 164.28 ± 0.52a | 162.58 ± 2.53a | 163.70 ± 2.16a | 153.65 ± 2.21b | 144.54 ± 1.42c |
| Hydrocinnamic acid | 1855.03 ± 8.85a | 1705.19 ± 4.17c | 1832.75 ± 10.49ab | 1793.33 ± 7.11ab | 1772.85 ± 6.41ab | 1740.28 ± 4.99bc | 1693.27 ± 3.20c |
| Apigenin | 182.32 ± 2.58a | 153.91 ± 2.08c | 177.78 ± 1.51a | 177.65 ± 2.16a | 168.06 ± 1.85b | 155.81 ± 3.49c | 147.71 ± 2.79c |
| Naringenin | 190.41 ± 2.29a | 164.80 ± 2.39c | 186.99 ± 0.26a | 185.71 ± 1.89a | 181.75 ± 1.38ab | 175.15 ± 4.67b | 158.78 ± 2.63c |
| Kaempferol | 174.14 ± 1.61a | 165.63 ± 2.68ab | 165.63 ± 3.66ab | 159.37 ± 1.55bc | 135.70 ± 3.11d | 152.21 ± 2.42c | 143.30 ± 2.10d |
| Isorhamnetin | 176.52 ± 2.13a | 166.34 ± 2.11b | 165.47 ± 2.10b | 164.95 ± 2.52b | 134.98 ± 2.65d | 153.88 ± 1.90c | 142.40 ± 1.79d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Qayyum, A.; Junejo, S.A.; Xu, Z.; Hassan, M.Z.; Liu, B.; Chen, Z. The Effect of Lactic Acid Bacteria Fermentation on the Anti-Diabetic Activity of Pumpkin Puree. Foods 2026, 15, 1882. https://doi.org/10.3390/foods15111882
Qayyum A, Junejo SA, Xu Z, Hassan MZ, Liu B, Chen Z. The Effect of Lactic Acid Bacteria Fermentation on the Anti-Diabetic Activity of Pumpkin Puree. Foods. 2026; 15(11):1882. https://doi.org/10.3390/foods15111882
Chicago/Turabian StyleQayyum, Aqsa, Shahid Ahmed Junejo, Zuoting Xu, Muhammad Zubair Hassan, Bingjie Liu, and Zhong Chen. 2026. "The Effect of Lactic Acid Bacteria Fermentation on the Anti-Diabetic Activity of Pumpkin Puree" Foods 15, no. 11: 1882. https://doi.org/10.3390/foods15111882
APA StyleQayyum, A., Junejo, S. A., Xu, Z., Hassan, M. Z., Liu, B., & Chen, Z. (2026). The Effect of Lactic Acid Bacteria Fermentation on the Anti-Diabetic Activity of Pumpkin Puree. Foods, 15(11), 1882. https://doi.org/10.3390/foods15111882