
Nitraria sibirica Pall.: A Halophytic Resource for Antioxidant-Rich Functional Foods and Ecological Resilience


Abstract
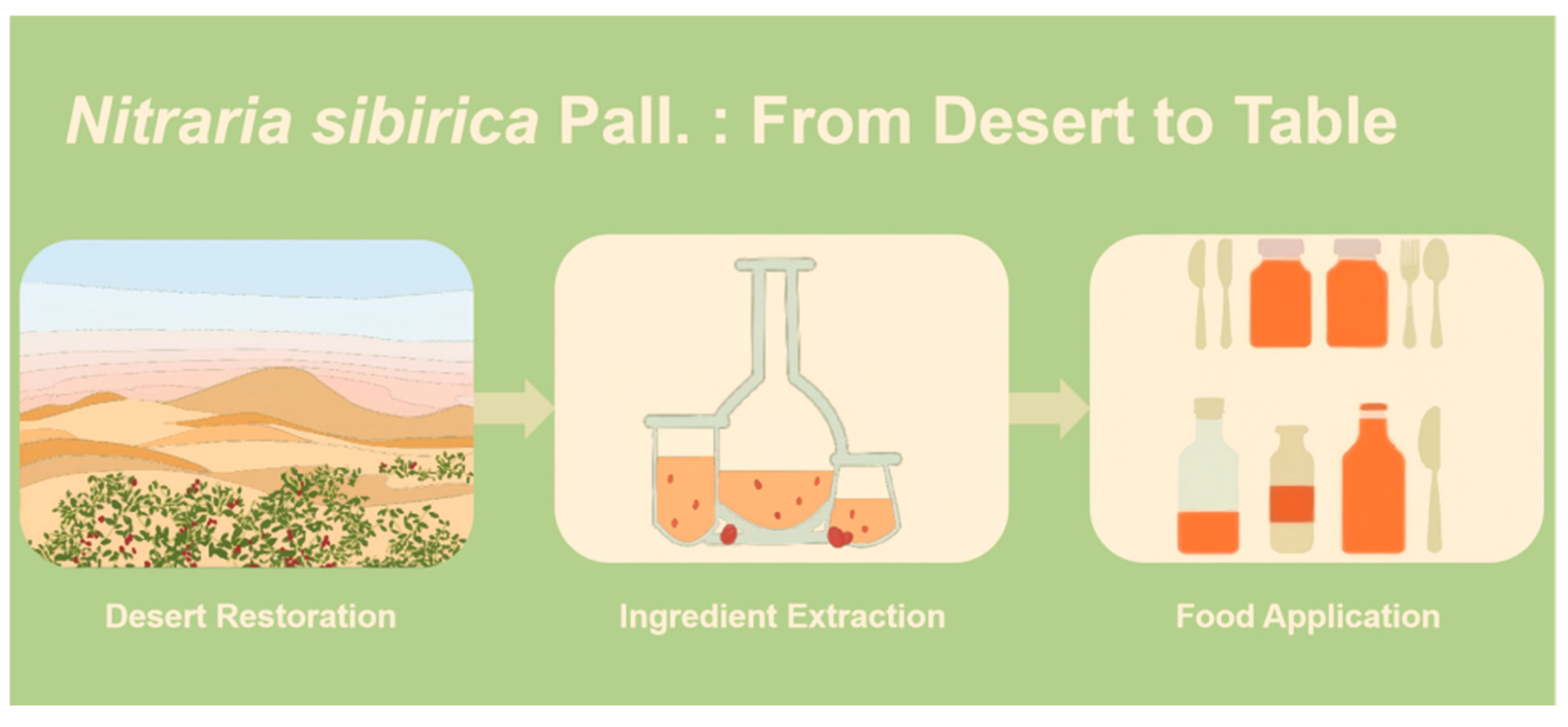
1. Introduction
2. Environmental Role of N. sibirica
2.1. Ion Homeostasis and Transport Regulation
2.2. Antioxidant Defense and Osmoprotectant Accumulation
2.3. Hormonal and Calcium Signaling
3. Chemical Composition Study
3.1. Phenolic Compounds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | Part of Plant | Molecular Formula | Reference |
|---|---|---|---|---|
| 1 | Rutin-7-O-α-L-rhamnopyranoside | Leaves | C33H40O20 | [51] |
| 2 | Clovin | Leaves | C33H40O20 | [51] |
| 3 | Rutin | Leaves; fruit | C27H30O16 | [51] |
| 4 | Narcissin | Leaves | C28H32O16 | [51] |
| 5 | Diosmin | Leaves | C28H32O15 | [51] |
| 6 | Quercetin-7-O-α-L-rhamnopyranoside | Leaves | C21H20O11 | [51] |
| 7 | Diosmetin | Leaves | C16H12O6 | [51] |
| 8 | Luteolin | Leaves | C15H10O6 | [51] |
| 9 | 3,5-dimethoxykaempferol-7-O-glucoside | Fruit | C23H24O11 | [52] |
| 10 | Chryseriol-3-O-rutinoside | Fruit | C28H32O15 | [52] |
3.2. Alkaloids
| No. | Compound | Part of Plant | Molecular Formula | Reference |
|---|---|---|---|---|
| 1 | Nitraramine | Aerial parts | C15H24N2O | [24,56] |
| 2 | Isonitramine | Aerial parts | C10H19NO | [53] |
| 3 | Sibirine | Aerial parts | C11H21NO | [25] |
| 4 | Nitrabirine | Aerial parts | C12H18N2O | [54,55] |
| 5 | Nitraramine N-oxide | Aerial parts | C15H24N2O2 | [55] |
| 6 | Deoxyvasicinone | Aerial parts | C11H10N2O | [55] |
| 7 | Schoberine | Aerial parts | C15H26N2 | [55] |
| 8 | Dehydroschoberine | Aerial parts | C15H24N2 | [55] |
| 9 | Dihydroschoberine | Aerial parts | C15H28N2 | [55] |
| 10 | Nitrabirine N-oxide | Aerial parts | C12H18N2O2 | [55] |
| 11 | Nitraramidine | Aerial parts | C23H30N2O3 | [56] |
| 12 | Nitraraidine | Aerial parts | C20H25N2 | [56] |
| 13 | Isonitramine | Aerial parts | C10H19NO | [56] |
| 14 | L-vasicinone | Aerial parts | C11H10N2O2 | [56] |
| 15 | Nitraroxine | Aerial parts | C15H24N2O2 | [56] |
| 16 | Sibirinine | Aerial parts | C12H21NO2 | [56] |
| 17 | Schobemine | Leaves | C15H24N2 | [58] |
| 18 | Schoberidine | Leaves | C20H21N3 | [58] |
| 19 | Schoberimine | Leaves | C22H25N3 | [58] |
| 20 | N-malonyl-tryptophan | Fruit | C14H14N2O5 | [59] |
| 21 | 5-(methoxymethyl)-1H-pyrrole-2-carbaldehyde | Fruit | C7H9NO2 | [59,60] |
| 22 | 2-[2-formyl-5-(methoxymethyl)-1H-pyrrol-1-yl]-propanoate | Fruit | C11H15NO4 | [59,61] |
3.3. Polysaccharides
3.4. Pigments
3.5. Lipids and Volatile Oils
3.6. Protein and Amino Acids
3.7. Other Constituents
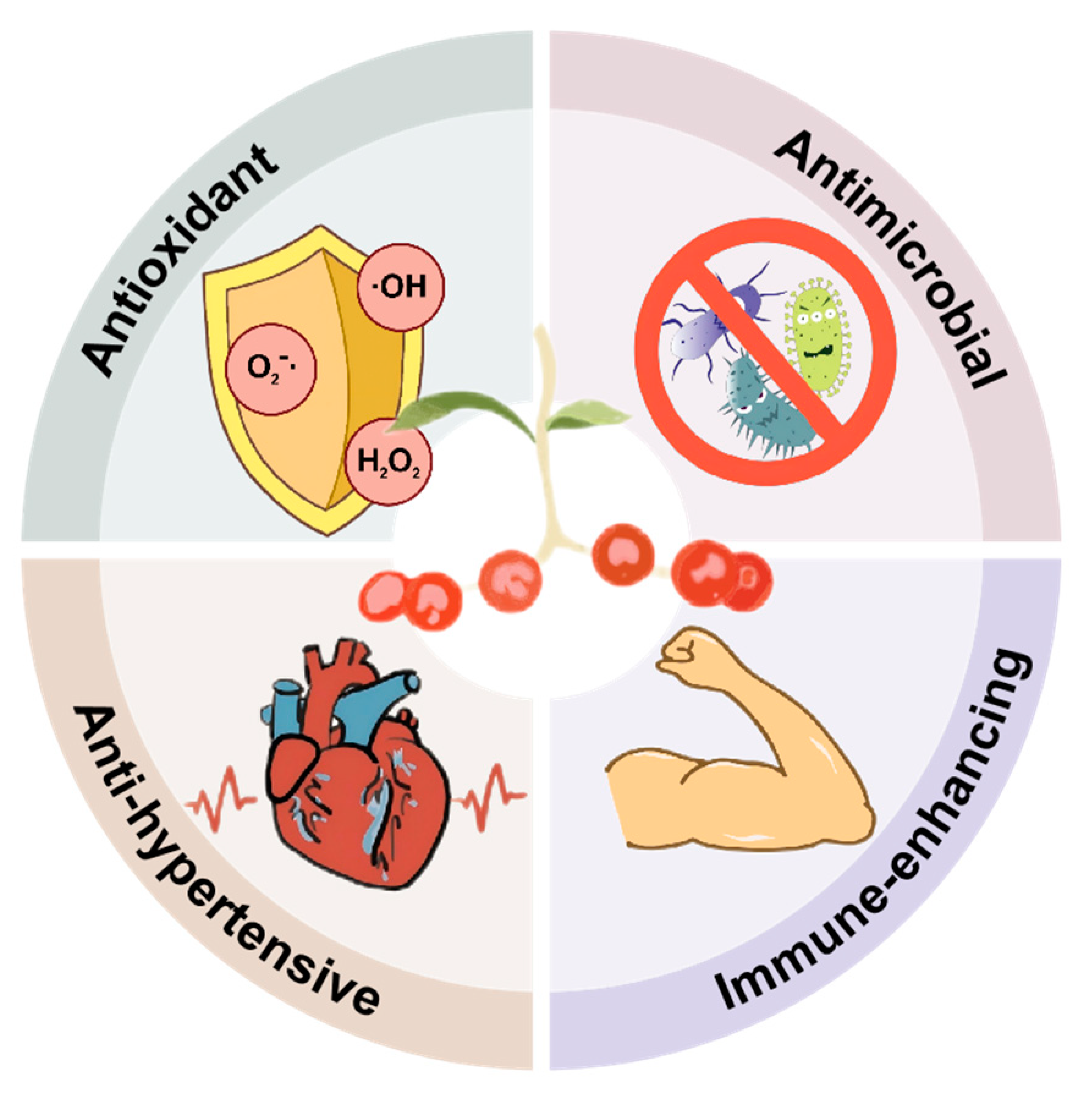
4. Pharmacological Effects
4.1. Antioxidant Activity
4.2. Antimicrobial Activity
4.3. Anti-Hypertensive Activity
4.4. Other Activities
| Pharmacological Activity | Key Findings/Data | Reference |
|---|---|---|
| Antioxidant | DPPH radical scavenging (IC50: 0.19 mg/mL; superior to L. barbarum and L. ruthenicum); Significant O2−· and DPPH scavenging by freeze-dried polysaccharides; Anthocyanins from subcritical water extraction stronger than ascorbic acid and 14x acarbose | [62,68,81,82] |
| Antimicrobial | Ethyl acetate extract: inhibition zones 12–18 mm (E. coli, B. subtilis, S. aureus); MIC: 25–50 mg/mL; No antifungal activity | [81,84] |
| Anti-hypertensive | Leaf extracts reduce BP in SHR models; Aqueous extract ACE inhibitory (IC50 = 55.85 g/L); Fruit extracts induce vasodilation via NOS; Total alkaloids reduce albuminuria in mice fed high-salt diet with Ang II | [23,88,89,90] |
| Immunomodulatory | Polysaccharides promote NO secretion in macrophages, immune enhancement; No pro-inflammatory effect unlike Lycium polysaccharides | [62] |
| Anti-obesity | Flavonoid-rich extract inhibits adipogenesis by 40% in 3T3-L1 cells | [83] |
| Hypoallergic effect | Pectin from N. sibirica inhibits ovalbumin absorption in mice (reduced serum OVA) | [26] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tomoshevich, M.; Banaev, E.; Khozyaykina, S.; Erst, A. Pollen morphology of some species from genus Nitraria. Plants 2022, 11, 2359. [Google Scholar] [CrossRef] [PubMed]
- Poliakova, T.A.; Banaev, E.V.; Tomoshevich, M.A. Identification of species in the genus Nitraria L. (Nitrariaceae) based on nucleotide variability of nuclear ribosomal DNA. Vavilov J. Genet. Breed. 2020, 24, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Banaev, E.V.; Tomoshevich, M.A.; Khozyaykina, S.A.; Erst, A.A.; Erst, A.S. Integrative taxonomy of Nitraria (Nitrariaceae), description of the new enigmatic species and key to all currently known species. Plants 2023, 12, 593. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, J.; Li, Y.; Yang, Z.; Wang, Q.; Jia, Q. Integrated metabolomic and transcriptomic analysis of Nitraria berries indicate the role of flavonoids in adaptation to high altitude. Metabolites 2024, 14, 591. [Google Scholar] [CrossRef]
- Wang, J.; Su, H.; Han, H.; Wang, W.; Li, M.; Zhou, Y.; Li, Y.; Li, M. Transcriptomics reveals host-dependent differences of polysaccharides biosynthesis in Cynomorium songaricum. Molecules 2021, 27, 44. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Wang, B.; Li, C.; Li, X.; He, C.; Li, Y. Integrated pacBio SMRT and illumina sequencing uncovers transcriptional and physiological responses to drought stress in whole-plant Nitraria tangutorum. Front. Genet. 2024, 15, 1474259. [Google Scholar] [CrossRef]
- Du, Q.; Xin, H.; Peng, C. Pharmacology and phytochemistry of the Nitraria genus. Mol. Med. Rep. 2015, 11, 11–20. [Google Scholar] [CrossRef]
- Song, L.; Liu, S.; Zhang, L.; Pan, L.; Xu, L. Polysaccharides from Nitraria retusa fruit: Extraction, purification, structural characterization, and antioxidant activities. Molecules 2023, 28, 1266. [Google Scholar] [CrossRef]
- Ren, S.; Jiang, L.; Lv, G. A Study on endogenous inhibitors of Nitraria roborowskii Kom. Seeds. Forests 2024, 15, 773. [Google Scholar] [CrossRef]
- Myagmarsuren, D.; Danzhalova, E.V.; Bayasgalan, D. Vegetation of the sand massifs of the northern Gobi Desert. Arid Ecosyst. 2024, 14, 307–315. [Google Scholar] [CrossRef]
- Kono, A.; Okuro, T. Development and validation of vegetation indicators for the occurrence of saltation that consider the spatial distribution of vegetation. Aeolian Res. 2023, 63–65, 100886. [Google Scholar] [CrossRef]
- Nyamjav, J.; Batsaikhan, M.E.; Li, G.; Li, J.; Luvsanjamba, A.; Jin, K.; Xiao, W.; Wu, L.; Indree, T.; Qin, A. Allometric equations for estimating above-ground biomass of Nitraria sibirica Pall. in Gobi Desert of Mongolia. PLoS ONE 2020, 15, e0239268. [Google Scholar] [CrossRef] [PubMed]
- Banaev, E.V.; Tomoshevich, M.A.; Yamtyrov, M.B. Variability of metric and qualitative traits of Nitraria species in relation to ecological and climatic conditions of Siberian habitats. Contemp. Probl. Ecol. 2017, 10, 664–673. [Google Scholar] [CrossRef]
- Banaev, E.V.; Khozyaykina, S.A.; Dorogina, O.V.; Tomoshevich, M.A. Genetic diversity of Nitraria sibirica Pall. (Nitrariaceae) in the populations of Siberia and Kazakhstan based on the polymorphism of intermicrosatellite Markers. Contemp. Probl. Ecol. 2022, 15, 400–408. [Google Scholar] [CrossRef]
- Ankipovich, E.; Lagunova, E. A study of the Overburden dumps flora at Chernogorsky quarry. In Proceedings of the Conference on Results and Prospects of Geobotanical Research in Siberia Dedicated to the 75th Anniversary of the Laboratory of Ecology and Geobotany of CSBG SB RAS, Novosibirsk, Russia, 13–17 May 2019. [Google Scholar]
- Xu, L.; Zhang, Y.; Read, N.; Liu, S.; Friman, V.-P. Devosia Nitraria sp nov., a novel species isolated from the roots of Nitraria sibirica in China. Antonie Van Leeuwenhoek 2017, 110, 1475–1483. [Google Scholar] [CrossRef]
- Hu, A.; Yang, X.; Zhu, J.; Wang, X.; Liu, J.; Wang, J.; Wu, H.; Zhang, H.; Zhang, H. Selection and validation of appropriate reference genes for RT-qPCR analysis of Nitraria sibirica under various abiotic stresses. BMC Plant Biol. 2022, 22, 592. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, W.; Zheng, X.; Li, S. Root distribution of Nitraria sibirica with seasonally varying water sources in a desert habitat. J. Plant Res. 2015, 128, 613–622. [Google Scholar] [CrossRef]
- Banaev, E.V.; Tomoshevich, M.A.; Ak-Lama, T.A. Morphological variability of Nitraria species in central and southern Kazakhstan. Contemp. Probl. of Ecol. 2021, 14, 255–268. [Google Scholar] [CrossRef]
- Wang, W.; Du, W.; Wang, S. Study on development of Nitraria sibirica Pall. in the East of Hebei Province. Chin. Wild Plant Resour. 2009, 28, 29–31, 35. [Google Scholar]
- Chang, Y.; Lv, G. Dynamic change mechanism of the desert plant Nitraria sibirica growth in natural habitat. Ecol. Indic. 2023, 154, 110695. [Google Scholar] [CrossRef]
- Cheng, T.; Li, H.; Wu, H.; Liu, Z.; Wu, x.; Yang, S.; Zhang, H.; Yang, X. Comparison on osmotica accumulation of different salt-tolerant plants under salt stress. Hubei Agric. Sci. 2016, 55, 3674–3677. [Google Scholar]
- Turghun, C.; Bakri, M.; Abdulla, R.; Ma, Q.; Aisa, H.A. Comprehensive characterisation of phenolics from Nitraria sibirica leaf extracts by UHPLC-quadrupole-orbitrap- MS and evaluation of their anti-hypertensive activity. J. Ethnopharmacol. 2020, 261, 113019. [Google Scholar] [CrossRef] [PubMed]
- Novgorodova, N.Y.; Maekh, S.K.; Yunusov, S.Y. Nitraramine—A new alkaloid from Nitraria schoberi. Chem. Nat. Compd. 1975, 11, 455–456. [Google Scholar] [CrossRef]
- Osmanov, Z.; Ibragimov, A.A.; Yunusov, S.Y. Structure of sibirine. Chem. Nat. Compd. 1982, 18, 206–208. [Google Scholar] [CrossRef]
- Golovchenko, V.V.; Khramova, D.S.; Shashkov, A.S.; Otgonbayar, D.; Chimidsogzol, A.; Ovodov, Y.S. Structural characterisation of the polysaccharides from endemic Mongolian desert plants and their effect on the intestinal absorption of ovalbumin. Carbohydr. Res. 2012, 356, 265–272. [Google Scholar] [CrossRef]
- Xue, Y.; Wang, T.; Xue, Y.; Liu, X.; Zhang, D.; Li, Y.; Gao, H. Comparison study of main active compounds in Nitraria from inner Mongolia. Sci. Technol. Food Ind. 2014, 35, 106–108+117. [Google Scholar]
- Jinmei, W.; Wenyi, K. Volatiles from the Leaves of Nitraria tangutorum and Nitraria sibirica. Nat. Prod. Res. Dev. 2011, 23, 680–683. [Google Scholar]
- Li, C.Y.; Ma, X.; Guo, P.; Qu, J.S. Comprehensive utilization research of desert “Cherry”—Nitraria Fruit. Chem. Ind. For. Prod. 1991, 11, 225–230. (In Chinese) [Google Scholar]
- Lu, L.; Wang, Y.; Chen, Y.; Zhu, L.; Wu, X.; Shi, J.; Chen, J.; Cheng, T. Salt stimulates carbon fixation in the halophyte Nitraria sibirica to enhance growth. For. Res. 2025, 5, e004. [Google Scholar] [CrossRef]
- Ni, J.; Wu, X.; Zhang, H.; Liu, T.; Zhang, L. Comparative analysis of salt tolerance of three Nitraria Species. Forest Res. 2012, 25, 48–53. [Google Scholar]
- Hou, Y.; Luo, J.; Li, X.; Cui, X.; Jia, B.; Diao, F.; Wang, L.; Guo, W. Effects of arbuscular mycorrhizal fungi on the growth of Nitraria sibirica under salinity and heavy metal stress. J. South China Agric. Univ. 2022, 43, 68–78. [Google Scholar]
- Li, H.; Tang, X.; Yang, X.; Zhang, H. Comprehensive transcriptome and metabolome profiling reveal metabolic mechanisms of Nitraria sibirica Pall. to salt stress. Sci. Rep. 2021, 11, 12878. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, H.; Shabala, S.; Li, H.; Yang, X.; Zhang, H. Tissue tolerance mechanisms conferring salinity tolerance in a halophytic perennial species Nitraria sibirica Pall. Tree Physiol. 2021, 41, 1264–1277. [Google Scholar] [CrossRef]
- Yan, H.; Zhang, H.; Feng, F.; Yu, Z.; Yang, X. Metabolic response mechanism of two Nitraria species to salt stress. Sci. Silvae Sin. 2021, 57, 20–29. [Google Scholar]
- Zhang, P.; Zhang, F.; Wu, Z.; Cahaeraduqin, S.; Liu, W.; Yan, Y. Analysis on the salt tolerance of Nitraria sibirica Pall. based on Pacbio full-length transcriptome sequencing. Plant Cell Rep. 2023, 42, 1665–1686. [Google Scholar] [CrossRef]
- Mei, X.; Dai, T.; Shen, Y. Adaptive strategy of Nitraria sibirica to transient salt, alkali and osmotic stresses via the alteration of Na+/K+ fluxes around root tips. J. For. Res. 2022, 34, 425–432. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, J.; Yang, X.; Zhang, H.; Zhang, H. Genome identification and expression analysis of NHX gene family in woody halophyte Nitraria sibirica. Genom. Appl. Biol. 2023, 42, 698–714. [Google Scholar]
- Zhang, H.; Qi, C.; Li, C.; Huang, D.; Mao, H.; Lin, X. Overexpression of high affinity K+ transporter from Nitraria sibirica enhanced salt tolerance of transgenic plants. Plant Sci. 2024, 342, 112052. [Google Scholar] [CrossRef]
- Chen, S.; Geng, X.; Lou, J.; Huang, D.; Mao, H.; Lin, X. Overexpression of a plasmalemma Na+/H+ antiporter from the halophyte Nitraria sibirica enhances the salt tolerance of transgenic poplar. Plant Sci. 2024, 343, 112061. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, W.; Wang, H.; Wu, H.; Xiao, Y.; Yan, Y. Effects of exogenous CaCl2 on 2reactive oxygen species metabolism in Nitraria sibirica under NaCl stress. Plant Physiol. J. 2021, 57, 1105–1112. [Google Scholar]
- Chang, Y.; Lv, G. Nitraria sibirica adapts to long-term soil water deficit by reducing photosynthesis, stimulating antioxidant systems, and accumulating osmoregulators. Plant Physiol. Biochem. 2024, 206, 108265. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Lv, G.; He, X. Growth adaptation of Nitraria sibirica in drought: Hormone-mediated flavonoid accumulation, altered metabolic pathways, and decreased network complexity. Environ. Exp. Bot. 2024, 221, 105720. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, J.; Zhu, L.; Tang, Y.; Hao, Z.; Zhang, J.; Shi, J.; Cheng, T.; Lu, L. Genome-wide analyses of calmodulin and calmodulin-like proteins in the halophyte Nitraria sibirica reveal their involvement in response to salinity, drought and cold stress. Int. J. Biol. Macromol. 2023, 253, 127442. [Google Scholar] [CrossRef]
- Yan, Q.; Wang, S.; Zhang, Y.; Huang, A.; Na, D. Cloning and expression analysis of NsCBL2 gene from Nitraria sibirica. J. Anhui Agric. Univ. 2022, 49, 540–546. [Google Scholar]
- Banaev, E.V.; Voronkova, M.S.; Vysochina, G.I.; Tomoshevich, M.A. Population structure and differentiation of the Siberian representatives of the genus Nitraria L. (Nitrariaceae) based on the composition and content of phenolic compounds in leaves. Contemp. Probl. Ecol. 2015, 8, 735–742. [Google Scholar] [CrossRef]
- Aydin, D.; Coskun, O.F. Comparison of edta-enhanced phytoextraction strategies with Nasturtium officinale (Watercress) on an artificially arsenic contaminated water. Pak. J. Bot. 2013, 45, 1423–1429. [Google Scholar]
- Coskun, F.O.; Gulsen, O. Molecular, morphological and phytochemical characterization of some watermelon (Citrullus lanatus L.) genotypes. Hortic. Sci. 2024, 51, 189–201. [Google Scholar] [CrossRef]
- Turghun, C.; Bakri, M.; Zou, G.A.; Bobakulov, K.M.; Aisa, H.A. Phenolic compounds from leaves of Nitraria sibirica. Chem. Nat. Compd. 2018, 54, 987–989. [Google Scholar] [CrossRef]
- Turghun, C.; Bakri, M.; Liu, G.Y.; Bobakulov, K.; Aisa, H.A. Phenolic glycosides from Nitraria sibirica leaves and their in vitro biological activities. Nat. Prod. Res. 2021, 35, 1388–1392. [Google Scholar] [CrossRef]
- Turghun, C.; Bobakulov, K.M.; Bakri, M.; Aisa, H.A. Flavonoids from Leaves of Nitraria sibirica. Chem. Nat. Compd. 2019, 55, 1156–1158. [Google Scholar] [CrossRef]
- Song, Q.; Xia, X.; Ji, C.; Chen, D.; Lu, Y. Optimized flash extraction and UPLC-MS analysis on antioxidant compositions of Nitraria sibirica fruit. J. Pharm. Biomed. Anal. 2019, 172, 379–387. [Google Scholar] [CrossRef]
- Osmanov, Z.; Ibragimov, A.A.; Yunusov, S.Y. Isonitramine—A new alkaloid from Nitraria sibirica. Chem. Nat. Compd. 1977, 13, 607–608. [Google Scholar] [CrossRef]
- Ibragimov, A.A.; Yunusov, S.Y. Alkaloids of Nitraria schoberi. Structure of nitraraine. Chem. Nat. Compd. 1985, 21, 502–509. [Google Scholar] [CrossRef]
- Tulyaganov, T.S.; Allaberdiev, F.K. Alkaloids of Nitraria sibirica: Dihydroschoberine and nitrabirine N-oxide. Chem. Nat. Compd. 2001, 37, 556–558. [Google Scholar] [CrossRef]
- Tulyaganov, T.S.; Allaberdiev, F.K. Alkaloids of Nitraria sibirica: Structures of nitraramidine and nitraraidine. Chem. Nat. Compd. 2002, 38, 602–604. [Google Scholar] [CrossRef]
- Banaev, E.V.; Vysochina, G.I.; Kukushkina, T.A. Variability in the content of biologically active substances in the leaves of Nitraria sibirica Pall. (Nitrariaceae). Contemp. Probl. Ecol. 2014, 7, 90–96. [Google Scholar] [CrossRef]
- Bakri, M.; Chen, Q.; Ma, Q.; Yang, Y.; Abdukadir, A.; Aisa, H.A. Separation and purification of two new and two known alkaloids from leaves of Nitraria sibirica by pH-zone-refining counter-current chromatography. J. Chromatogr. B 2015, 1006, 138–145. [Google Scholar] [CrossRef]
- Song, Q.B.; Zhao, H.D.; Fu, Z.L.; Chen, D.F.; Lu, Y. Antioxidant and anticomplement compounds isolated from Nitraria sibirica fruit by High-Speed Counter-Current Chromatography. Pharmacogn. Mag. 2018, 14, 541–547. [Google Scholar]
- Don, M.-J.; Shen, C.-C.; Lin, Y.-L.; Syu, W., Jr.; Ding, Y.-H.; Sun, C.-M. Nitrogen-containing compounds from Salvia miltiorrhiza. J. Nat. Prod. 2005, 68, 1066–1070. [Google Scholar] [CrossRef]
- Joung Youn, U.; Kil, Y.S.; Nam, J.W.; Jin Lee, Y.; Kim, J.; Lee, D.; Lee, J.H.; Seo, E.K. New Pyrrole Alkaloids with Bulky N-Alkyl Side chains containing stereogenic centers from Lycium chinense. Helv. Chim. Acta 2013, 96, 1482–1487. [Google Scholar] [CrossRef]
- Zhang, Q.; Gao, H.; Ling, L.; Ji, C.; Lu, Y.; Chen, D. Crude polysaccharides extracted from three characteristic plants in Qinghai province and their activities. Nat. Prod. Res. Dev. 2017, 29, 34–39. [Google Scholar]
- Abuduwaili, A.; Mutailifu, P.; Nuerxiati, R.; Gao, Y.; Aisa, H.A.; Yili, A. Structure and biological activity of polysaccharides from Nitraria sibirica pall fruit. Food Biosci. 2021, 40, 100903. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Wang, M.; Zheng, Y.; Wang, M. Pigment extraction from fruits of Nitraria sibirica fruit and its physical and chemical propertries. J. Shandong Agric. Univ. 2005, 36, 75–81. [Google Scholar]
- Zhu, Y.; Liu, J.; Cheng, Y.; Du, H. Study on stability of red pigment extracted from fruit of Nitraria sibirica Pall. Food Sci. 2008, 29, 113–115. [Google Scholar]
- Yunus, Z. Study on red pigment extraction technology from fruit of N. sibirica Pall. Food Sci. 2008, 29, 181–185. [Google Scholar]
- Gao, Z.; Te, B.; Wang, J.; Wang, Y. Analysis of anthocyanins in Nitraria sibirica fruits at different ripening stage. Acta Agric. Boreali-Sin. 2014, 29, 130–134. [Google Scholar]
- Ren, L.; Dong, Q.; Liu, Z.; Wang, Y.; Tan, N.; Wang, H.; Hu, N. Optimization of subcritical water extraction, UPLC-triple-TOF-MS/MS analysis, antioxidant and α-glucosidase inhibitory activities of anthocyanins from Nitraria sibirica Pall. fruits. Food Chem. X 2024, 23, 101626. [Google Scholar] [CrossRef]
- Bao, X.; Zong, Y.; Hu, N.; Li, S.; Liu, B.; Wang, H. Functional R2R3-MYB transcription factor NsMYB1, regulating anthocyanin biosynthesis, was relative to the fruit color differentiation in Nitraria sibirica Pall. BMC Plant Biol. 2022, 22, 186. [Google Scholar] [CrossRef]
- Bao, X.; Zong, Y.; Hu, N.; Liu, B.; Wang, H. NsMYB5 of Nitraria sibirica Pall regulation for anthocyanin biosynthesis in fruits. Acta Agric. Boreali-Occident. Sin. 2023, 32, 1215–1222. [Google Scholar]
- Zhu, Y.; Liu, J.; Wang Hang, y.; Zhao, W.; Xiang, Y. Comparison and analysis on fruit oils of Nitraria prepared by supercrtical CO2 fluid extraction and impregnation method. Food Ferment. Ind. 2006, 32, 84–86. [Google Scholar]
- Hu, N.; Zhang, S.; Ji, Z.; You, J.; Suo, Y. Determination of fatty acids in three Nitraria species by precolumn fluorescence labeling for high-performance liquid chromatography and atmospheric pressure chemical ionization-mass spectrometr. Anal. Lett. 2014, 47, 2475–2487. [Google Scholar] [CrossRef]
- Gu, D.; Yang, Y.; Bakri, M.; Chen, Q.; Aisa, H.A. Biological activity and LC-MS profiling of ethyl acetate extracts from Nitraria sibirica (Pall.) fruits. Nat. Prod. Res. 2018, 32, 2054–2057. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Suo, Y. Amino acid content and its nutritional evaluation of Nitraria sibirica Pall. and Nitraria tangutorum Bobr. in Tsaidam Basin. Amino Acids Biotic Res. 2002, 24, 4–7. [Google Scholar]
- Zhang, Y.; Li, P.; Li, C.X.; Pan, H.P.; Zhao, Y.P.; Chen, S.L. Determination and analysis of Nutrient contents in leaves of three Nitraria species in Gansu. Pratacult. Sci. 2007, 24, 37–39. (In Chinese) [Google Scholar]
- Liu, L.P. Nutritional Composition Analysis and Evaluation of Four Nitraria Species in Inner Mongolia. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2009. (In Chinese). [Google Scholar]
- Ni, J.; Wu, X.; Zhang, H. Amino acids and mineral elements in leaves and branches of Nitraria sibirica in Tianjin. J. North-East For. Univ. 2012, 40, 41–43, 97. [Google Scholar]
- Liu, L.P.; Siqinbilige; Xu, Z.M.; He, Z. Nutritional composition and utilization analysis of Nitraria fruits in Alxa Desert region. Inner Mong. For. Sci. Technol. 2016, 42, 29–31+42. (In Chinese) [Google Scholar]
- Zhou, L.B. Factor analysis and cluster analysis of trace elements in Nitraria leaves from Qinghai region. J. Anhui Agric. Sci. 2010, 38, 10360–10361. (In Chinese) [Google Scholar]
- Zhang, N.; Li, Z.X.; Li, J.; Liu, J.; Dai, J.M.; Li, Y.W.; Li, S.X. Research progress on chemical constituents and biological activities of Poria cocos. World Sci. Technol. Mod. Tradit. Chin. Med. 2019, 21, 220–233. (In Chinese) [Google Scholar]
- Chen, G.L.; Qi, J.H.; Zhan, G.X.; Jun, R.H.; Ren, L.Y. Antioxidant and antimicrobial activities of extracts from fruits of Nitraria sibirica Pall. in vitro. In Proceedings of the 29th International Horticultural Congress on Horticulture—Sustaining Lives, Livelihoods and Landscapes (IHC)/6th International Symposium on Human Health Effects of Fruits and Vegetables (FAVHEALTH), Brisbane, Australia, 17–22 August 2014; pp. 137–141. [Google Scholar]
- Xie, R.; Wei, Y.; Ding, Y.; Liu, Y.; Li, M.; Zhang, J. Effects of different processing methods on the antioxidant activity of polysaccharide from Nitraria sibirica Pall. Nat. Prod. Res. Dev. 2016, 28, 41–45, 130. [Google Scholar]
- Zhao, T.; Ding, Y.; Sun, W.; Turghun, C.; Han, B. Ultrasonic-assisted extraction of flavonoids from Nitraria sibirica leaf using response surface methodology and their anti-proliferative activity on 3T3-L1 preadipocytes and antioxidant activities. J. Food Sci. 2023, 88, 2325–2338. [Google Scholar] [CrossRef]
- Zhang, G.; Qi, J.; Ren, X.; Chen, G. In vitro antimicrobial activity of extracts from fruits of Nitraria sibirica pall. Sci. Technol. Food Ind. 2012, 33, 104–106. [Google Scholar]
- Chen, X.; Lan, W.; Xie, J. Natural phenolic compounds: Antimicrobial properties, antimicrobial mechanisms, and potential utilization in the preservation of aquatic products. Food Chem. 2024, 440, 138198. [Google Scholar] [PubMed]
- Zhang, Z.; Zhao, L.; Zhou, X.; Meng, X.; Zhou, X. Role of inflammation, immunity, and oxidative stress in hypertension: New insights and potential therapeutic targets. Front. Immunol. 2022, 13, 1098725. [Google Scholar] [CrossRef]
- Liu, Y.M.; Liu, W.X. Pharmacography of Uighur; Science/Technology and Health Publishing House of Xinjiang: Urumqi, China, 1999; Volume II. [Google Scholar]
- Liu, Y.; Tan, C.; Wu, D.; Yi, L.; Zhao, Y.; Huang, X.; Wang, G.; Hu, J.; Liu, C. Study on inhibition activity of angiotensin I-converting enzyme (ACE) on 10 Halophytes. Chin. Pharm. J. 2010, 45, 736–738. [Google Scholar]
- Senejoux, F.; Girard, C.; Aisa, H.A.; Bakri, M.; Kerram, P.; Berthelot, A.; Bévalot, F.; Demougeot, C. Vasorelaxant and hypotensive effects of a hydroalcoholic extract from the fruits of Nitraria sibirica Pall. (Nitrariaceae). J. Ethnopharmacol. 2012, 141, 629–634. [Google Scholar] [CrossRef]
- Bakri, M.; Yi, Y.; Chen, L.-D.; Aisa, H.A.; Wang, M.-H. Alkaloids of Nitraria sibirica Pall, decrease hypertension and albuminuria in angiotensin II-salt hypertension. Chin. J. Nat. Med. 2014, 12, 266–272. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, K.; Zhang, X.; Zhao, L.; Xu, J.; Li, J. Nitraria sibirica Pall.: A Halophytic Resource for Antioxidant-Rich Functional Foods and Ecological Resilience. Foods 2025, 14, 1646. https://doi.org/10.3390/foods14091646
Lu K, Zhang X, Zhao L, Xu J, Li J. Nitraria sibirica Pall.: A Halophytic Resource for Antioxidant-Rich Functional Foods and Ecological Resilience. Foods. 2025; 14(9):1646. https://doi.org/10.3390/foods14091646
Chicago/Turabian StyleLu, Keyi, Xinmei Zhang, Liping Zhao, Jikun Xu, and Jianmei Li. 2025. "Nitraria sibirica Pall.: A Halophytic Resource for Antioxidant-Rich Functional Foods and Ecological Resilience" Foods 14, no. 9: 1646. https://doi.org/10.3390/foods14091646
APA StyleLu, K., Zhang, X., Zhao, L., Xu, J., & Li, J. (2025). Nitraria sibirica Pall.: A Halophytic Resource for Antioxidant-Rich Functional Foods and Ecological Resilience. Foods, 14(9), 1646. https://doi.org/10.3390/foods14091646