
Proteomics-Based Investigation of Sexual Dimorphism in Swim Bladder Texture of Chu’s Croaker (Nibea coibor)


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Tissue Collection
2.2. Textural Properties Analyses of Swim Bladder
2.3. Section and Masson Staining
2.4. Transmission Electron Microscopy (TEM)
2.5. Proximate Composition and Collagen Content Analysis
2.6. Collagen Content Analysis
2.7. Proteomics Analysis
2.8. Statistics
3. Results and Discussion
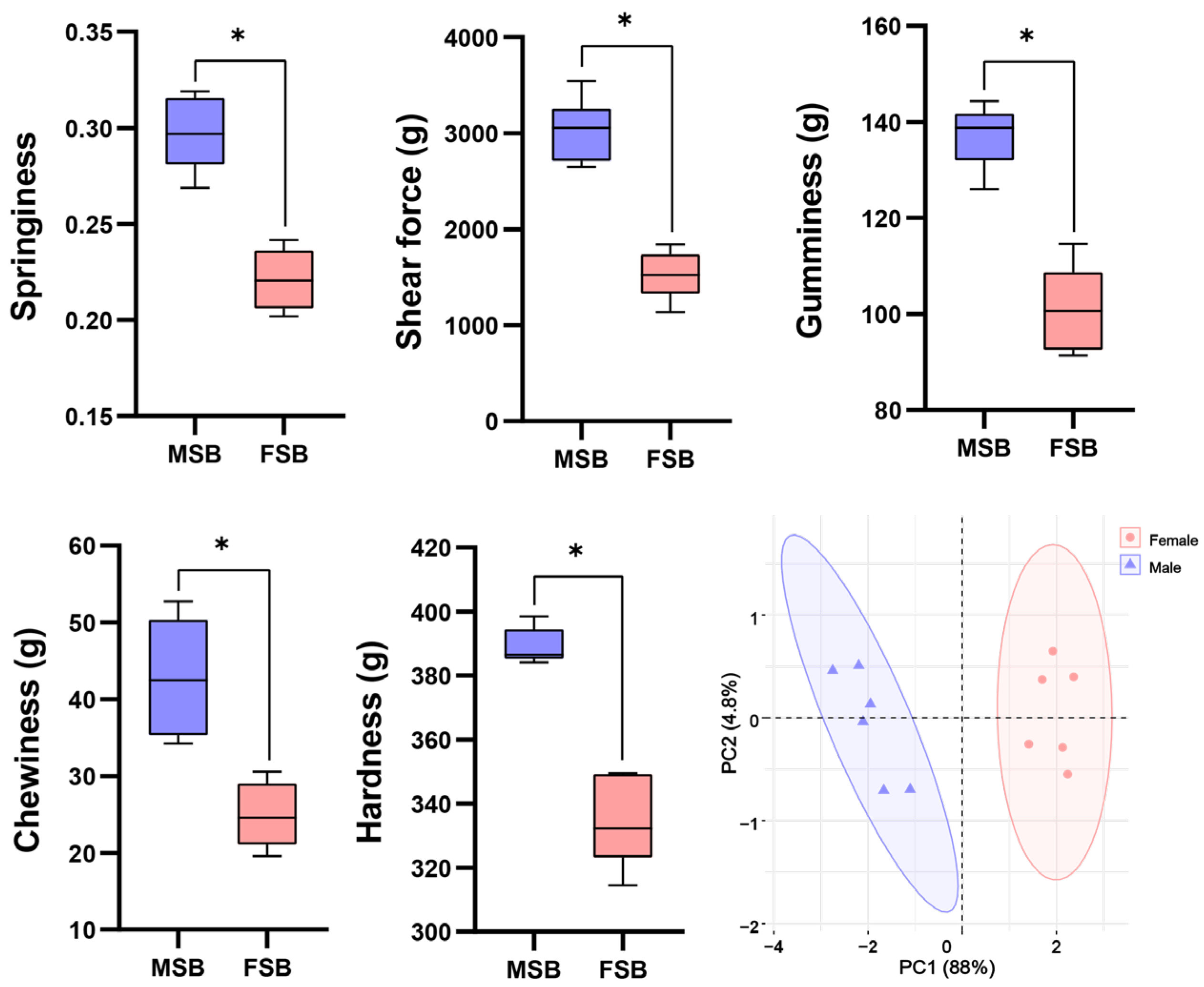
3.1. Texture Quality Between MSB and FSB
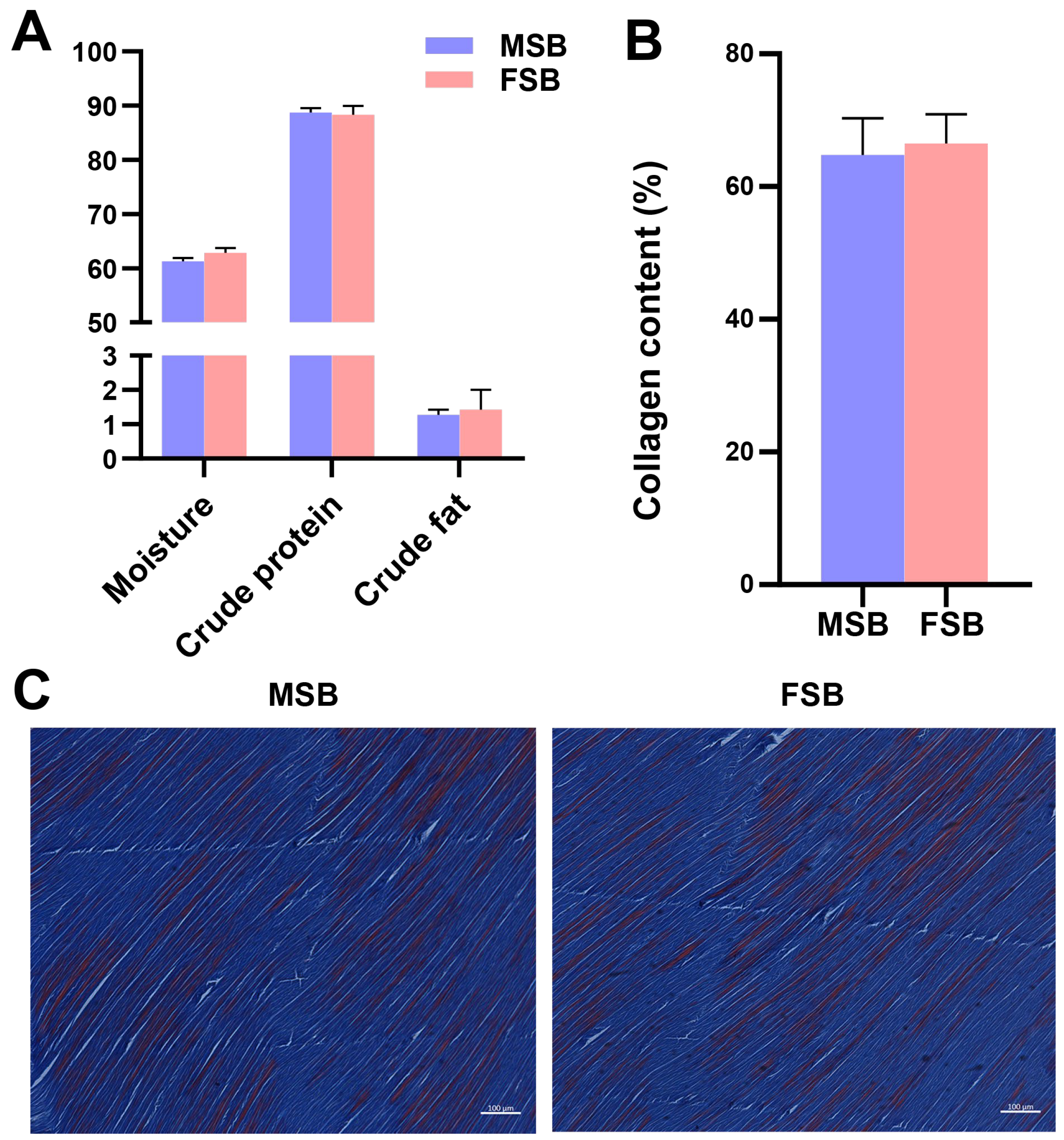
3.2. Proximate Composition and Collagen Content Between MSB and FSB
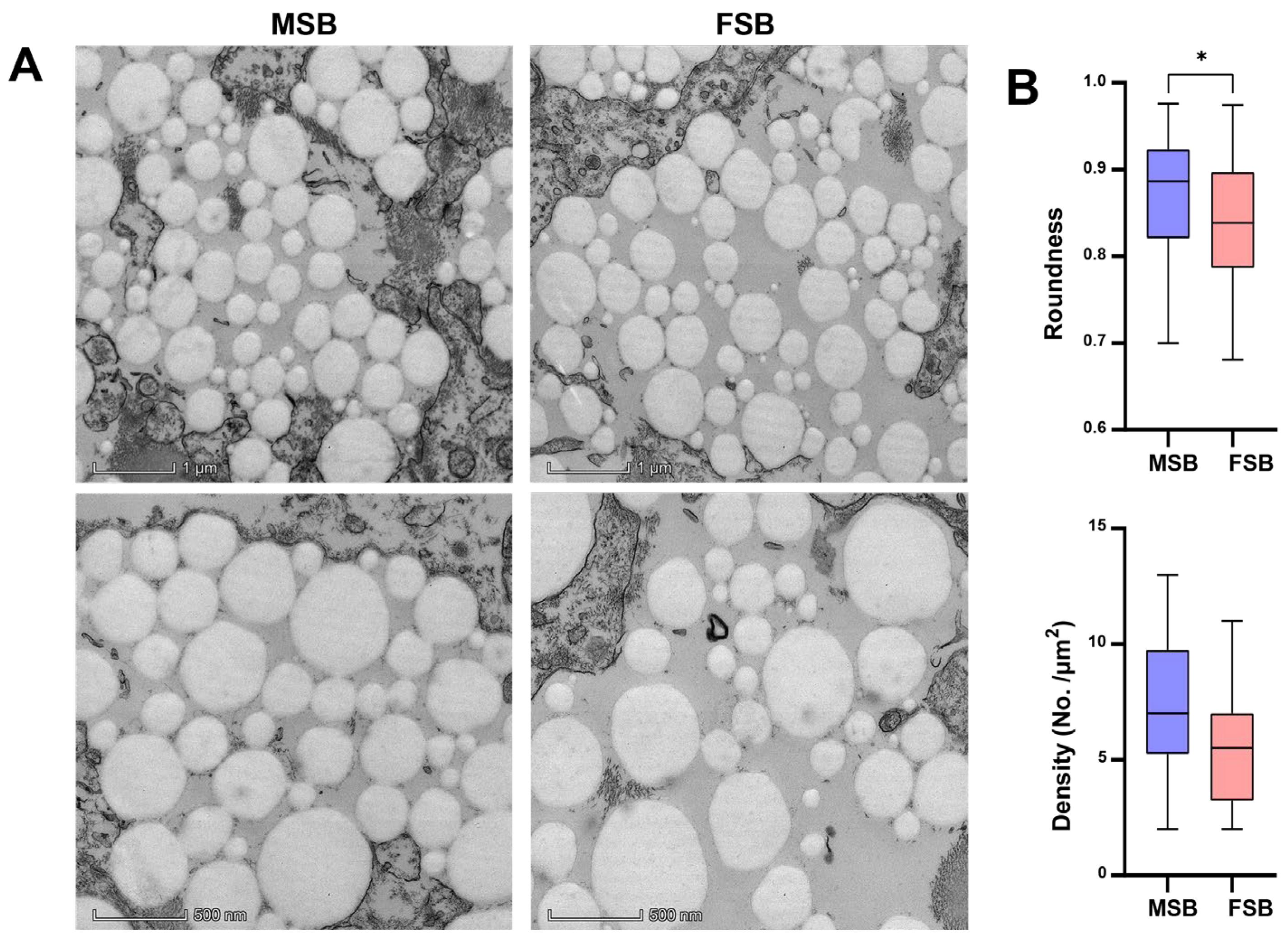
3.3. Microstructures Between MSB and FSB
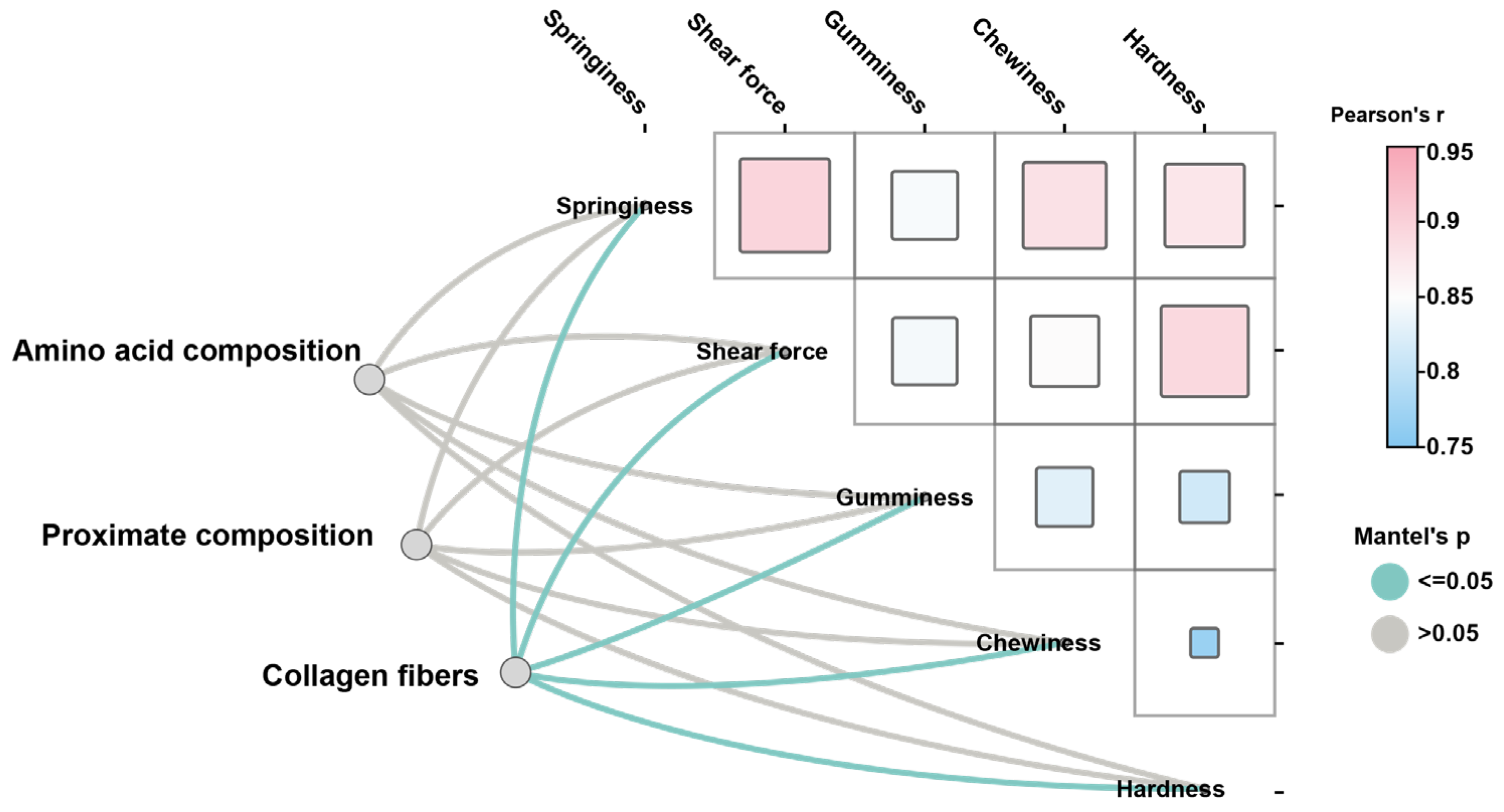
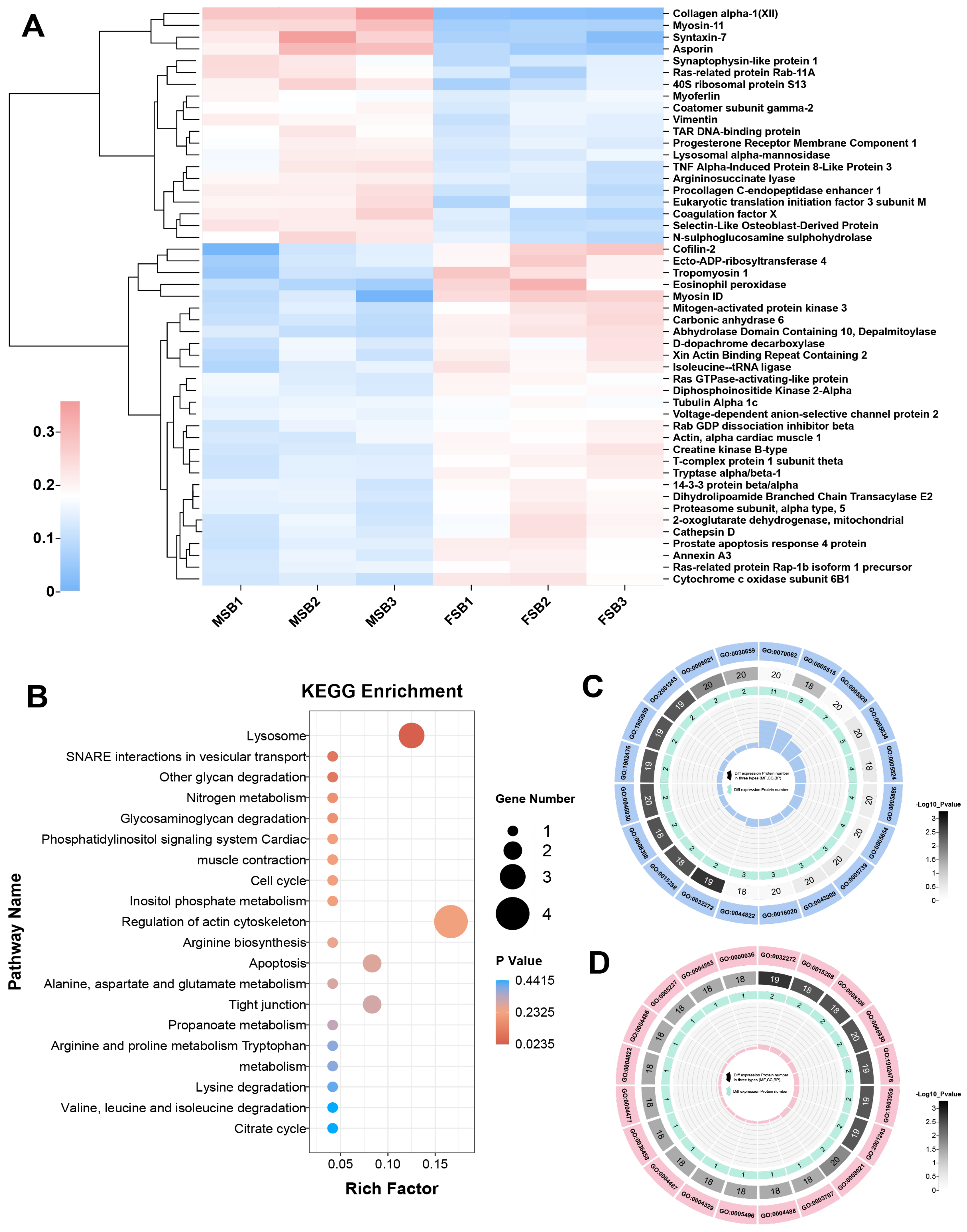
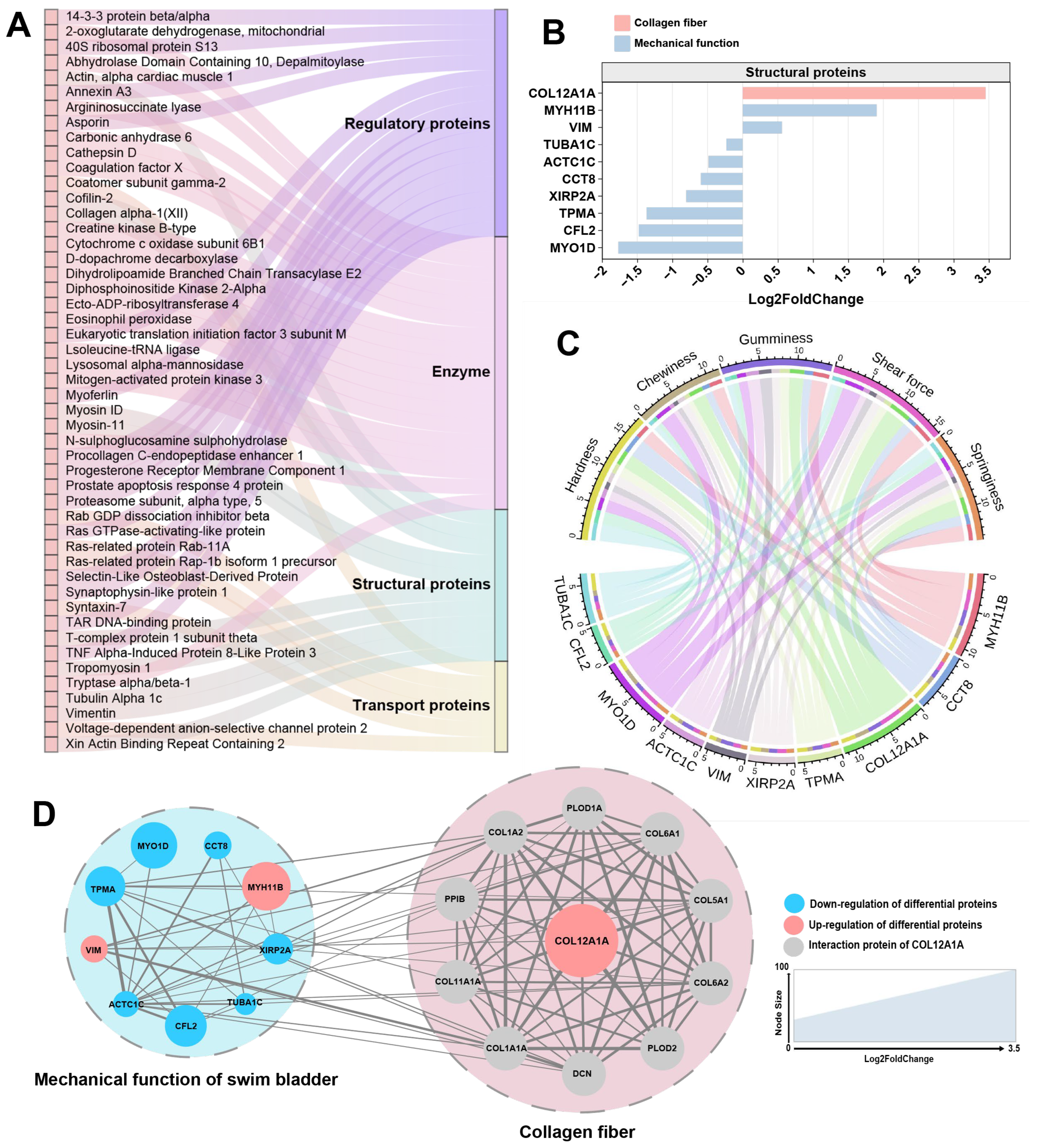
3.4. Proteomic Analysis of MSB and FSB
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hyde, R.J.; Witherly, S.A. Dynamic Contrast: A Sensory Contribution to Palatability. Appetite 1993, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Meilgaard, M.C.; Carr, B.T.; Civille, G.V. Sensory Evaluation Techniques, 3rd ed.; CRC Press: Boca Raton, FL, USA, 1999; ISBN 978-1-00-304072-9. [Google Scholar]
- Hagen, Ø.; Solberg, C.; Sirnes, E.; Johnston, I.A. Biochemical and Structural Factors Contributing to Seasonal Variation in the Texture of Farmed Atlantic Halibut (Hippoglossus Hippoglossus L.) Flesh. J. Agric. Food Chem. 2007, 55, 5803–5808. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.-H.; Sun, D.-W.; Han, Z.; Zeng, X.-A. Texture and Structure Measurements and Analyses for Evaluation of Fish and Fillet Freshness Quality: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 52–61. [Google Scholar] [CrossRef]
- Jiang, H.; Kong, Y.; Song, L.; Liu, J.; Wang, Z. A Thermostable Type I Collagen from Swim Bladder of Silver Carp (Hypophthalmichthys Molitrix). Mar. Drugs 2023, 21, 280. [Google Scholar] [CrossRef]
- Ramachandran, G.N.; Kartha, G. Structure of Collagen. Nature 1954, 174, 269–270. [Google Scholar] [CrossRef]
- Holmes, D.F.; Lu, Y.; Starborg, T.; Kadler, K.E. Collagen Fibril Assembly and Function. Curr. Top. Dev. Bio. 2018, 130, 107–142. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Ottani, V.; Raspanti, M.; Ruggeri, A. Collagen Structure and Functional Implications. Micron 2001, 32, 251–260. [Google Scholar] [CrossRef]
- Ramachandran, G. Stereochemistry of Collagen. Int. J. Pept. Protein Res. 1988, 31, 1–16. [Google Scholar] [CrossRef]
- Huang, Y.; Wen, X.; Li, S.; Xuan, X.; Zhu, D. Effects of Protein Levels on Growth, Feed Utilization, Body Composition, Amino Acid Composition and Physiology Indices of Juvenile Chu’s Croaker, Nibea Coibor. Aquac. Nutr. 2017, 23, 594–602. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, Q.; Liu, X.; Zhang, H.; Wang, S.; Wen, X.; Lin, F. Dietary Hydroxyproline Promotes Collagen Deposition in Swim Bladder through Regulating Biosynthesis of Amino Acid: In-Vitro and in-Vivo Investigations in Nibea Coibor. Aquaculture 2023, 573, 739614. [Google Scholar] [CrossRef]
- Lin, F.; Liu, X.; Guo, H.; Liu, G.; Ye, X.; Yuan, Y.; Lin, J.; Wang, S.; Wen, X. Sexual Dimorphism in the Texture Properties of Swim Bladder of Nibea Coibor and Its Molecular Basis Revealed by Comparative Transcriptome Analysis. Aquac. Rep. 2022, 25, 101207. [Google Scholar] [CrossRef]
- Barua, S.; Liu, Q.; Chen, X.; Eleneen, E.A.A. Stock and Market Status of High-Valued Blackspotted Croaker (Protonibea Diacanthus) in Bangladesh. Fish. Manag. Ecol. 2025, e12791. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2006. [Google Scholar]
- Yu, E.-M.; Zhang, H.-F.; Li, Z.-F.; Wang, G.-J.; Wu, H.-K.; Xie, J.; Yu, D.-G.; Xia, Y.; Zhang, K.; Gong, W.-B. Proteomic Signature of Muscle Fibre Hyperplasia in Response to Faba Bean Intake in Grass Carp. Sci. Rep. 2017, 7, 45950. [Google Scholar] [CrossRef]
- Chen, J.; Fan, Z.; Tan, D.; Jiang, D.; Wang, D. A Review of Genetic Advances Related to Sex Control and Manipulation in Tilapia. J. World Aquac. Soc. 2018, 49, 277–291. [Google Scholar] [CrossRef]
- Imsland, A.; Folkvord, A.; Grung, G.; Stefansson, S.; Taranger, G. Sexual Dimorphism in Growth and Maturation of Turbot, Scophthalmus Maximus (Rafinesque, 1810). Aquac. Res. 1997, 28, 101–114. [Google Scholar] [CrossRef]
- Saillant, E.; Fostier, A.; Menu, B.; Haffray, P.; Chatain, B. Sexual Growth Dimorphism in Sea Bass Dicentrarchus Labrax. Aquaculture 2001, 202, 371–387. [Google Scholar] [CrossRef]
- Ramcharitar, J.; Gannon, D.P.; Popper, A.N. Bioacoustics of Fishes of the Family Sciaenidae (Croakers and Drums). Trans. Am. Fish. Soc. 2006, 135, 1409–1431. [Google Scholar] [CrossRef]
- Pearce, K.L.; Rosenvold, K.; Andersen, H.J.; Hopkins, D.L. Water Distribution and Mobility in Meat during the Conversion of Muscle to Meat and Ageing and the Impacts on Fresh Meat Quality Attributes--a Review. Meat Sci. 2011, 89, 111–124. [Google Scholar] [CrossRef]
- Andersen, U.B.; Thomassen, M.S.; Rørå, A.M.B. Texture Properties of Farmed Rainbow Trout (Oncorhynchus Mykiss): Effects of Diet, Muscle Fat Content and Time of Storage on Ice. J. Sci. Food Agric. 1997, 74, 347–353. [Google Scholar] [CrossRef]
- Dransfield, E. Intramuscular Composition and Texture of Beef Muscles. J. Sci. Food Agric. 1977, 28, 833–842. [Google Scholar] [CrossRef]
- Lin, J.; Shi, Y.; Men, Y.; Wang, X.; Ye, J.; Zhang, C. Mechanical Roles in Formation of Oriented Collagen Fibers. Tissue Eng. Part B Rev. 2020, 26, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Ojima, K.; Liu, A.; Hattori, A.; Takahashi, K. Structural Changes in the Intramuscular Connective Tissue During Development of Bovine Semitendinosus Muscle. Tissue Cell 1996, 28, 527–536. [Google Scholar] [CrossRef]
- Light, N.; Champion, A.E.; Voyle, C.; Bailey, A.J. The Rôle of Epimysial, Perimysial and Endomysial Collagen in Determining Texture in Six Bovine Muscles. Meat Sci. 1985, 13, 137–149. [Google Scholar] [CrossRef]
- Edgardo Tapia-Vasquez, A.; Marina Ezquerra-Brauer, J.; Martinez-Cruz, O.; Marquez-Rios, E.; Enrique Ramirez-Guerra, H.; Minjarez-Osorio, C.; Torres-Arreola, W. Relationship between Muscle Texture and the Crosslinking Degree of Collagen Fibers from Octopus (Octopus Vulgaris), Guitarfish (Rhinobatos Productus), and Cazon (Mustelus Lunulatus). J. Food Process Preserv. 2019, 43, e14241. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Elango, J.; Hou, C.; Bao, B.; Wang, S.; Maté Sánchez de Val, J.E.; Wenhui, W. The Molecular Interaction of Collagen with Cell Receptors for Biological Function. Polymers 2022, 14, 876. [Google Scholar] [CrossRef]
- Niu, C.; Hu, Y.; Xu, K.; Pan, X.; Wang, L.; Yu, G. The Role of the Cytoskeleton in Fibrotic Diseases. Front. Cell Dev. Biol. 2024, 12, 1490315. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 Years and Still GOing Strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef]
- Obinata, T.; Maruyama, K.; Sugita, H.; Kohama, K.; Ebashi, S. Dynamic Aspects of Structural Proteins in Vertebrate Skeletal Muscle. Muscle Nerve 1981, 4, 456–488. [Google Scholar] [CrossRef]
- Walchli, C.; Koch, M.; Chiquet, M.; Odermatt, B.F.; Trueb, B. Tissue-Specific Expression of the Fibril-Associated Collagens XII and XIV. J. Cell Sci. 1994, 107 Pt 2, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.K.; Gerecke, D.R.; Dublet, B.; van der Rest, M.; Olsen, B.R. Type XII Collagen. A Large Multidomain Molecule with Partial Homology to Type IX Collagen. J. Biol. Chem. 1989, 264, 19772–19778. [Google Scholar] [CrossRef] [PubMed]
- Chiquet, M.; Birk, D.E.; Boennemann, C.G.; Koch, M. Collagen XII: Protecting Bone and Muscle Integrity by Organizing Collagen Fibrils. Int. J. Biochem. Cell Biol. 2014, 53, 51–54. [Google Scholar] [CrossRef]
- Zhang, G.; Young, B.B.; Birk, D.E. Differential Expression of Type XII Collagen in Developing Chicken Metatarsal Tendons. Am. J. Anat. 2003, 202, 411–420. [Google Scholar] [CrossRef]
- Schönborn, K.; Willenborg, S.; Schulz, J.-N.; Imhof, T.; Eming, S.A.; Quondamatteo, F.; Brinckmann, J.; Niehoff, A.; Paulsson, M.; Koch, M.; et al. Role of Collagen XII in Skin Homeostasis and Repair. Matrix Biol. 2020, 94, 57–76. [Google Scholar] [CrossRef]
- Delbaere, S.; Dhooge, T.; Syx, D.; Petit, F.; Goemans, N.; Destree, A.; Vanakker, O.; De Rycke, R.; Symoens, S.; Malfait, F. Novel Defects in Collagen XII and VI Expand the Mixed Myopathy/Ehlers-Danlos Syndrome Spectrum and Lead to Variant-Specific Alterations in the Extracellular Matrix. Genet. Med. 2020, 22, 112–123. [Google Scholar] [CrossRef]
- Chiquet, M.; Matthisson, M.; Koch, M.; Tannheimer, M.; Chiquet-Ehrismann, R. Regulation of Extracellular Matrix Synthesis by Mechanical Stress. Biochem. Cell Biol. 1996, 74, 737–744. [Google Scholar] [CrossRef]
- Jin, X.; Iwasa, S.; Okada, K.; Ooi, A.; Mitsui, K.; Mitsumata, M. Shear Stress-Induced Collagen XII Expression Is Associated with Atherogenesis. Biochem. Biophys. Res. Commun. 2003, 308, 152–158. [Google Scholar] [CrossRef]
- Nishiyama, T.; McDonough, A.M.; Bruns, R.R.; Burgeson, R.E. Type XII and XIV Collagens Mediate Interactions between Banded Collagen Fibers in Vitro and May Modulate Extracellular Matrix Deformability. J. Biol. Chem. 1994, 269, 28193–28199. [Google Scholar] [CrossRef]
- Trächslin, J.; Koch, M.; Chiquet, M. Rapid and Reversible Regulation of Collagen XII Expression by Changes in Tensile Stress. Exp. Cell Res. 1999, 247, 320–328. [Google Scholar] [CrossRef]
- Sun, M.; Zafrullah, N.; Devaux, F.; Hemmavanh, C.; Adams, S.; Ziebarth, N.M.; Koch, M.; Birk, D.E.; Espana, E.M. Collagen XII Is a Regulator of Corneal Stroma Structure and Function. Investig. Opthalmology Vis. Sci. 2020, 61, 61. [Google Scholar] [CrossRef] [PubMed]
- Moinzadeh, P.; Agarwal, P.; Bloch, W.; Orteu, C.; Hunzelmann, N.; Eckes, B.; Krieg, T. Systemic Sclerosis with Multiple Nodules: Characterization of the Extracellular Matrix. Arch. Dermatol. Res. 2013, 305, 645–652. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid (%) | MSB | FSB |
|---|---|---|
| Arg | 6.12 ± 0.62 | 6.76 ± 0.78 |
| His | 0.54 ± 0.04 | 0.59 ± 0.06 |
| Lys | 1.52 ± 0.10 | 1.68 ± 0.19 |
| Gly | 15.93 ± 0.86 | 15.64 ± 0.65 |
| Ser | 1.72 ± 0.04 | 1.80 ± 0.05 |
| Cys-Cys | 0.03 ± 0.00 | 0.03 ± 0.01 |
| Asp | 4.02 ± 0.10 | 3.87 ± 0.53 |
| Glu | 8.09 ± 0.37 | 8.24 ± 0.41 |
| Ala | 7.75 ± 0.25 | 7.63 ± 0.15 |
| Thr | 2.02 ± 0.12 | 2.09 ± 0.10 |
| Pro | 7.94 ± 0.36 | 7.89 ± 0.18 |
| Met | 1.63 ± 0.17 | 1.27 ± 0.80 |
| Val | 2.26 ± 0.16 | 1.88 ± 0.79 |
| Tyr | 0.73 ± 0.05 | 0.65 ± 0.13 |
| Ile | 0.82 ± 0.06 | 0.82 ± 0.15 |
| Leu | 1.85 ± 0.09 | 1.77 ± 0.47 |
| Phe | 1.63 ± 0.14 | 1.67 ± 0.22 |
| Hyp | 4.09 ± 0.09 | 4.52 ± 0.18 |
| Imino acid | 12.02 ± 0.27 | 12.42 ± 0.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Lin, J.; Guo, H.; He, X.; Jiang, W.; Yan, L.; Yan, K.; Wen, X.; Lin, F. Proteomics-Based Investigation of Sexual Dimorphism in Swim Bladder Texture of Chu’s Croaker (Nibea coibor). Foods 2025, 14, 1586. https://doi.org/10.3390/foods14091586
Zhang H, Lin J, Guo H, He X, Jiang W, Yan L, Yan K, Wen X, Lin F. Proteomics-Based Investigation of Sexual Dimorphism in Swim Bladder Texture of Chu’s Croaker (Nibea coibor). Foods. 2025; 14(9):1586. https://doi.org/10.3390/foods14091586
Chicago/Turabian StyleZhang, Haoran, Jiali Lin, Haoji Guo, Xianda He, Wanying Jiang, Lin Yan, Kuoqiu Yan, Xiaobo Wen, and Fan Lin. 2025. "Proteomics-Based Investigation of Sexual Dimorphism in Swim Bladder Texture of Chu’s Croaker (Nibea coibor)" Foods 14, no. 9: 1586. https://doi.org/10.3390/foods14091586
APA StyleZhang, H., Lin, J., Guo, H., He, X., Jiang, W., Yan, L., Yan, K., Wen, X., & Lin, F. (2025). Proteomics-Based Investigation of Sexual Dimorphism in Swim Bladder Texture of Chu’s Croaker (Nibea coibor). Foods, 14(9), 1586. https://doi.org/10.3390/foods14091586