
Exploring the Microbiome and Functional Metabolism of Fermented Camel Milk (Shubat) Using Metagenomics


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Total DNA Extraction
2.3. Library Construction and Illumina Sequencing
2.4. Data Quality Control and Assembly
2.5. Bioinformatics Analysis
2.6. Statistical Analysis
3. Results
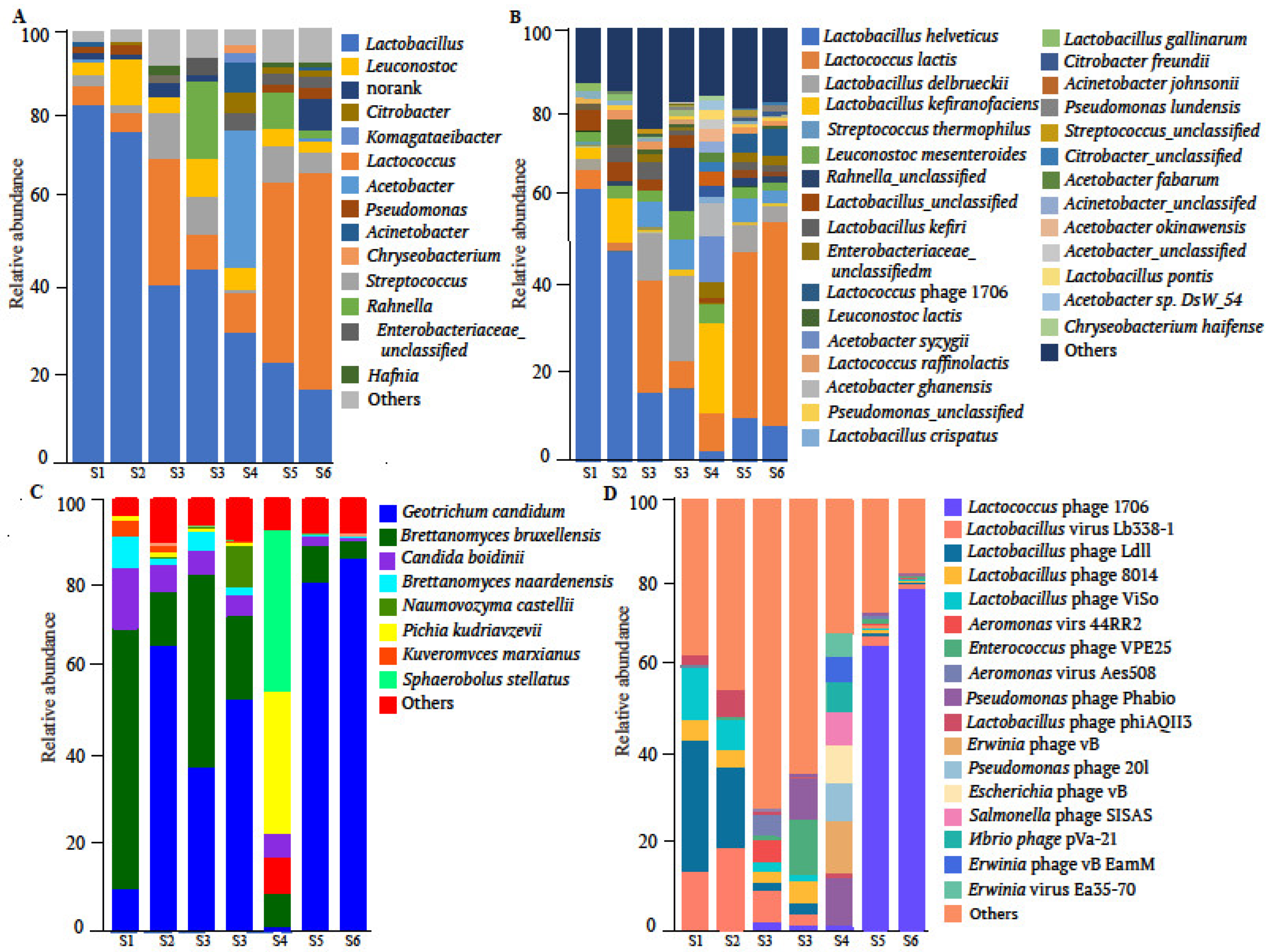
3.1. Species Annotation Results
3.2. Microbial Community Diversity Analysis
3.2.1. Alpha Diversity Analysis
3.2.2. Beta Diversity Analysis
3.3. Functional Annotation and Analysis
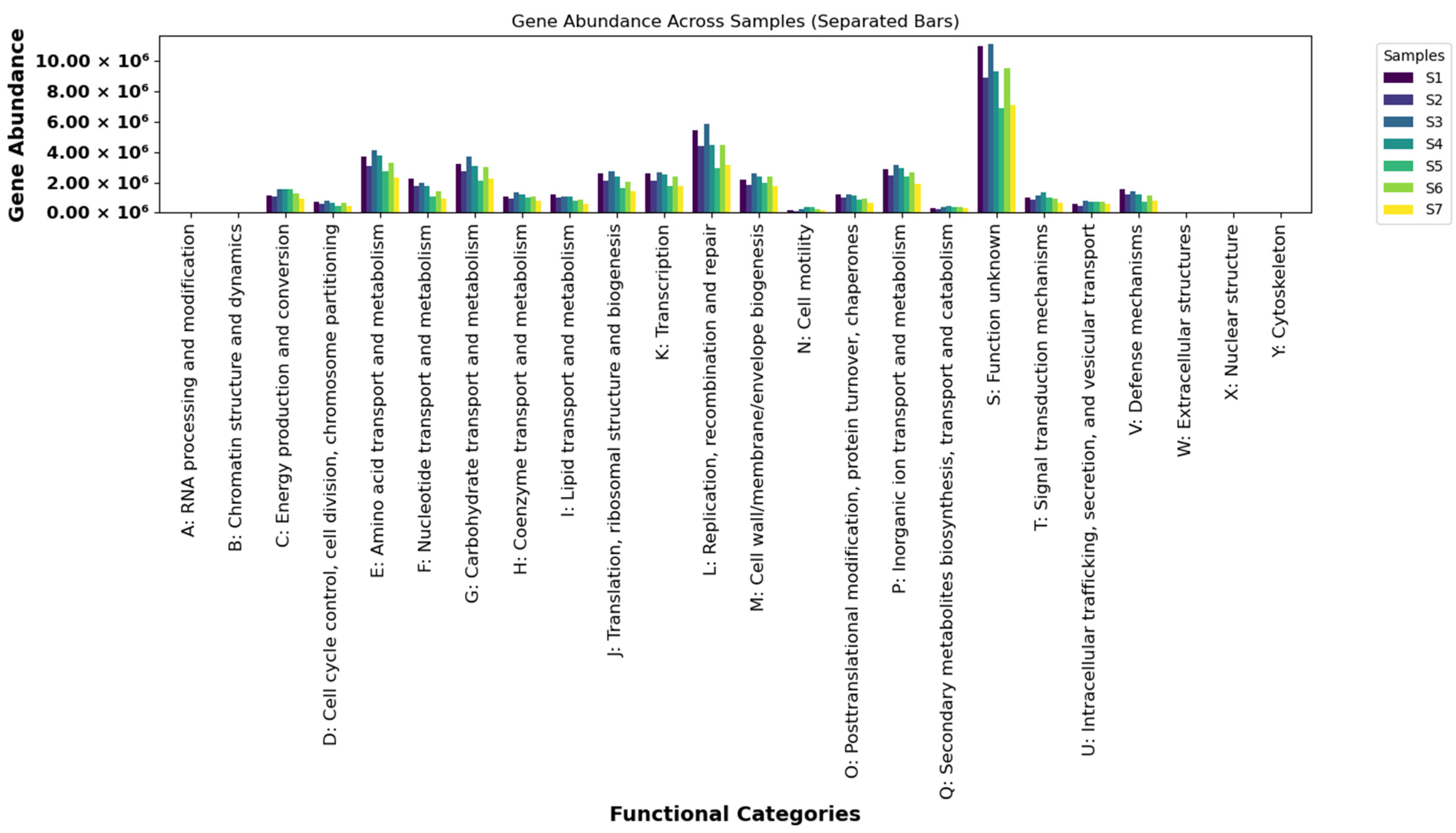
3.3.1. COG Functional Classification Analysis
3.3.2. KEGG Pathway Analysis
3.3.3. Carbohydrate-Active Enzyme (CAZy) Analysis
4. Discussion
4.1. Microbial Community Structure Analysis
4.1.1. Bacterial Diversity of Shubat
4.1.2. Dominant Fungal Species in Shubat
4.1.3. Viral Composition in Shubat
4.2. Functional Annotation Analysis
4.2.1. COG Functional Classification Analysis
4.2.2. KEGG Pathway Analysis
4.2.3. CAZy Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Izadi, A.; Khedmat, L.; Mojtahedi, S.Y. Nutritional and Therapeutic Perspectives of Camel Milk and Its Protein Hydrolysates: A Review on Versatile Biofunctional Properties. J. Funct. Foods 2019, 60, 103441. [Google Scholar]
- Salami, M.; Moosavi-Movahedi, A.A.; Moosavi-Movahedi, F.; Ehsani, M.R.; Yousefi, R.; Farhadi, M.; Niasari-Naslaji, A.; Saboury, A.A.; Chobert, J.M.; Haertlé, T. Biological Activity of Camel Milk Casein Following Enzymatic Digestion. J. Dairy Res. 2011, 78, 471–478. [Google Scholar] [CrossRef]
- Hailu, Y.; Hansen, E.B.; Seifu, E.; Eshetu, M.; Ipsen, R.; Kappeler, S. Functional and Technological Properties of Camel Milk Proteins: A Review. J. Dairy Res. 2016, 83, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N. Beneficial Plant-Microbe Interactions for Agricultural Sustainability. J. Appl. Biol. Biotechnol. 2021, 9, i–iv. [Google Scholar]
- Thapa, S.; Li, H.; OHair, J.; Bhatti, S.; Chen, F.-C.; Nasr, K.A.; Johnson, T.; Zhou, S. Biochemical Characteristics of Microbial Enzymes and Their Significance from Industrial Perspectives. Mol. Biotechnol. 2019, 61, 579–601. [Google Scholar] [PubMed]
- Sarıtaş, S.; Portocarrero, A.C.M.; Miranda López, J.M.; Lombardo, M.; Koch, W.; Raposo, A.; El-Seedi, H.R.; de Brito Alves, J.L.; Esatbeyoglu, T.; Karav, S. The Impact of Fermentation on the Antioxidant Activity of Food Products. Molecules 2024, 29, 3941. [Google Scholar] [CrossRef]
- Ağagündüz, D.; Şahin, T.Ö.; Ayten, Ş.; Yılmaz, B.; Güneşliol, B.E.; Russo, P.; Spano, G.; Özogul, F. Lactic Acid Bacteria as Pro-Technological, Bioprotective and Health-Promoting Cultures in the Dairy Food Industry. Food Biosci. 2022, 47, 101617. [Google Scholar] [CrossRef]
- Bao, W.; He, Y.; Yu, J.; Yang, X.; Liu, M.; Ji, R. Diversity analysis and gene function prediction of bacteria and fungi of Bactrian camel milk and naturally fermented camel milk from Alxa in Inner Mongolia. LWT 2022, 169, 114001. [Google Scholar]
- Baubekova, A.; Akhmetsadykova, S.; Konuspayeva, G.; Akhmetsadykov, N.; Faye, B.; Loiseau, G. Biodiversity study of the yeast in fresh and fermented camel and mare’s milk by denaturing gradient gel electrophoresis. J. Camel Pract. Res. 2015, 22, 91–95. [Google Scholar]
- Ledormand, P.; Desmasures, N.; Dalmasso, M. Phage community involvement in fermented beverages: An open door to technological advances? Crit. Rev. Food Sci. Nutr. 2021, 61, 2911–2920. [Google Scholar]
- Lema, N.K.; Gemeda, M.T.; Woldesemayat, A.A. Recent Advances in Metagenomic Approaches, Applications, and Challenge. Curr. Microbiol. 2023, 80, 347. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Napoli, S.; Alessandri, G.; Mancabelli, L.; Anzalone, R.; Longhi, G.; Viappiani, A.; Mangifesta, M.; Lugli, G.A.; et al. Colonization of the Human Gut by Bovine Bacteria Present in Parmesan Cheese. Nat. Commun. 2019, 10, 1286. [Google Scholar] [CrossRef]
- Quijada, N.M.; Mann, E.; Wagner, M.; Rodríguez-Lázaro, D.; Hernández, M.; Schmitz-Esser, S. Autochthonous Facility-Specific Microbiota Dominates Washed-Rind Austrian Hard Cheese Surfaces and Its Production Environment. Int. J. Food Microbiol. 2018, 267, 54–61. [Google Scholar]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating Viruses and Cellular Organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Cantarel, B.I.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes Database (CAZy): An Expert Resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- Du, R.; Xiong, W.; Xu, L.; Xu, Y.; Wu, Q. Metagenomics reveals the habitat specificity of biosynthetic potential of secondary metabolites in global food fermentations. Microbiome 2023, 11, 115. [Google Scholar] [CrossRef]
- Crauwels, S.; Van Opstaele, F.; Jaskula-Goiris, B.; Steensels, J.; Verreth, C.; Bosmans, L.; Paulussen, C.; Herrera-Malaver, B.; de Jonge, R.; De Clippeleer, J. Fermentation Assays Reveal Differences in Sugar and (off-) Flavor Metabolism across Different Brettanomyces Bruxellensis Strains. Yeast Res. 2017, 17, fow105. [Google Scholar]
- Zhadyra, S.; Han, X.; Anapiyayev, B.B.; Tao, F.; Xu, P. Bacterial Diversity Analysis in Kazakh Fermented Milks Shubat and Ayran by Combining Culture-Dependent and Culture-Independent Methods. LWT 2021, 141, 110877. [Google Scholar] [CrossRef]
- Chelladhurai, K.; Ayyash, M.; Turner, M.S.; Kamal-Eldin, A. Lactobacillus helveticus: Health effects, current applications, and future trends in dairy fermentation. Trends Food Sci. Technol. 2023, 136, 159–168. [Google Scholar]
- Ghosh, T.; Beniwal, A.; Semwal, A.; Navani, N.K. Mechanistic Insights into Probiotic Properties of Lactic Acid Bacteria Associated with Ethnic Fermented Dairy Products. Front. Microbiol. 2019, 10, 502. [Google Scholar]
- Sharma, H.; Ozogul, F.; Bartkiene, E.; Rocha, J.M. Impact of lactic acid bacteria and their metabolites on the techno-functional properties and health benefits of fermented dairy products. Crit. Rev. Food Sci. Nutr. 2023, 63, 4819–4841. [Google Scholar] [CrossRef]
- You, L.; Yang, C.; Jin, H.; Kwok, L.-Y.; Lv, R.; Ma, T.; Zhao, Z.; Zhang, H.; Sun, Z. Shotgun Metagenomic Analysis of Microbiota Dynamics during Long-Term Backslopping Fermentation of Traditional Fermented Milk in a Controlled Laboratory Environment. J. Dairy. Sci. 2024, 107, 7619–7630. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Ma, H.; Hou, Q.; Li, W.; Xu, H.; Liu, W.; Sun, Z.; Haobisi, H.; Menghe, B. Profiling of Koumiss Microbiota and Organic Acids and Their Effects on Koumiss Taste. BMC Microbiol. 2020, 20, 85. [Google Scholar] [CrossRef] [PubMed]
- Jokovic, N.; Nikolic, M.; Begovic, J.; Jovcic, B.; Savic, D.; Topisirovic, L. A Survey of the Lactic Acid Bacteria Isolated from Serbian Artisanal Dairy Product Kajmak. Int. J. Food Microbiol. 2008, 127, 305–311. [Google Scholar] [CrossRef]
- Tournay, R.J.; Firrincieli, A.; Parikh, S.S.; Sivitilli, D.M.; Doty, S.L. Effect of Arsenic on EPS Synthesis, Biofilm Formation, and Plant Growth-Promoting Abilities of the Endophytes Pseudomonas PD9R and Rahnella Laticis PD12R. Environ. Sci. Technol. 2023, 57, 8728–8738. [Google Scholar] [CrossRef] [PubMed]
- Baschali, A.; Tsakalidou, E.; Kyriacou, A.; Karavasiloglou, N.; Matalas, A.-L. Traditional Low-Alcoholic and Non-Alcoholic Fermented Beverages Consumed in European Countries: A Neglected Food Group. Nutr. Res. Rev. 2017, 30, 1–24. [Google Scholar] [CrossRef]
- Ekawati, N.; Mutiara, I.; Hertati, A.; Kusdianawati; Mustopa, A.Z.; Fatimah; Manguntungi, B.; Elviantari, A. Biodiversity and Probiotic Potential of Yeasts Isolated from Sumbawa Horse Milk. Mol. Biol. Rep. 2024, 51, 911. [Google Scholar] [CrossRef]
- Eliskases-Lechner, F.; Guéguen, M.; Panoff, J.M. Yeasts and Molds|Geotrichum Candidum. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 765–771. ISBN 978-0-12-374407-4. [Google Scholar]
- Rahman, N.; Xiaohong, C.; Meiqin, F.; Mingsheng, D. Characterization of the Dominant Microflora in Naturally Fermented Camel Milk Shubat. World J. Microbiol. Biotechnol. 2009, 25, 1941–1946. [Google Scholar] [CrossRef]
- Akhmetsadykova, S.; Baubekova, A.; Konuspayeva, G.; Akhmetsadykov, N.; Loiseau, G. Microflora Identification of Fresh and Fermented Camel Milk from Kazakhstan. Emir. J. Food Agric. (EJFA) 2014, 26, 327–332. [Google Scholar] [CrossRef]
- Garneau, J.E.; Tremblay, D.M.; Moineau, S. Characterization of 1706, a Virulent Phage from Lactococcus Lactis with Similarities to Prophages from Other Firmicutes. Virology 2008, 373, 298–309. [Google Scholar] [CrossRef]
- Moineau, S.; Lévesque, C. Control of Bacteriophages in Industrial Fermentations. In Bacteriophages: Biology and Applications; CRC Press: Boca Raton, FL, USA, 2005; pp. 285–296. [Google Scholar]
- Marcó, M.B.; Moineau, S.; Quiberoni, A. Bacteriophages and Dairy Fermentations. Bacteriophage 2012, 2, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, Y.; Gesudu, Q.; Zhang, J.; Sun, Z.; Halatu, H.; Menghe, B.; Liu, W. Bacterial Composition and Function during Fermentation of Mongolia Koumiss. Food Sci. Nutr. 2021, 9, 4146–4155. [Google Scholar] [CrossRef]
- Kumar, J.; Sharma, N.; Kaushal, G.; Samurailatpam, S.; Sahoo, D.; Rai, A.K.; Singh, S.P. Metagenomic Insights Into the Taxonomic and Functional Features of Kinema, a Traditional Fermented Soybean Product of Sikkim Himalaya. Front. Microbiol. 2019, 10, 1744. [Google Scholar] [CrossRef]
- Kobayashi, S.; Kawaguchi, H.; Shirai, T.; Ninomiya, K.; Takahashi, K.; Kondo, A.; Tsuge, Y. Automatic Redirection of Carbon Flux between Glycolysis and Pentose Phosphate Pathway Using an Oxygen-Responsive Metabolic Switch in Corynebacterium Glutamicum. ACS Synth. Biol. 2020, 9, 814–826. [Google Scholar] [CrossRef]
- Zhang, R.; Jia, W. Brown Goat Yogurt: Metabolomics, Peptidomics, and Sensory Changes during Production. J. Dairy. Sci. 2023, 106, 1712–1733. [Google Scholar] [CrossRef] [PubMed]
- Bourrie, B.C.T.; Willing, B.P.; Cotter, P.D. The Microbiota and Health Promoting Characteristics of the Fermented Beverage Kefir. Front. Microbiol. 2016, 7, 196946. [Google Scholar]
- Walsh, A.M.; Crispie, F.; Kilcawley, K.; O’Sullivan, O.; O’Sullivan, M.G.; Claesson, M.J.; Cotter, P.D. Microbial Succession and Flavor Production in the Fermented Dairy Beverage Kefir. mSystems 2016, 1, e00052-16. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- Valentino, V.; Magliulo, R.; Farsi, D.; Cotter, P.D.; O’Sullivan, O.; Ercolini, D.; De Filippis, F. Fermented Foods, Their Microbiome and Its Potential in Boosting Human Health. Microb. Biotechnol. 2024, 17, e14428. [Google Scholar]
- Gomes Soares, M.; Bevilaqua, G.C.; Marcondes Tassi, É.M.; Reolon Schmidt, V.C. Fermented Foods and Beverages: A Potential in Situ Vitamin B12 Biofortification—A Literature Review. Int. J. Food Sci. Nutr. 2023, 74, 655–667. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, U.; Tiwari, A.; Tiwari, P.; Sahu, J.K.; Sharma, S. Vitamin B12: Strategies for Enhanced Production, Fortified Functional Food Products and Health Benefits. Process Biochem. 2023, 127, 44–55. [Google Scholar]
- Wu, Z.; Liang, X.; Li, M.; Ma, M.; Zheng, Q.; Li, D.; An, T.; Wang, G. Advances in the Optimization of Central Carbon Metabolism in Metabolic Engineering. Microb. Cell Fact. 2023, 22, 76. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Jiang, T.; Liang, Z.; Zhang, N.; Dong, B.; Wu, Q.; Gu, B. Carbohydrate-Active Enzyme Profiles of Lactiplantibacillus Plantarum Strain 84-3 Contribute to Flavor Formation in Fermented Dairy and Vegetable Products. Food Chem. X 2023, 20, 101036. [Google Scholar]
- Shen, J.; Zheng, L.; Chen, X.; Han, X.; Cao, Y.; Yao, J. Metagenomic Analyses of Microbial and Carbohydrate-Active Enzymes in the Rumen of Dairy Goats Fed Different Rumen Degradable Starch. Front. Microbiol. 2020, 11, 1003. [Google Scholar]
- Kfoury, B.; Rodrigues, W.F.C.; Kim, S.-J.; Brandizzi, F.; Del-Bem, L.-E. Multiple Horizontal Gene Transfer Events Have Shaped Plant Glycosyl Hydrolase Diversity and Function. New Phytol. 2024, 242, 809–824. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Y.; Liaqat, F.; Khazi, M.I.; Sethupathy, S.; Zhu, D. Utilization of Glycosyltransferases as a Seamless Tool for Synthesis and Modification of the Oligosaccharides-A Review. Int. J. Biol. Macromol. 2023, 249, 125916. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, H.; Yu, Z.; Luo, X.; Zhao, J.; Wang, N.; Li, Z. Metagenomics-Based Gene Exploration and Biochemical Characterization of Novel Glucoamylases and α-Amylases in Daqu and Pu-Erh Tea Microorganisms. Int. J. Biol. Macromol. 2024, 278, 134182. [Google Scholar]
- Gong, G.; Zhou, S.; Luo, R.; Gesang, Z.; Suolang, S. Metagenomic Insights into the Diversity of Carbohydrate-Degrading Enzymes in the Yak Fecal Microbial Community. BMC Microbiol. 2020, 20, 302. [Google Scholar]
- Neves, A.L.A.; Yu, J.; Suzuki, Y.; Baez-Magana, M.; Arutyunova, E.; O’Hara, E.; McAllister, T.; Ominski, K.H.; Lemieux, M.J.; Guan, L.L. Accelerated Discovery of Novel Glycoside Hydrolases Using Targeted Functional Profiling and Selective Pressure on the Rumen Microbiome. Microbiome 2021, 9, 229. [Google Scholar]
- Li, L.-L.; McCorkle, S.R.; Monchy, S.; Taghavi, S.; van der Lelie, D. Bioprospecting Metagenomes: Glycosyl Hydrolases for Converting Biomass. Biotechnol. Biofuels 2009, 2, 10. [Google Scholar]
- Mareček, F.; Terrapon, N.; Janeček, Š. Two Newly Established and Mutually Related Subfamilies GH13_48 and GH13_49 of the α-Amylase Family GH13. Appl. Microbiol. Biotechnol. 2024, 108, 415. [Google Scholar]
- Korczynska, J.E.; Danielsen, S.; Schagerlöf, U.; Turkenburg, J.P.; Davies, G.J.; Wilson, K.S.; Taylor, E.J. The Structure of a Family GH25 Lysozyme from Aspergillus Fumigatus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 973–977. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modules | S1 | S2 | S3 | S4 | S5 | S6 | S7 | Definition |
|---|---|---|---|---|---|---|---|---|
| M00126 | 87,186 | 87,941 | 79,462 | 84,788 | 65,367 | 63,582 | 56,931 | Folate biosynthesis (genes per million reads) |
| M00114 | 40,426 | 41,904 | 41,105 | 40,314 | 42,411 | 36,005 | 33,733 | Ascorbic acid biosynthesis (plant pathway) |
| M00125 | 12,642 | 25,768 | 33,743 | 36,125 | 38,846 | 31,054 | 31,515 | Riboflavin biosynthesis |
| M00119 | 17,479 | 24,192 | 29,167 | 29,532 | 21,888 | 27,604 | 25,812 | Pantothenic acid biosynthesis |
| M00129 | 29,475 | 28,593 | 24,405 | 27,100 | 16,887 | 20,047 | 17,807 | Ascorbic acid biosynthesis (animal pathway) |
| M00124 | 14,037 | 14,364 | 15,664 | 19,006 | 15,394 | 15,746 | 15,714 | Pyridoxal biosynthesis |
| M00122 | 1831 | 10,572 | 13,428 | 5385 | 11,303 | 7824 | 6933 | Cobalamin (vitamin B12) biosynthesis |
| M00123 | 711 | 1147 | 2818 | 6537 | 11,655 | 3079 | 3000 | Biotin biosynthesis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhadyra, S.; Tao, F.; Xu, P. Exploring the Microbiome and Functional Metabolism of Fermented Camel Milk (Shubat) Using Metagenomics. Foods 2025, 14, 1102. https://doi.org/10.3390/foods14071102
Zhadyra S, Tao F, Xu P. Exploring the Microbiome and Functional Metabolism of Fermented Camel Milk (Shubat) Using Metagenomics. Foods. 2025; 14(7):1102. https://doi.org/10.3390/foods14071102
Chicago/Turabian StyleZhadyra, Sagyman, Fei Tao, and Ping Xu. 2025. "Exploring the Microbiome and Functional Metabolism of Fermented Camel Milk (Shubat) Using Metagenomics" Foods 14, no. 7: 1102. https://doi.org/10.3390/foods14071102
APA StyleZhadyra, S., Tao, F., & Xu, P. (2025). Exploring the Microbiome and Functional Metabolism of Fermented Camel Milk (Shubat) Using Metagenomics. Foods, 14(7), 1102. https://doi.org/10.3390/foods14071102