
Effect of Freeze Drying and Hot Air Drying on the Composition and Bioactivities of Lipids from Razor Clam Sinonovacula constricta

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Regents
2.2. Drying and Extraction of Lipid Samples
2.3. Lipid Pre-Treatment
2.4. Lipid Composition Analysis
2.5. Zebrafish Maintenance and Larvae Collection
2.6. Antithrombotic Assay
2.7. Anti-Inflammatory Assay
2.8. Antioxidant Assay
2.9. Statistical Analysis
3. Results
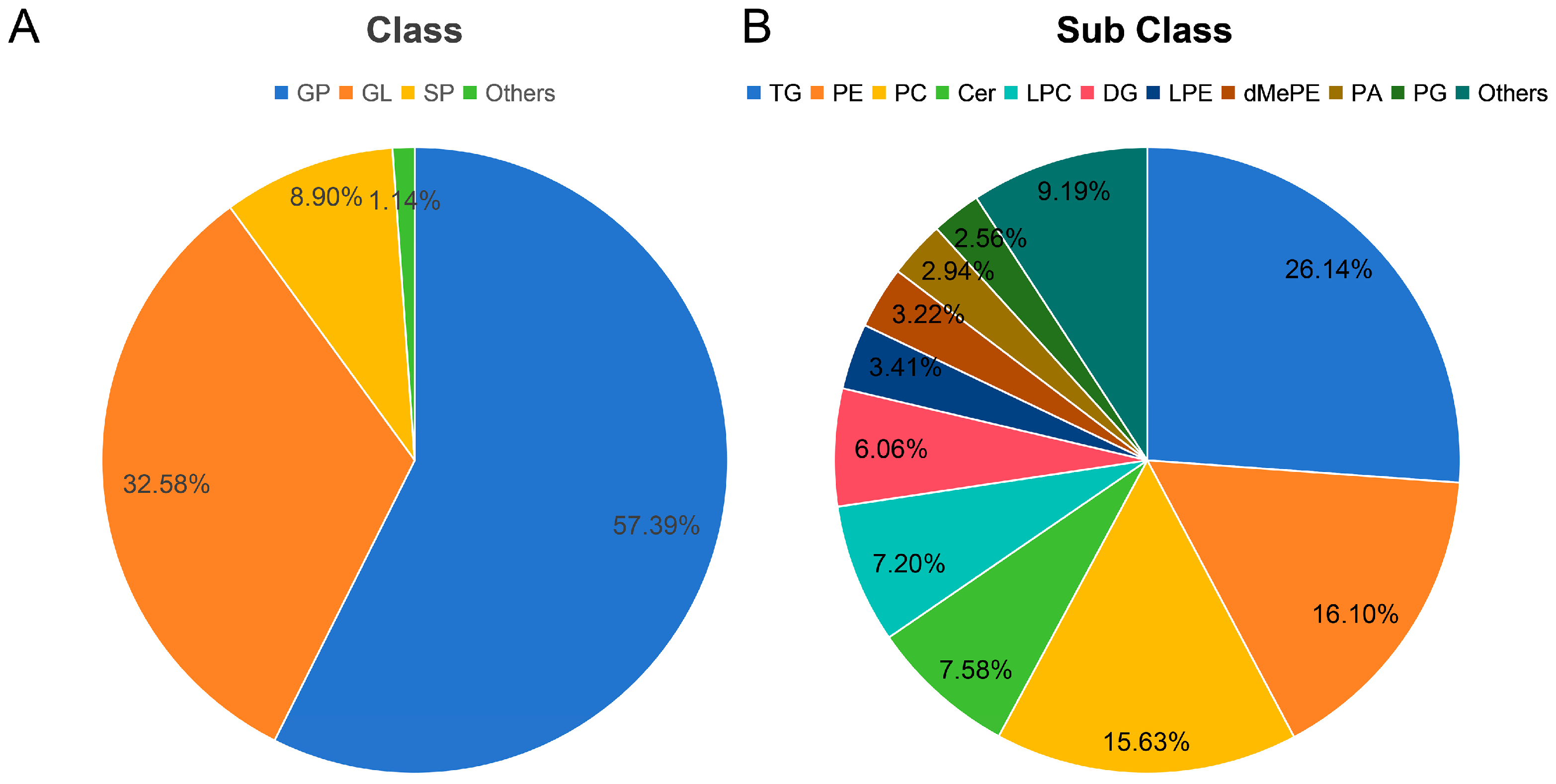
3.1. Lipidomic Analysis
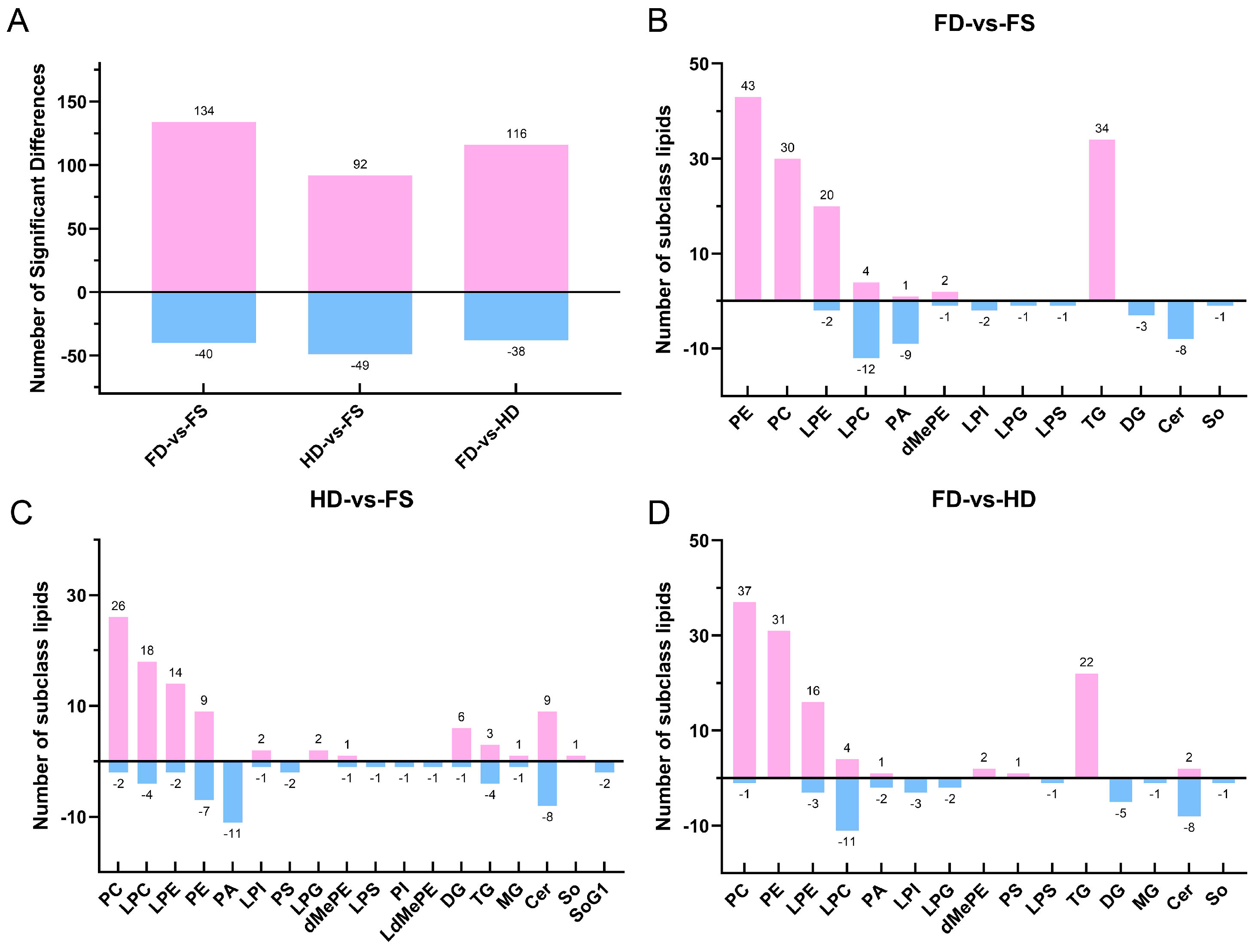
3.2. Comparative Analysis of Differences in Lipids
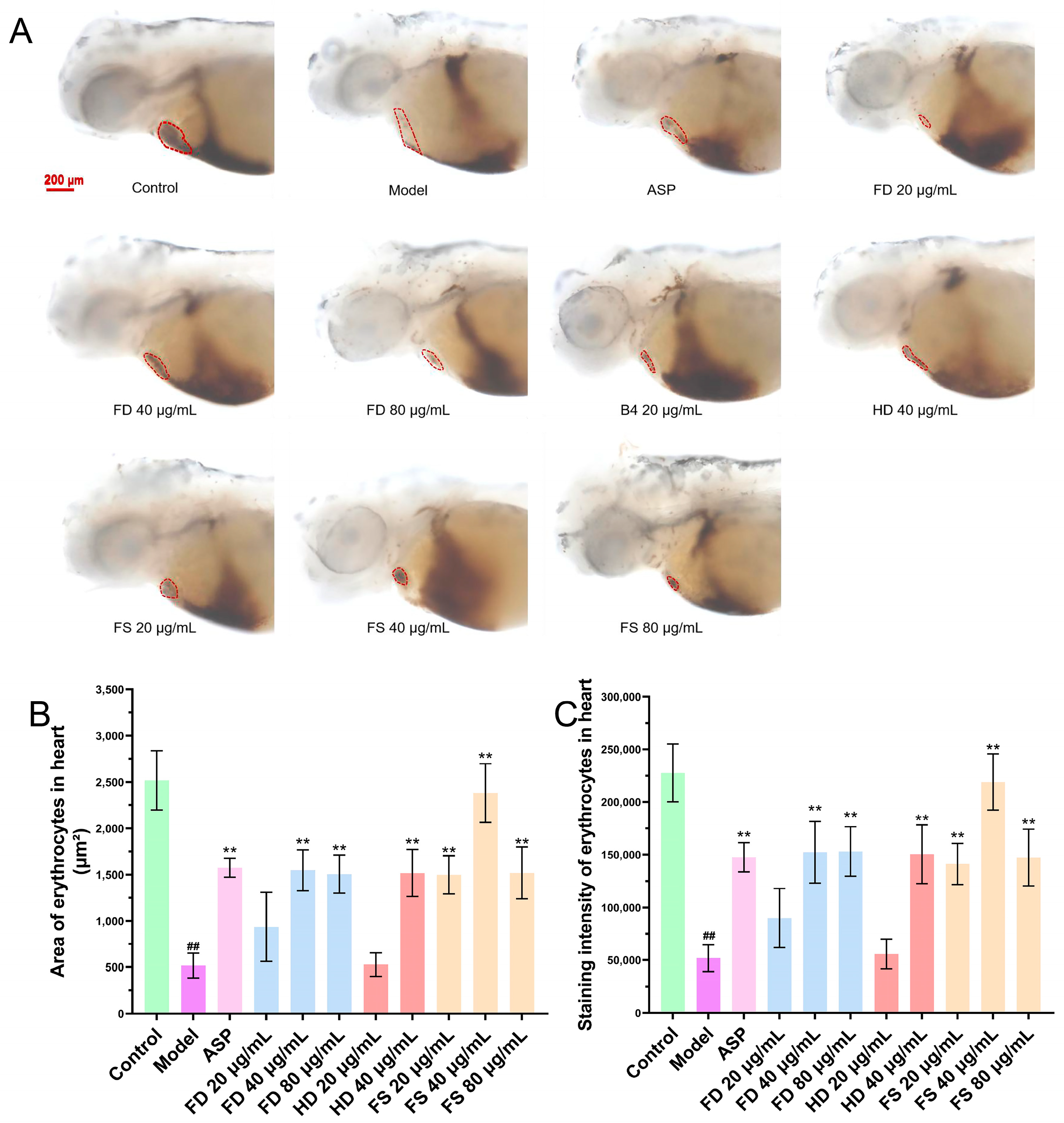
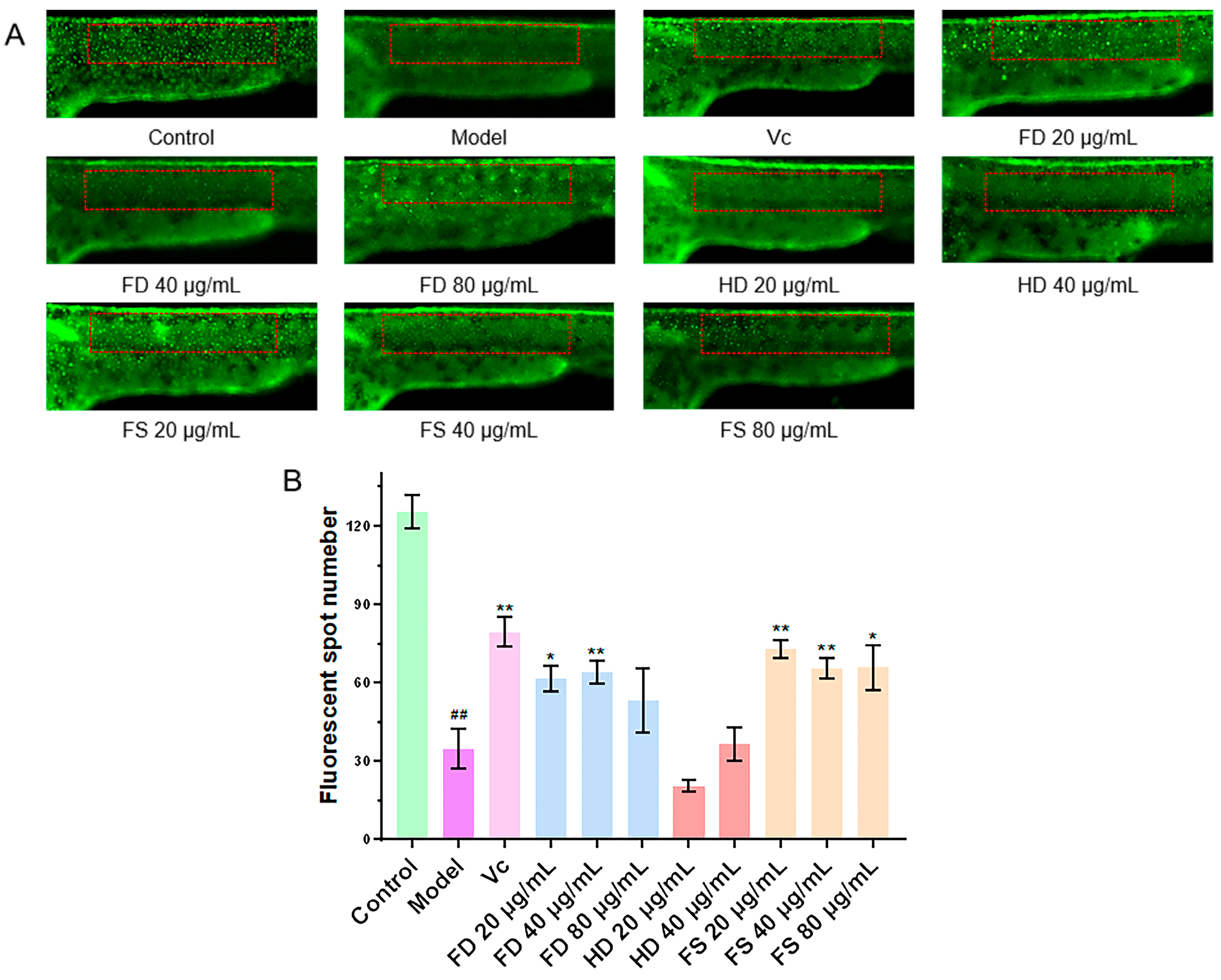
3.3. Evaluation of the Antithrombotic Activity of Lipids
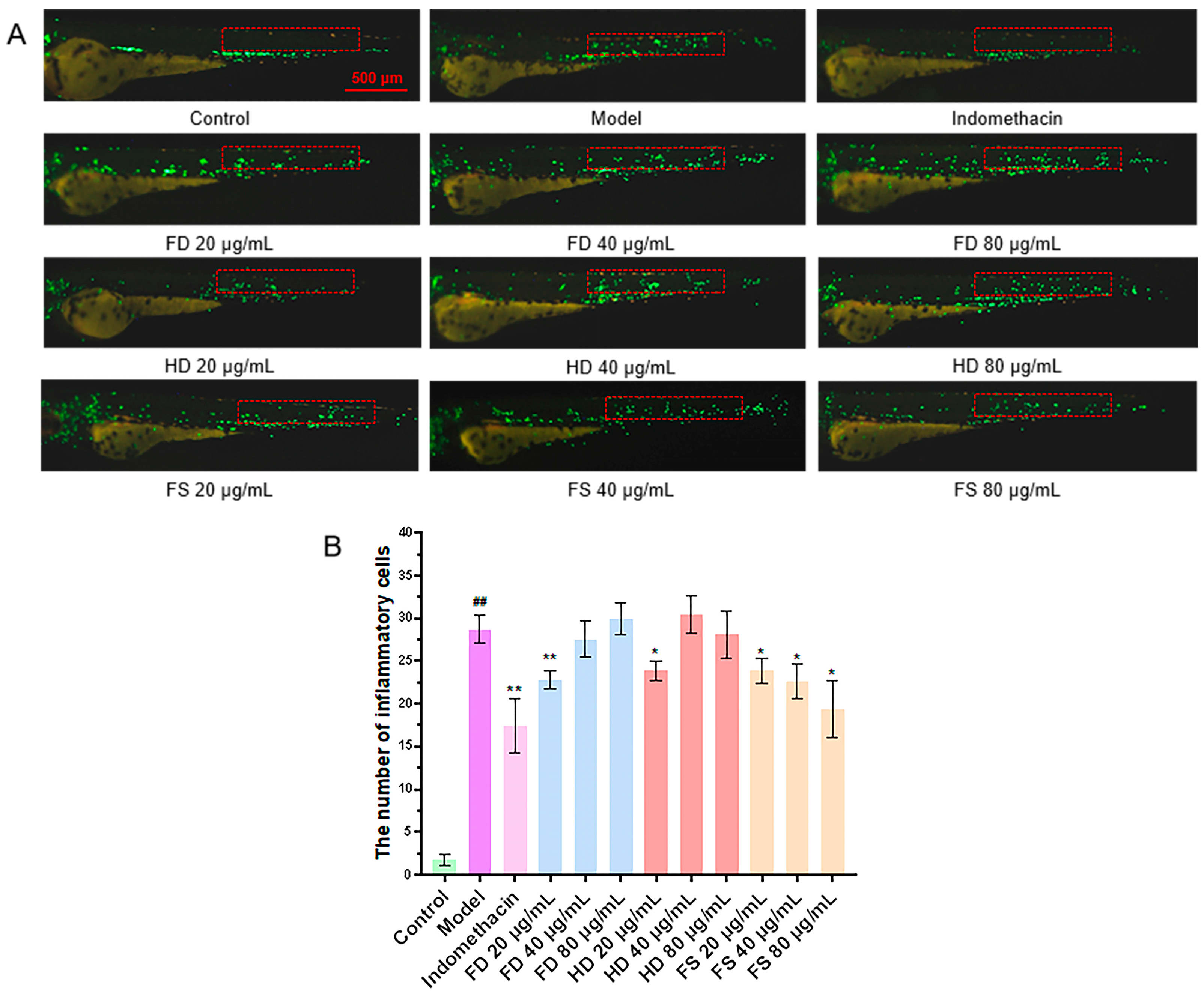
3.4. Evaluation of Anti-Inflammatory Activity of Lipids
3.5. Evaluation of Antioxidant Activity of Lipids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, Y.; Li, Q.; Kong, L.; Yu, H. The Complete Mitochondrial Genome of the Grand Jackknife Clam, Solen grandis (Bivalvia: Solenidae): A Novel Gene Order and Unusual Non-coding Region. Mol. Biol. Rep. 2012, 39, 1287–1292. [Google Scholar]
- Wei, X.; Yang, J.; Yang, D.; Xu, J.; Liu, X.; Yang, J.; Fang, J.; Qiao, H. Molecular Cloning and mRNA Expression of Two Peptidoglycan Recognition Protein (PGRP) Genes from Mollusk Solen grandis. Fish Shellfish Immunol. 2012, 32, 178–185. [Google Scholar] [CrossRef]
- Liu, Y.F. Analysis and Research on Lipid Fatty Acid Composition of Shellfish Foods Such as Sinonovacula constricta. Fish. Sci. Technol. Inf. 1991, 06, 179–180. [Google Scholar]
- Ran, Z.; Chen, H.; Ran, Y.; Yu, S.; Li, S.; Xu, J.; Liao, K.; Yu, X.; Zhong, Y.; Ye, M.; et al. Fatty Acid and Sterol Changes in Razor Clam Sinonovacula constricta (Lamarck 1818) Reared at Different Salinities. Aquaculture 2017, 473, 493–500. [Google Scholar] [CrossRef]
- Whittle, D.P.; Evans, D.J.; Scully, D.B.; Wood, G.C. Composition Changes and Oxidation Development. Acta Metall. 1967, 15, 1421–1430. [Google Scholar] [CrossRef]
- Yu, D.; Fu, Z.Y.; Li, L.; Liu, Y.J.; Yang, X.R.; Bai, Y.A.; Wang, Z.S.; Zheng, J. Study on the Nutritional Composition, Taste Characteristics and in vitro Antioxidant Activity of Solen grandis. Chin. Condiment 2024, 49, 49–56. [Google Scholar]
- China Market Research Network. Market Research and Development Trend Report on the Global and Chinese Solen Industry from 2023 to 2029; China Market Research Network: Guangzhou, China, 2025. [Google Scholar]
- Qi, Y.L.; Li, H.; Qiu, W.W.; Wang, Z.J. Study on Anhydrous Keeping Alive Technology of Sinonovacula constricta. Refrig. Technol. 2016, 157, 17–21. [Google Scholar]
- Vega–Gálvez, A.; Miranda, M.; Clavería, R.; Quispe, I.; Vergara, J.; Uribe, E.; Paez, H.; Di Scala, K. Effect of Air Temperature on Drying Kinetics and Quality Characteristics of Osmo–treated Jumbo Squid (Dosidicus gigas). LWT Food Sci. Technol. 2011, 44, 16–23. [Google Scholar] [CrossRef]
- Xie, H.; Yin, F.; Liu, Z.; Hu, Y.; Yu, M.; Zhou, D.; Zhu, B. Oxidation Kinetics of Polyunsaturated Fatty Acids Esterified into Triacylglycerols and Phospholipids in Dried Scallop (Argopecten irradians) Adductor Muscles during Storage. Food Funct. 2020, 11, 2349–2357. [Google Scholar] [PubMed]
- Sang, W.G.; Dong, M.M.; Tong, G.; Wang, L. Development of Seasoned Semi–dried Products of Cultured Sinonovacula constricta. Food Sci. Technol. 2008, 10, 119–122. [Google Scholar]
- Gang, K.Q.; Wu, Z.X.; Zhou, D.Y.; Zhao, Q.; Zhou, X.; Lv, D.D.; Rakariyatham, K.; Liu, X.Y.; Shahidi, F. Effects of Hot Air Drying Process on Lipid Quality of Whelks Neptunea arthritica cumingi Crosse and Neverita didyma. J. Food Sci. Technol. 2019, 56, 4166–4176. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, M.; Mujumdar, A.S. Trends in Processing Technologies for Dried Aquatic Products. Dry Technol. 2011, 29, 382–394. [Google Scholar] [CrossRef]
- Jiang, P.; Miao, X.; Li, J.; Qi, H.; Shang, S.; Dong, X. Volatile flavor characteristics of scallops (Chlamys farreri) with different drying methods were analyzed based on GC-IMS and GC-O-QTOF. Food Chem. X 2024, 24, 101960. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Fei, X.; Guo, S.; Du, M. Effects of drying methods on physical and chemical quality of walnut kernel and walnut oil. China Oils Fats 2019, 44, 8–13. [Google Scholar]
- Wang, B.; Liu, Y.; Dong, M.; Zhang, Y.; Huang, X.; Qin, L. Flavor enhancement during the drying of scallop (Patinopecten yessoensis) as revealed by integrated metabolomic and lipidomic analysis. Food Chem. 2024, 432, 137218. [Google Scholar] [CrossRef]
- NY/T 1712-2009; Green Food-Dried Aquatic Products. China Agricultural Press: Beijing, China, 2009.
- Sun, T.; Xue, C.; Xue, Y.; Zuo, T.; Yue, R. Comparison of Lipid Extraction Methods for Antarctic Krill. Sci. Technol. Food Ind. 2012, 16, 115–117+121. [Google Scholar]
- Li, P.; Xie, D.; Chen, H.; Qiu, Y.; Zhang, X.; Zhang, S.; Wang, L.; Lin, H.; Li, X.; Liu, K. Secondary metabolites from marine derived fungus Penicillium chrysogenum Y19-1 with proangiogenic and antithrombotic activities. Biochem. Syst. Ecol. 2023, 107. [Google Scholar] [CrossRef]
- Li, P.; Zhang, M.; Xie, D.; Zhang, X.; Zhang, S.; Gao, F.; Wang, Y.; Hsiao, C.; Li, X.; Liu, K. Characterization and bioactivities of phospholipids from squid viscera and gonads using ultra-performance liquid chromatography-Q-exactive orbitrap/mass spectrometry-based lipidomics and zebrafish models. Food Funct. 2021, 12, 7986–7996. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, M.; Li, H.; Wang, R.; Hou, H.; Li, X.; Liu, K.; Chen, H. New Prenylated Indole Homodimeric and Pteridine Alkaloids from the Marine-Derived Fungus Aspergillus austroafricanus Y32-2. Mar. Drugs 2021, 19, 98. [Google Scholar] [CrossRef]
- Liao, Y.; Ding, Y.; Du, Q.; Wu, Y.; Lin, H.; Benjakul, S.; Zhang, B. Changes in the lipid profiles of hairtail (Trichiurus lepturus) muscle during air-drying via liquid chromatography-mass spectrometry analysis. Food Chem. X 2023, 24, 100610. [Google Scholar] [CrossRef]
- Xie, H.K.; Zhao, G.H.; Wu, Z.X.; Li, D.Y.; Zhao, M.T.; Li, A.; Liu, H.L.; Zhou, D.Y.; Zhu, B.W. Differences in oxidative susceptibilities between glycerophosphocholine and glycerophosphoethanolamine in dried scallop (Argopecten irradians) adductor muscle during storage: An oxidation kinetic assessment. J. Sci. Food Agric. 2021, 101, 1554–1561. [Google Scholar] [CrossRef]
- Jia, M.; Cong, H.H.; Xue, C.H.; Xue, Y.; Sun, Z.M.; Li, J.Z. Abalone Hot Air Drying Kinetics and Drying Process Mathematical Simulation. Food Ind. Sci. Technol. 2012, 33, 72–76+80. [Google Scholar]
- Züllig, T.; Trötzmüller, M.; Köfeler, H.C. Lipidomics from Sample Preparation to Data Analysis: A Primer. Anal. Bioanal. Chem. 2020, 412, 2191–2209. [Google Scholar] [CrossRef]
- Li, Q.; Liang, X.; Zhao, L.; Zhang, Z.; Xue, X.; Wang, K.; Wu, L. UPLC–Q–exactive orbitrap/MS–based Lipidomics Approach to Characterize Lipid Extracts from Bee Pollen and Their in vitro Anti–inflammatory Properties. J. Agric. Food Chem. 2017, 65, 6848–6860. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Chen, H.; Tang, X.; Yang, Q.; Zhang, H.; Chen, Y.Q.; Chen, W. Ultra Performance Liquid Chromatography–Q exactive orbitrap/mass Spectrometry–based Lipidomics Reveals the Influence of Nitrogen Sources on Lipid Biosynthesis of Mortierella alpine. J. Agric. Food Chem. 2019, 67, 10984–10993. [Google Scholar] [CrossRef] [PubMed]
- Nie, Q.; Xing, M.; Chen, H.; Hu, J.; Nie, S. Metabolomics and Lipidomics Profiling Reveals Hypocholesterolemic and Hypolipidemic Effects of Arabinoxylan on Type 2 Diabetic Rats. J. Agric. Food Chem. 2019, 67, 10614–10623. [Google Scholar] [CrossRef]
- Ai, R.; Xu, J.; Ji, G.; Cui, B. Exploring the Phosphatidylcholine in Inflammatory Bowel Disease: Potential Mechanisms and Therapeutic Interventions. Curr. Pharm. Des. 2022, 28, 3486–3491. [Google Scholar]
- Xiao, Y.; Wang, H.; Han, L.; Huang, Z.; Lyu, G.; Li, S. Predictive Value of Anthropometric and Biochemical Indices in Non–alcoholic Fatty Pancreas Disease: A Cross–sectional Study. BMJ Open 2024, 14, e081131. [Google Scholar] [CrossRef]
- Paul, D.; Morrissey, J.H. Stoichiometric Analysis Reveals a Unique Phosphatidylserine Binding Site in Coagulation Factor X. J. Thromb. Haemost. 2022, 20, 600–604. [Google Scholar] [CrossRef]
- Elvas, F.; Stroobants, S.; Wyffels, L. Phosphatidylethanolamine Targeting for Cell Death Imaging in Early Treatment Response Evaluation and Disease Diagnosis. Apoptosis 2017, 22, 971–987. [Google Scholar] [CrossRef]
- Miao, L.; Yang, Y.; Yan, M.; Li, Y.; Zhao, J.; Guo, J.; Zheng, D. Enhanced Immune Response to Rabies Viruses by the Use of a lipid Adjuvant in Vaccines. Viral Immunol. 2017, 30, 727–733. [Google Scholar] [CrossRef]
- Li, C.; Al-Dalali, S.; Zhou, H.; Wang, Z.; Xu, B. Influence of mixture of spices on phospholipid molecules during water-boiled salted duck processing based on shotgun lipidomics. Food Res. Int. 2021, 149, 110651. [Google Scholar] [CrossRef] [PubMed]
- Mariappan, R.; Sukumaran, V.; Ayyavoo, M. Antibacterial Activity of Marine Bivalves Meretrix casta from South East Coast of India. Adv. Bioresearch 2010, 1, 92–96. [Google Scholar]
- Chatterjee, A.; Ansari, Z.A.; Ingole, B.S.; Bichurina, M.A.; Sovetova, M.; Baikov, Y.A. Indian Marine Bivalves: Potential Source of Antiviral Drugs. Curr. Sci. 2010, 2, 1279–1282. [Google Scholar]
- Nazeer, R.A.; Divya Prabha, K.R.; Sampath Kumar, N.S.; Jai Ganesh, R. Isolation of Antioxidant Peptides from Clam, Meretrix casta (Chemnitz). J. Food Sci. Technol. 2013, 50, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Wahidullah, S.; Devi, P.; D’Souza, L. Chemical Composition, Nutritive Value and Health Benefits of Edible Clam Meretrix casta (Chemnitz) from West Coast of India. J. Food Sci. Technol. 2021, 58, 1165–1176. [Google Scholar] [CrossRef]
- Wu, Z.; Zhou, D.; Zhao, M.; Xie, H.; Liu, Z.; Liu, Y.; Liu, Y.; Shahidi, F. Lipid profile and glycerophospholipid molecular species in two species of edible razor clams Sinonovacula constricta and Solen gouldi. Lipids 2019, 54, 347–356. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, D.; Zhou, X.; Yin, F.; Zhao, Q.; Xie, H.; Li, D.; Zhu, B.; Wang, T.; Shahidi, F. Effect of Various Hot-Air Drying Processes on Clam Ruditapes philippinarum Lipids: Composition Changes and Oxidation Development. J. Food Sci. 2018, 83, 2976–2982. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Chang, R.; Liu, C.; Li, J.; Liu, J.; Li, N.; Zhang, Y.; Li, X.; Li, P.; Liu, K. Effect of Freeze Drying and Hot Air Drying on the Composition and Bioactivities of Lipids from Razor Clam Sinonovacula constricta. Foods 2025, 14, 915. https://doi.org/10.3390/foods14060915
Wang D, Chang R, Liu C, Li J, Liu J, Li N, Zhang Y, Li X, Li P, Liu K. Effect of Freeze Drying and Hot Air Drying on the Composition and Bioactivities of Lipids from Razor Clam Sinonovacula constricta. Foods. 2025; 14(6):915. https://doi.org/10.3390/foods14060915
Chicago/Turabian StyleWang, Dexu, Runjia Chang, Changyu Liu, Jiaxun Li, Jibin Liu, Ning Li, Yun Zhang, Xiaobin Li, Peihai Li, and Kechun Liu. 2025. "Effect of Freeze Drying and Hot Air Drying on the Composition and Bioactivities of Lipids from Razor Clam Sinonovacula constricta" Foods 14, no. 6: 915. https://doi.org/10.3390/foods14060915
APA StyleWang, D., Chang, R., Liu, C., Li, J., Liu, J., Li, N., Zhang, Y., Li, X., Li, P., & Liu, K. (2025). Effect of Freeze Drying and Hot Air Drying on the Composition and Bioactivities of Lipids from Razor Clam Sinonovacula constricta. Foods, 14(6), 915. https://doi.org/10.3390/foods14060915