

Ecuadorian Cacao Mucilage as a Novel Culture Medium Ingredient: Unveiling Its Potential for Microbial Growth and Biotechnological Applications

 , ,
, ,
Abstract
1. Introduction
2. Methodology
2.1. Experimental Procedure
2.1.1. Obtaining and Characterizing of Cacao Mucilage Powder (CMP)
2.1.2. Culture Medium
Culture Medium Formulation
Culture Medium Preparation
2.1.3. Microbial Cultivation
Preparation of Inoculum
Inoculation and Incubation
2.2. Statistical Procedure
2.2.1. Experimental Design
2.2.2. Model Response Variables
2.2.3. Optimization Procedure
Numerical and Graphical Optimization
Confirmation Analysis
2.2.4. Effect Size Assessment
2.3. Analytical Determinations
2.3.1. Determination of the Total Microbial Population
2.3.2. Determination of Colony-Forming Units (CFU)
2.3.3. Recovery Rates
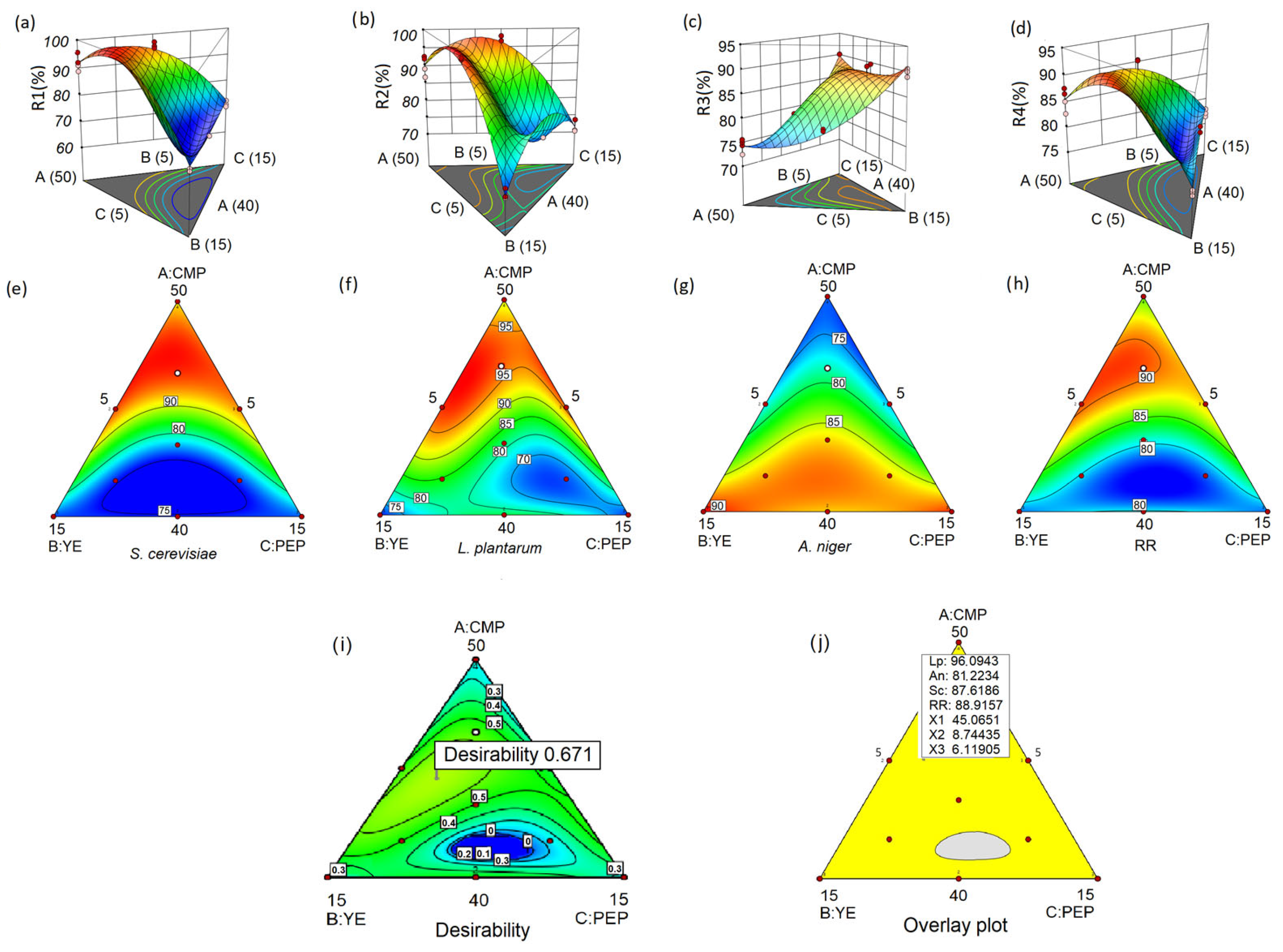
3. Results
3.1. Compositional Profile of Cacao Mucilage
3.2. Analysis of the Obtained Models
3.3. Microbial Species’ Recovery Rate
3.4. Optimization Results
4. Discussion
4.1. Cacao Mucilage Acts as a Catalyst for Microbial Growth
4.2. S. cerevisiae and L. plantarum Rely on Higher Concentrations of Pure Mucilage for Optimal Growth
4.3. High Levels of PEP and YE Are Essential for Optimal Growth of A. niger
4.4. A. niger Shows Limited Growth at High Concentrations of Mucilage
4.5. The Effect Size Indicates That the YPM Medium Is Comparable to Its Traditional Counterparts
4.6. Future Contributions of the YPM Medium and Mucilage
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bastidas, J.V.; Badillo Melo, W.A.; Briones-Bitar, J. Sustainability of the Cocoa Industry: Cocoa Waste Mucilage Use to Produce Fermented Beverages. Case Study in Los Ríos Province. Int. J. Sustain. Dev. Plan. 2022, 17, 1147–1152. [Google Scholar] [CrossRef]
- Balladares, C.; Garca, J.; ChezGuaranda, I.; Prez, S.; Gonzlez, J.; Sosa, D.; Viteri, R.; Barragn, A.; QuijanoAviles, M.; Manzano, P. Physicochemical Characterization of Theobroma Cacao L. Mucilage, in Ecuadorian Coast. Emir. J. Food Agric. 2016, 28, 741. [Google Scholar] [CrossRef]
- Chicaiza Intriago, J.G.; Zambrano Briones, G.E.; Delgado Villafuerte, C.R.; Ávila Martínez, M.F.; Pincay Cantos, M.F. Linear Correlation Analysis of Production Parameters of Biofuel from Cacao (Theobroma cacao L.) Mucilage. J. Ecol. Eng. 2024, 25, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Llerena, W.; Samaniego, I.; Vallejo, C.; Arreaga, A.; Zhunio, B.; Coronel, Z.; Quiroz, J.; Angós, I.; Carrillo, W. Profile of Bioactive Components of Cocoa (Theobroma cacao L.) By-Products from Ecuador and Evaluation of Their Antioxidant Activity. Foods 2023, 12, 2583. [Google Scholar] [CrossRef]
- Martínez, R.; Torres, P.; Meneses, M.A.; Figueroa, J.G.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical, Technological and in Vitro Antioxidant Properties of Cocoa (Theobroma cacao L.) Co-Products. Food Res. Int. 2012, 49, 39–45. [Google Scholar] [CrossRef]
- Vargas-Arana, G.; Merino-Zegarra, C.; Tang, M.; Pertino, M.W.; Simirgiotis, M.J. UHPLC–MS Characterization, and Antioxidant and Nutritional Analysis of Cocoa Waste Flours from the Peruvian Amazon. Antioxidants 2022, 11, 595. [Google Scholar] [CrossRef]
- Morante-Carriel, L.; Abasolo, F.; Bastidas-Caldes, C.; Paz, E.A.; Huaquipán, R.; Díaz, R.; Valdes, M.; Cancino, D.; Sepúlveda, N.; Quiñones, J. Isolation and Characterization of Lactic Acid Bacteria from Cocoa Mucilage and Meat: Exploring Their Potential as Biopreservatives for Beef. Microbiol. Res. 2023, 14, 1150–1167. [Google Scholar] [CrossRef]
- Ghisolfi, R.; Bandini, F.; Vaccari, F.; Bellotti, G.; Bortolini, C.; Patrone, V.; Puglisi, E.; Morelli, L. Bacterial and Fungal Communities Are Specifically Modulated by the Cocoa Bean Fermentation Method. Foods 2023, 12, 2024. [Google Scholar] [CrossRef]
- Parapouli, M.; Vasileiadi, A.; Afendra, A.-S.; Hatziloukas, E. Saccharomyces cerevisiae and Its Industrial Applications. AIMS Microbiol. 2020, 6, 1–32. [Google Scholar] [CrossRef]
- Narendra, K.S. Lactobacillus Plantarum: A Potential Health Booster—A Comprehensive Review. Int. J. Sci. Res. Eng. Manag. 2023, 7. [Google Scholar] [CrossRef]
- Lee, Y.; Jaikwang, N.; Kim, S.K.; Jeong, J.; Sukhoom, A.; Kim, J.-H.; Kim, W. Characterization of a Potential Probiotic Lactiplantibacillus plantarum LRCC5310 by Comparative Genomic Analysis and Its Vitamin B6 Production Ability. J. Microbiol. Biotechnol. 2023, 33, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, B.; Liu, G.; Shi, H.; Wang, J. Effect of Bacillus subtilis and Lactobacillus plantarum on Solid-state Fermentation of Soybean Meal. J. Sci. Food Agric. 2023, 103, 6070–6079. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.C. Citric Acid from Aspergillus niger: A Comprehensive Overview. Crit. Rev. Microbiol. 2020, 46, 727–749. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Xu, X.; Lv, R.; Huang, W.; ul Haq, H.; Gao, Y.; Ren, H.; Lan, C.; Tian, B. Differential Proteomics Reveals Main Determinants for the Improved Pectinase Activity in UV-Mutagenized Aspergillus niger Strain. Biotechnol. Lett. 2021, 43, 909–918. [Google Scholar] [CrossRef]
- Conesa, A.; van den Hondel, C.A.; Punt, P.J. Studies on the Production of Fungal Peroxidases in Aspergillus niger. Appl. Environ. Microbiol. 2000, 66, 3016–3023. [Google Scholar] [CrossRef]
- Vergara-Mendoza, M.; Martínez, G.R.; Blanco-Tirado, C.; Combariza, M.Y. Mass Balance and Compositional Analysis of Biomass Outputs from Cacao Fruits. Molecules 2022, 27, 3717. [Google Scholar] [CrossRef]
- Cervantes-Martínez, C.V.; Medina-Torres, L.; González-Laredo, R.F.; Calderas, F.; Sánchez-Olivares, G.; Herrera-Valencia, E.E.; Gallegos Infante, J.A.; Rocha-Guzman, N.E.; Rodríguez-Ramírez, J. Study of Spray Drying of the Aloe Vera Mucilage (Aloe vera barbadensis Miller) as a Function of Its Rheological Properties. LWT-Food Sci. Technol. 2014, 55, 426–435. [Google Scholar] [CrossRef]
- Weenk, G.H. Microbiological Assessment of Culture Media: Comparison and Statistical Evaluation of Methods. Int. J. Food Microbiol. 1992, 17, 159–181. [Google Scholar] [CrossRef]
- AOAC. Protein (Crude) in Animal Feed, Forage (Plant Tissue), Grain, and Oil Seeds. Block Digestion Method Using Copper Catalyst and Steam Distillation into Boric Acid; AOAC: Rockville, MD, USA, 2000. [Google Scholar]
- AOAC. Fat (Crude) or Ether Extract in Animal Feed; AOAC: Rockville, MD, USA, 2000. [Google Scholar]
- AOAC. Crude Fiber in Cacao Products; AOAC: Rockville, MD, USA, 2000. [Google Scholar]
- AOAC. Ash (Acid-Insoluble) of Cacao Products; AOAC: Rockville, MD, USA, 2000. [Google Scholar]
- AOAC. Loss on Drying (Moisture) in Plants; AOAC: Rockville, MD, USA, 2000. [Google Scholar]
- ISO 6887-1; Microbiology of the Food Chain—Preparation of Test Samples, Initial Suspension and Decimal Dilutions for Microbiological Examination. International Organization for Standardization: Geneva, Switzerland, 2017.
- Bonnet, M.; Lagier, J.C.; Raoult, D.; Khelaifia, S. Bacterial Culture through Selective and Non-Selective Conditions: The Evolution of Culture Media in Clinical Microbiology. New Microbes New Infect. 2020, 34, 100622. [Google Scholar] [CrossRef]
- Abdel Massih, M.; Planchon, V.; Pitchugina, E.; Mahillon, J. Enumeration of Lactic Acid Bacteria: Lacuna and Improvement Areas Highlighted by Proficiency Testing. Accredit. Qual. Assur. 2019, 24, 381–385. [Google Scholar] [CrossRef]
- Galvan, D.; Effting, L.; Cremasco, H.; Conte-Junior, C.A. Recent Applications of Mixture Designs in Beverages, Foods, and Pharmaceutical Health: A Systematic Review and Meta-Analysis. Foods 2021, 10, 1941. [Google Scholar] [CrossRef] [PubMed]
- Gunst, R.F. Response Surface Methodology: Process and Product Optimization Using Designed Experiments. Technometrics 1996, 38, 284–286. [Google Scholar] [CrossRef]
- Maher, J.M.; Markey, J.C.; Ebert-May, D. The Other Half of the Story: Effect Size Analysis in Quantitative Research. CBE—Life Sci. Educ. 2013, 12, 345–351. [Google Scholar] [CrossRef]
- Wang, R.; Lorantfy, B.; Fusco, S.; Olsson, L.; Franzén, C.J. Analysis of Methods for Quantifying Yeast Cell Concentration in Complex Lignocellulosic Fermentation Processes. Sci. Rep. 2021, 11, 11293. [Google Scholar] [CrossRef]
- Downey, A.S.; Da Silva, S.M.; Olson, N.D.; Filliben, J.J.; Morrow, J.B. Impact of Processing Method on Recovery of Bacteria from Wipes Used in Biological Surface Sampling. Appl. Environ. Microbiol. 2012, 78, 5872–5881. [Google Scholar] [CrossRef]
- Vizcaino-Almeida, C.R.; Guajardo-Flores, D.; Caroca-Cáceres, R.; Serna-Saldívar, S.O.; Briones-García, M.; Lazo-Vélez, M.A. Non-conventional Fermentation at Laboratory Scale of Cocoa Beans: Using Probiotic Microorganisms and Substitution of Mucilage by Fruit Pulps. Int. J. Food Sci. Technol. 2022, 57, 4307–4315. [Google Scholar] [CrossRef]
- Viesser, J.A.; de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Rogez, H.; Góes-Neto, A.; Azevedo, V.; Brenig, B.; Aburjaile, F.; Soccol, C.R. Co-Culturing Fructophilic Lactic Acid Bacteria and Yeast Enhanced Sugar Metabolism and Aroma Formation during Cocoa Beans Fermentation. Int. J. Food Microbiol. 2021, 339, 109015. [Google Scholar] [CrossRef]
- Da Veiga Moreira, I.M.; Miguel, M.G.d.C.P.; Duarte, W.F.; Dias, D.R.; Schwan, R.F. Microbial Succession and the Dynamics of Metabolites and Sugars during the Fermentation of Three Different Cocoa (Theobroma cacao L.) Hybrids. Food Res. Int. 2013, 54, 9–17. [Google Scholar] [CrossRef]
- Garcia Gonzalez, E.; Ochoa Muñoz, A.F.; Montalvo Rodríguez, C.; Ordoñez Narvaéz, G.A.; Londoño Hernández, L. Sucesión Microbiana Durante La Fermentación Espontánea de Cacao En Unidades Productivas. Cienc. Desarro. 2021, 12, 21–30. [Google Scholar] [CrossRef]
- Lima, C.O.D.C.; Vaz, A.B.; De Castro, G.M.; Lobo, F.; Solar, R.; Rodrigues, C.; Martins Pinto, L.R.; Vandenberghe, L.; Pereira, G.; Miúra da Costa, A.; et al. Integrating Microbial Metagenomics and Physicochemical Parameters and a New Perspective on Starter Culture for Fine Cocoa Fermentation. Food Microbiol. 2021, 93, 103608. [Google Scholar] [CrossRef]
- Ijadpanahsaravi, M.; Punt, M.; Wösten, H.A.B.; Teertstra, W.R. Minimal Nutrient Requirements for Induction of Germination of Aspergillus Niger Conidia. Fungal Biol. 2021, 125, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Bircher, L.; Sourabié, A.M.; Paurevic, M.; Hochuli, J.; Geirnaert, A.; Navas, C.; Drogue, B.; Lacroix, C. Faecalibacterium duncaniae A2-165 Growth Is Strongly Promoted by Yeast Extract and Vitamin B5 in CGMP Medium. Microb. Biotechnol. 2024, 17, e14374. [Google Scholar] [CrossRef] [PubMed]
- Kligler, I.J. Yeast autolysate as a culture medium for bacteria. J. Bacteriol. 1919, 4, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Nancib, N.; Branlant, C.; Boudrant, J. Metabolic Roles of Peptone and Yeast Extract for the Culture of a Recombinant Strain Of Escherichia coli. J. Ind. Microbiol. 1991, 8, 165–169. [Google Scholar] [CrossRef]
- Germec, M.; Turhan, I. Enhanced Production of Aspergillus Niger Inulinase from Sugar Beet Molasses and Its Kinetic Modeling. Biotechnol. Lett. 2020, 42, 1939–1955. [Google Scholar] [CrossRef]
- Waring, B.G.; Averill, C.; Hawkes, C.V. Differences in Fungal and Bacterial Physiology Alter Soil Carbon and Nitrogen Cycling: Insights from Meta-analysis and Theoretical Models. Ecol. Lett. 2013, 16, 887–894. [Google Scholar] [CrossRef]
- Araújo, C.A.; Ferreira, P.C.; Pupin, B.; Dias, L.P.; Avalos, J.; Edwards, J.; Hallsworth, J.E.; Rangel, D.E.N. Osmotolerance as a Determinant of Microbial Ecology: A Study of Phylogenetically Diverse Fungi. Fungal Biol. 2020, 124, 273–288. [Google Scholar] [CrossRef]
- Stratford, M.; Steels, H.; Novodvorska, M.; Archer, D.B.; Avery, S.V. Extreme Osmotolerance and Halotolerance in Food-Relevant Yeasts and the Role of Glycerol-Dependent Cell Individuality. Front. Microbiol. 2019, 9, 3238. [Google Scholar] [CrossRef]
- Bubnová, M.; Zemančíková, J.; Sychrová, H. Osmotolerant Yeast Species Differ in Basic Physiological Parameters and in Tolerance of Non-osmotic Stresses. Yeast 2014, 31, 309–321. [Google Scholar] [CrossRef]
- Liu, G.; Chen, Y.; Færgeman, N.J.; Nielsen, J. Elimination of the Last Reactions in Ergosterol Biosynthesis Alters the Resistance of Saccharomyces Cerevisiae to Multiple Stresses. FEMS Yeast Res. 2017, 17, fox063. [Google Scholar] [CrossRef]
- Meyer, V.; Damveld, R.A.; Arentshorst, M.; Stahl, U.; van den Hondel, C.A.; Ram, A.F. Survival in the Presence of Antifungals. J. Biol. Chem. 2007, 282, 32935–32948. [Google Scholar] [CrossRef] [PubMed]
- Perczyk, P.; Wójcik, A.; Wydro, P.; Broniatowski, M. The Role of Phospholipid Composition and Ergosterol Presence in the Adaptation of Fungal Membranes to Harsh Environmental Conditions–Membrane Modeling Study. Biochim. Biophys. Acta 2020, 1862, 183136. [Google Scholar] [CrossRef]
- Jordá, T.; Puig, S. Regulation of Ergosterol Biosynthesis in Saccharomyces Cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Zong, Y.; Zhang, X.; Gong, D.; Wang, B.; Prusky, D.; Sionov, E.; Xue, H.; Bi, Y. Erg4 Is Involved in Ergosterol Biosynthesis, Conidiation and Stress Response in Penicillium expansum. J. Fungi 2023, 9, 568. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.S.; Xiang, Y.; Liang, Z.-X.; Marcellin, E.; Pham, H.T. Cyclic-Di-AMP Signalling in Lactic Acid Bacteria. FEMS Microbiol. Rev. 2023, 47, fuad025. [Google Scholar] [CrossRef] [PubMed]
- Le Marrec, C. Responses of Lactic Acid Bacteria to Osmotic Stress. In Stress Responses of Lactic Acid Bacteria; Springer: Boston, MA, USA, 2011; pp. 67–90. [Google Scholar]
- Duveau, F.; Cordier, C.; Chiron, L.; LeBec, M.; Pouzet, S.; Séguin, J.; Llamosi, A.; Sorre, B.; Di Meglio, J.-M.; Hersen, P. Yeast Cell Responses and Survival during Periodic Osmotic Stress Are Controlled by Glucose Availability. Elife 2023, 12, RP88750. [Google Scholar] [CrossRef]
- Galello, F.; Bermúdez-Moretti, M.; Ortolá Martínez, M.C.; Rossi, S.; Portela, P. The CAMP-PKA Signalling Crosstalks with CWI and HOG-MAPK Pathways in Yeast Cell Response to Osmotic and Thermal Stress. Microb. Cell 2024, 11, 90–105. [Google Scholar] [CrossRef]
- Duran, R.; Cary, J.W.; Calvo, A.M. Role of the Osmotic Stress Regulatory Pathway in Morphogenesis and Secondary Metabolism in Filamentous Fungi. Toxins 2010, 2, 367–381. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, M.; Wang, H.; Yu, G.; Guo, A.; Ren, W.; Li, B.; Liu, N. The Mitogen-Activated Protein Kinase Hog1 Regulates Fungal Development, Pathogenicity, and Stress Response in Botryosphaeria dothidea. Phytopathology 2024, 114, 725–731. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Pei, J.; Yan, H.; Wang, Y. The AwHog1 Transcription Factor Influences the Osmotic Stress Response, Mycelium Growth, OTA Production, and Pathogenicity in Aspergillus westerdijkiae Fc-1. Toxins 2023, 15, 432. [Google Scholar] [CrossRef]

{kind=link}
| Labels | Central Tendency Measures | ||||||
|---|---|---|---|---|---|---|---|
| Factors | Variables | Minimum | Maximum | Minor Code | High Code | Mean | Sd |
| A | CMP | 40 | 50 | +0 ↔ 40 | +0.5 ↔ 50 | 43.63 | 3.83 |
| B | YE | 5 | 15 | +0 ↔ 5 | +0.5 ↔ 15 | 8.02 | 3.67 |
| C | PEP | 5 | 15 | +0 ↔ 5 | +0.5 ↔ 15 | 8.25 | 3.63 |
| Random Runs | Blocks | Runs | CMP | YE | PEP |
|---|---|---|---|---|---|
| 3 | Block 1 | 1 | 50 | 5 | 5 |
| 21 | Block 1 | 2 | 50 | 5 | 5 |
| 11 | Block 1 | 3 | 43.33 | 8.33 | 8.33 |
| 19 | Block 1 | 4 | 40 | 5 | 15 |
| 2 | Block 1 | 5 | 40 | 5 | 15 |
| 12 | Block 1 | 6 | 40 | 15 | 5 |
| 7 | Block 1 | 7 | 41.66 | 6.66 | 11.66 |
| 10 | Block 1 | 8 | 46.66 | 6.66 | 6.66 |
| 9 | Block 1 | 9 | 40 | 5 | 15 |
| 18 | Block 1 | 10 | 40 | 15 | 5 |
| 13 | Block 1 | 11 | 40 | 15 | 5 |
| 8 | Block 2 | 12 | 50 | 5 | 5 |
| 17 | Block 2 | 13 | 40 | 10 | 10 |
| 15 | Block 2 | 14 | 45 | 5 | 10 |
| 1 | Block 2 | 15 | 45 | 10 | 5 |
| 4 | Block 2 | 16 | 40 | 10 | 10 |
| 16 | Block 2 | 17 | 45 | 5 | 10 |
| 5 | Block 2 | 18 | 45 | 5 | 10 |
| 20 | Block 2 | 19 | 41.66 | 11.66 | 6.66 |
| 14 | Block 2 | 20 | 45 | 10 | 5 |
| 6 | Block 2 | 21 | 50 | 5 | 5 |
| Runs | CEP | YE | PEP | R1(%) | R2(%) | R3(%) | R4(%) |
|---|---|---|---|---|---|---|---|
| 1 | 50 | 5 | 5 | 88.74 | 86.72 | 72.42 | 82.627 |
| 2 | 50 | 5 | 5 | 91.91 | 91.9 | 75.53 | 86.447 |
| 3 | 43.33 | 8.33 | 8.33 | 73.35 | 83.78 | 84.64 | 80.590 |
| 4 | 40 | 5 | 15 | 71.03 | 70.55 | 87.77 | 76.450 |
| 5 | 40 | 5 | 15 | 72.52 | 70.31 | 88.58 | 77.137 |
| 6 | 40 | 15 | 5 | 71.25 | 70.61 | 90.6 | 77.487 |
| 7 | 41.66 | 6.66 | 11.66 | 70.9 | 72.71 | 86.43 | 76.680 |
| 8 | 46.66 | 6.66 | 6.66 | 96.22 | 94.77 | 77 | 89.330 |
| 9 | 40 | 5 | 15 | 70.16 | 73.73 | 90.15 | 78.013 |
| 10 | 40 | 15 | 5 | 70.28 | 70.96 | 88.95 | 76.730 |
| 11 | 40 | 15 | 5 | 70.25 | 73.27 | 89.99 | 77.837 |
| 12 | 50 | 5 | 5 | 95.56 | 92.55 | 74.33 | 87.480 |
| 13 | 40 | 10 | 10 | 70.37 | 89.56 | 85.56 | 81.830 |
| 14 | 45 | 5 | 10 | 97.07 | 98.06 | 76.03 | 90.387 |
| 15 | 45 | 10 | 5 | 98.41 | 98.7 | 78.13 | 91.747 |
| 16 | 40 | 10 | 10 | 74.93 | 77.44 | 89.67 | 80.680 |
| 17 | 45 | 5 | 10 | 95.52 | 96.68 | 78.53 | 90.243 |
| 18 | 45 | 5 | 10 | 94.81 | 93.95 | 75.28 | 88.013 |
| 19 | 41.66 | 11.66 | 6.66 | 72.87 | 77.56 | 90.36 | 80.263 |
| 20 | 45 | 10 | 5 | 97.12 | 99.02 | 77.68 | 91.273 |
| 21 | 50 | 5 | 5 | 90.89 | 89.09 | 74.98 | 84.987 |
| Models | Sig. Term p < 0.05 | Lack of Fit (p) | F-Value | p Value | R2 Adjusted | R2 Predicted | CV (%) | Adeq Precision |
|---|---|---|---|---|---|---|---|---|
| Model R1 | AB, AC | 0.56 | 84.43 | <0.00 | 0.97 | 0.93 | 2.14 | 22.48 |
| Model R2 | AB, AC, BC | 0.19 | 15.57 | <0.00 | 0.82 | 0.65 | 4.32 | 11.59 |
| Model R3 | AB, AC ABC | 0.98 | 52.14 | <0.00 | 0.94 | 0.88 | 1.86 | 17.59 |
| Model R4 | AB, AC ABC | 0.39 | 57.39 | <0.00 | 0.92 | 0.81 | 1.51 | 16.68 |
| Responses | Predicted Mean | Standard Deviation | Standard Error | Number of Runs | Lower Limit LI | Observed Mean | Upper Limit. LS |
|---|---|---|---|---|---|---|---|
| R1 | 87.607 | 1.771 | 1.680 | 7 | 83.920 | 88.160 | 91.316 |
| R2 | 94.280 | 3.700 | 2.357 | 7 | 89.187 | 91.191 | 99.372 |
| R3 | 82.705 | 1.537 | 1.111 | 7 | 80.303 | 81.742 | 85.103 |
| R4 | 87.399 | 1.252 | 1.187 | 7 | 84.788 | 87.897 | 90.018 |
| M.O | R (%) | YPM (UFC/g) | MRS (UFC/g) | MYA (UFC/g) | SDA (UFC/g) | d–Cohen | p |
|---|---|---|---|---|---|---|---|
| S. cerevisiae | 88.160 | 7.68 ± 5.95 a | – | 8.73 ± 0.62 a | – | 0.2 | 0.649 |
| L plantarum | 91.191 | 7.74 ± 6.08 a | 8.84 ± 0.62 a | – | – | 0.2 | 0.642 |
| A. niger | 81.742 | 7.31 ± 1.46 a | – | – | 9.16 ± 0.68 b | 1.6 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzmán-Armenteros, T.M.; Guerra, L.S.; Ruales, J.; Ramos-Guerrero, L. Ecuadorian Cacao Mucilage as a Novel Culture Medium Ingredient: Unveiling Its Potential for Microbial Growth and Biotechnological Applications. Foods 2025, 14, 261. https://doi.org/10.3390/foods14020261
Guzmán-Armenteros TM, Guerra LS, Ruales J, Ramos-Guerrero L. Ecuadorian Cacao Mucilage as a Novel Culture Medium Ingredient: Unveiling Its Potential for Microbial Growth and Biotechnological Applications. Foods. 2025; 14(2):261. https://doi.org/10.3390/foods14020261
Chicago/Turabian StyleGuzmán-Armenteros, Tania María, Luis Santiago Guerra, Jenny Ruales, and Luis Ramos-Guerrero. 2025. "Ecuadorian Cacao Mucilage as a Novel Culture Medium Ingredient: Unveiling Its Potential for Microbial Growth and Biotechnological Applications" Foods 14, no. 2: 261. https://doi.org/10.3390/foods14020261
APA StyleGuzmán-Armenteros, T. M., Guerra, L. S., Ruales, J., & Ramos-Guerrero, L. (2025). Ecuadorian Cacao Mucilage as a Novel Culture Medium Ingredient: Unveiling Its Potential for Microbial Growth and Biotechnological Applications. Foods, 14(2), 261. https://doi.org/10.3390/foods14020261