
Phyllanthus emblica Prevents Adipogenesis by Regulating Histone Acetylation

 , , ,
, , ,  ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Histone Acetyltransferase (HAT) Activity Assay
2.3. Cell Culture and Differentiation
2.4. Cell Viability
2.5. Oil-Red-O (ORO) Staining Assay
2.6. Triglyceride (TG) Content
2.7. Extraction of Histone from 3T3-L1 Cells
2.8. Quantitative Real-Time RT-PCR
2.9. Western Blotting
2.10. Analysis of Main Compounds Using LC-MS/MS
2.11. Statistical Analysis
3. Results
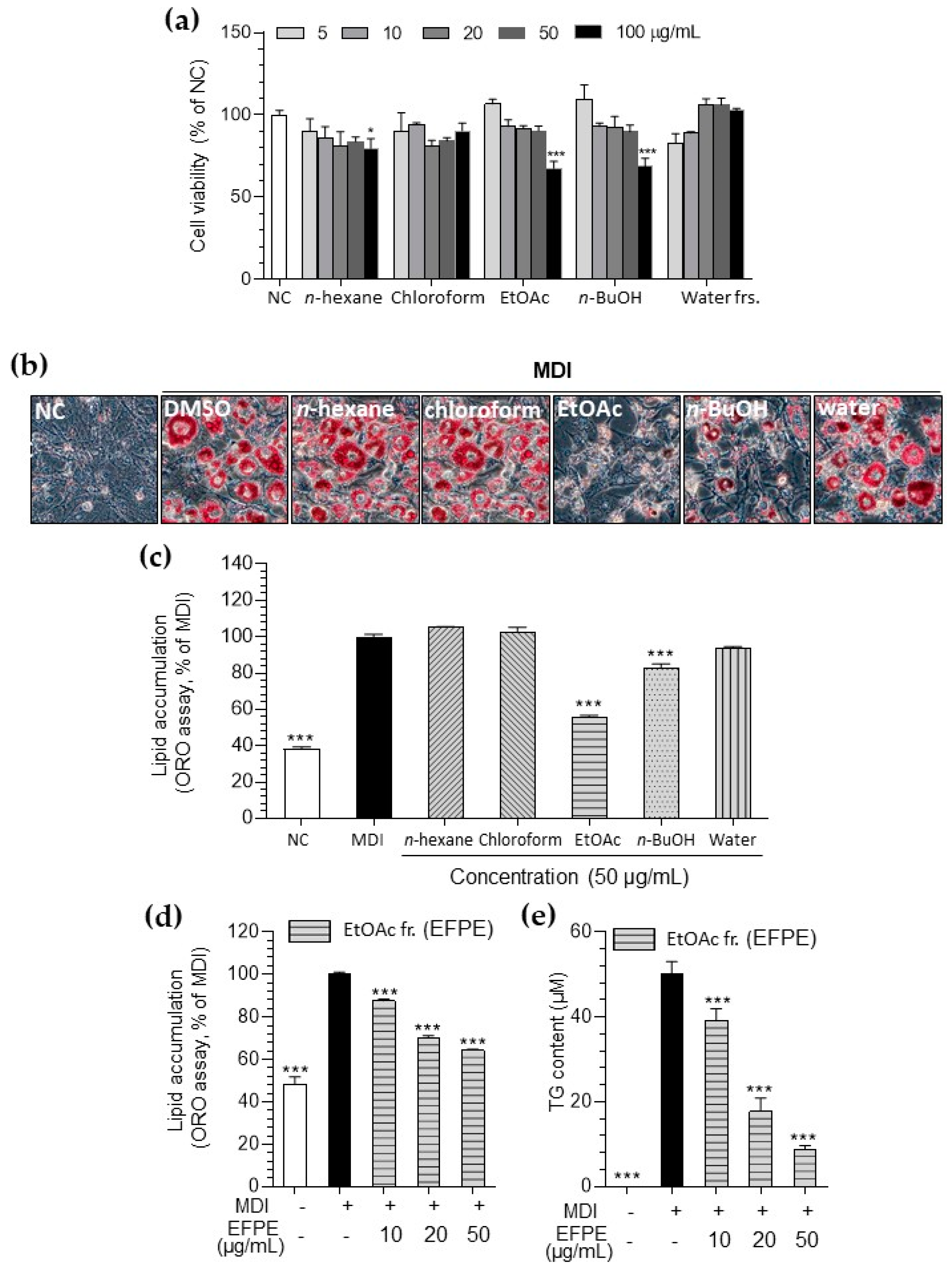
3.1. Inhibitory Effect of P. emblica on Lipid Accumulation in 3T3-L1 Adipocytes
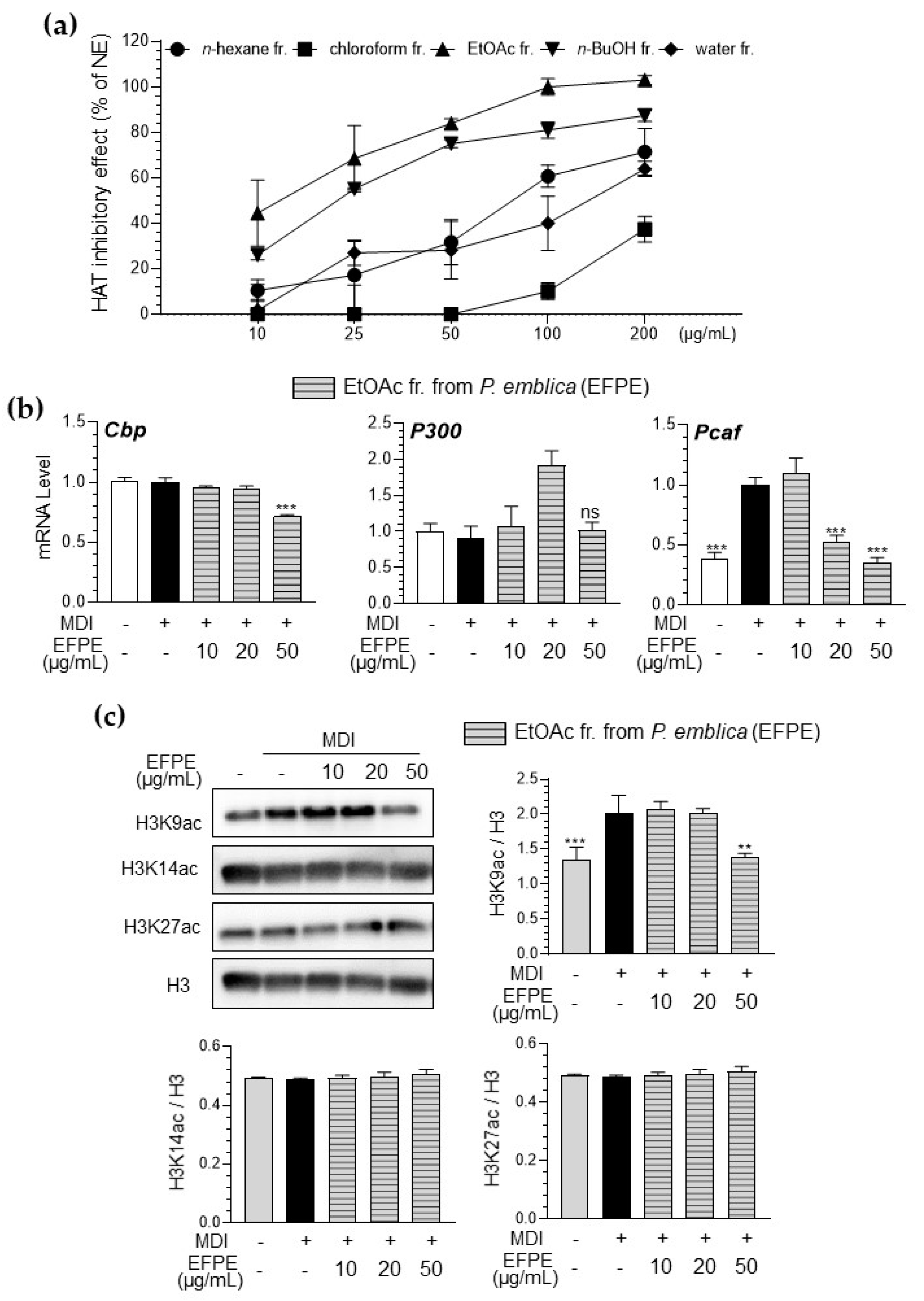
3.2. Inhibitory Effect of Various Fractions of P. emblica Against HAT Activity in a Cell-Free System
3.3. EFPE Regulates Adipogenesis-Related Gene Expression in 3T3-L1 Adipocytes
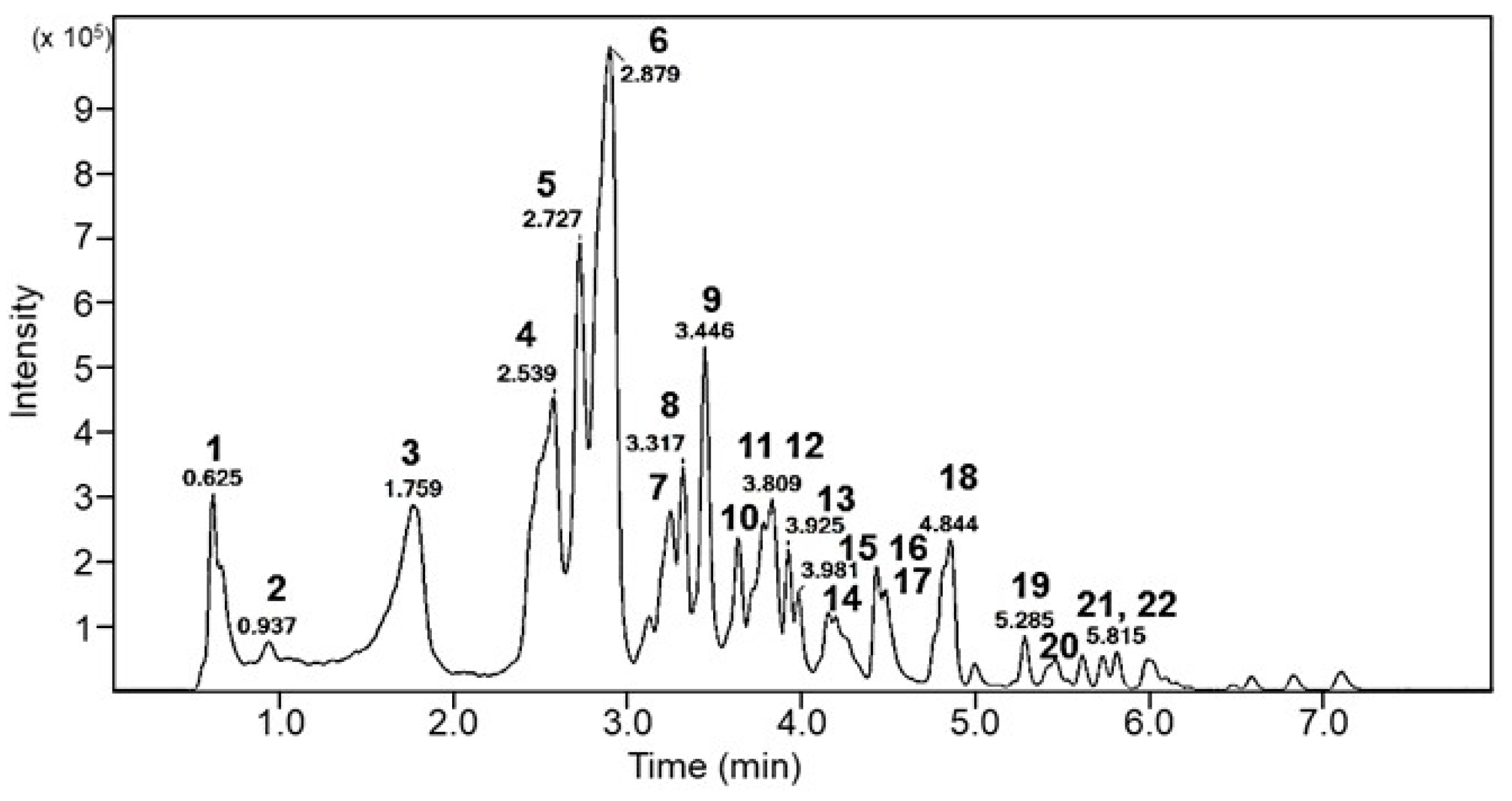
3.4. Analysis of Bioactive Components in EFPE
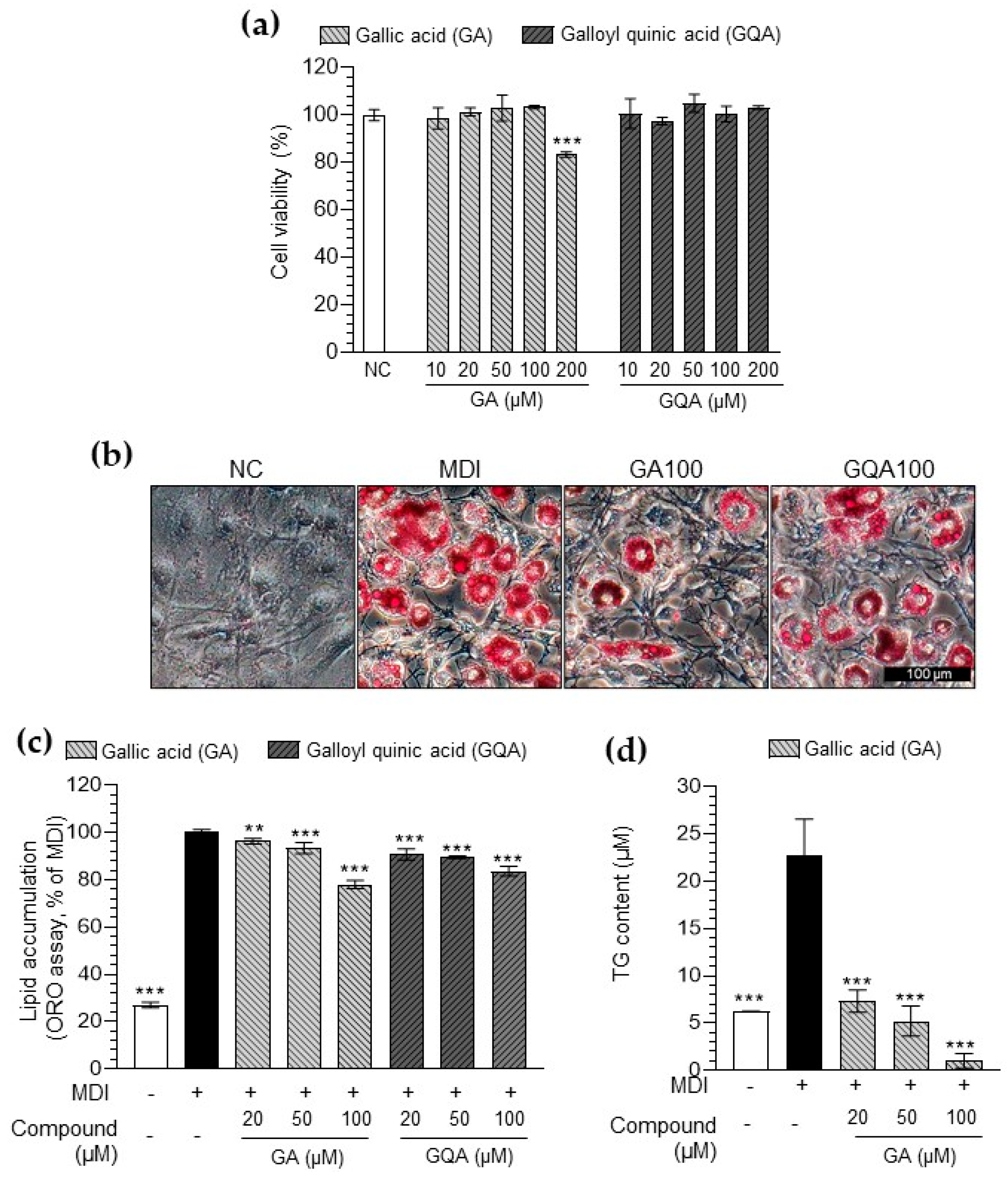
3.5. Inhibitory Effect of GA and GQA on Lipid Accumulation in 3T3-L1 Adipocytes
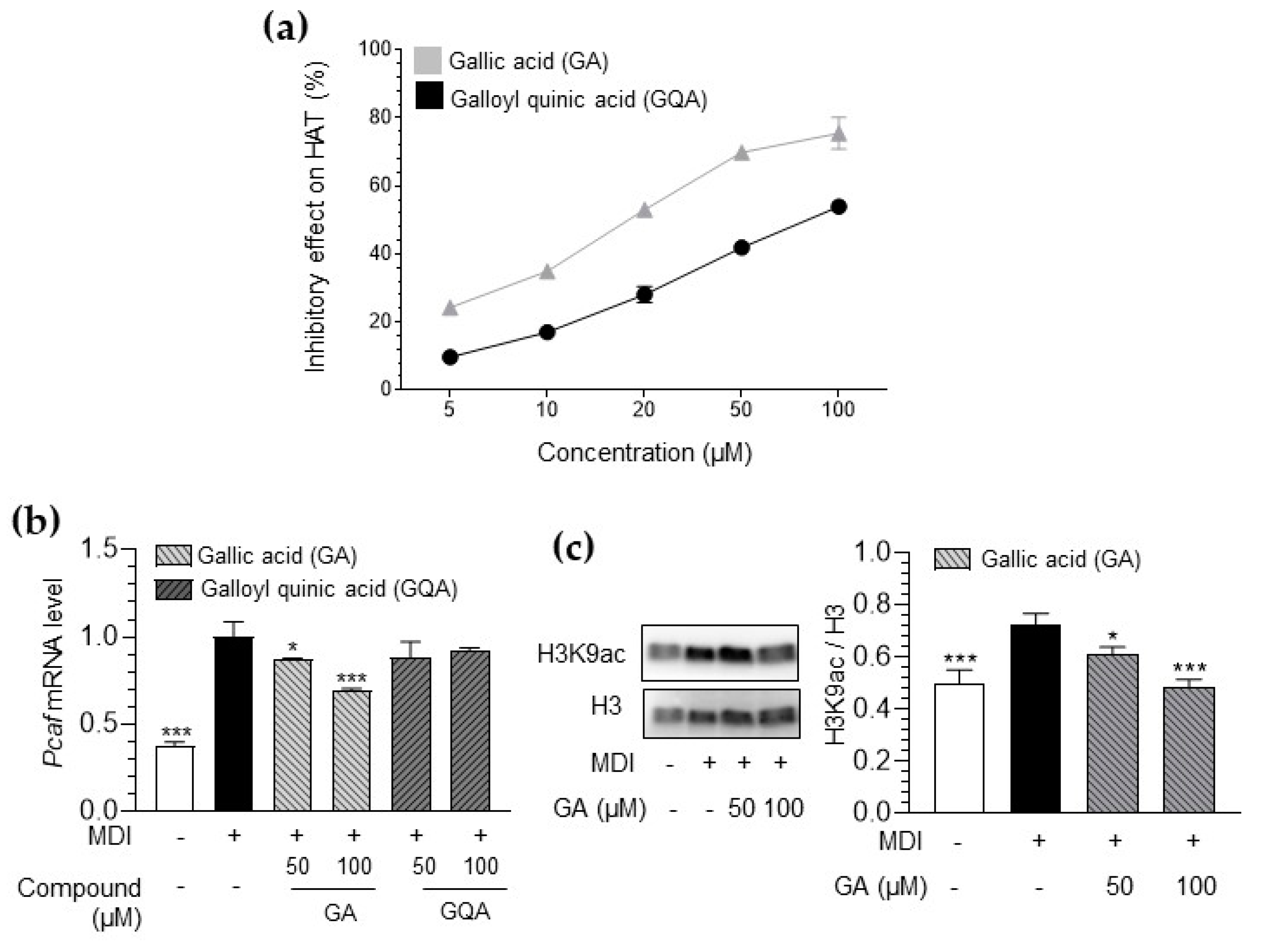
3.6. Inhibitory Effect of GA and GQA on HAT Activity and Histone Acetylation
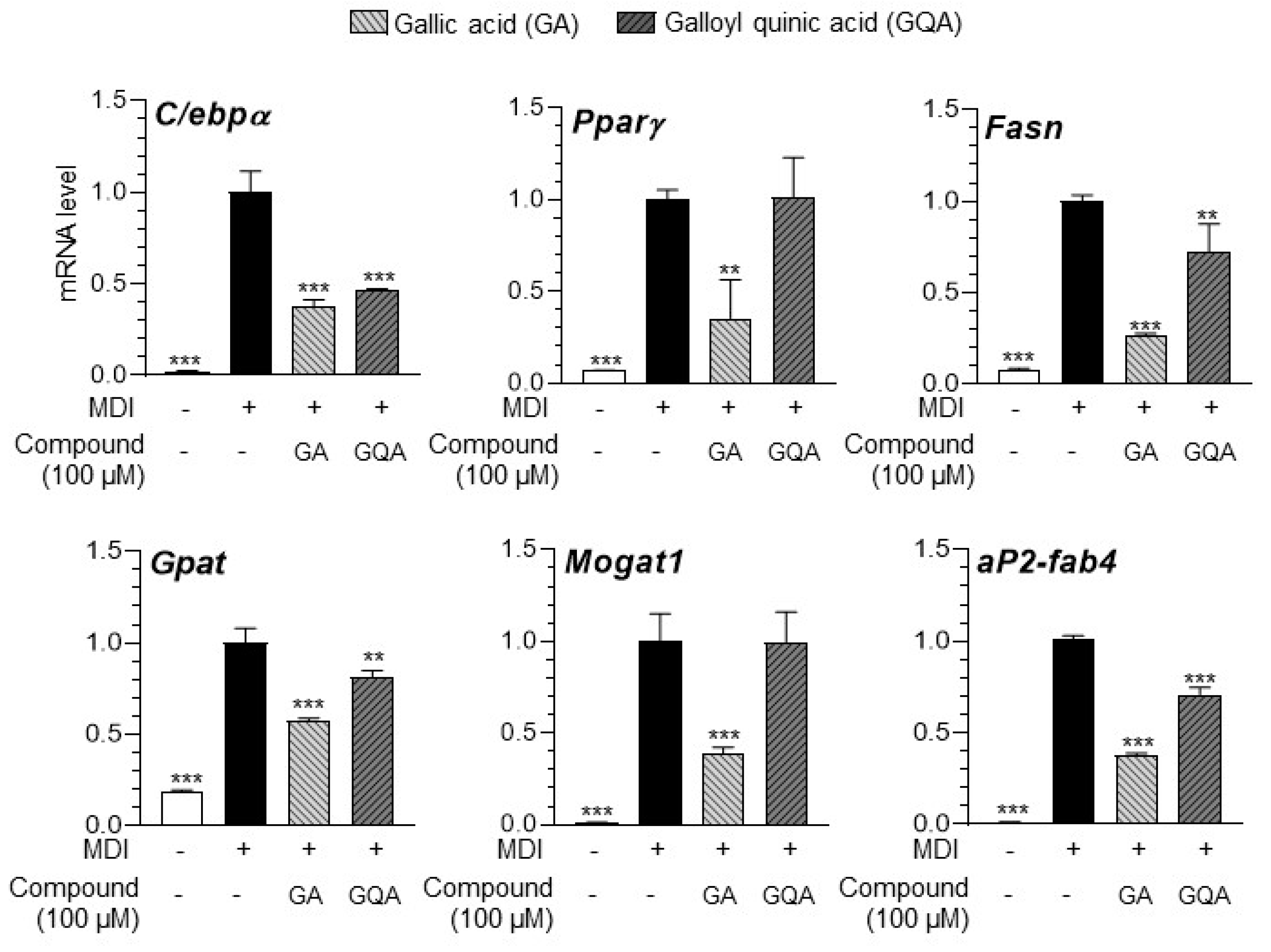
3.7. GA and GQA Regulates Adipogenesis-Related Gene Expression in 3T3-L1 Adipocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paniagua, J.A. Nutrition, insulin resistance and dysfunctional adipose tissue determine the different components of metabolic syndrome. World J. Diabetes 2016, 7, 483–514. [Google Scholar] [CrossRef] [PubMed]
- Balusamy, S.R.; Veerappan, K.; Ranjan, A.; Kim, Y.-J.; Chellappan, D.K.; Dua, K.; Lee, J.; Perumalsamy, H. Phyllanthus emblica fruit extract attenuates lipid metabolism in 3T3-L1 adipocytes via activating apoptosis mediated cell death. Phytomedicine 2020, 66, 153129. [Google Scholar] [CrossRef] [PubMed]
- Fajas, L. Adipogenesis: A cross-talk between cell proliferation and cell differentiation. Ann. Med. 2003, 35, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [CrossRef]
- Jee, W.; Lee, S.-H.; Ko, H.M.; Jung, J.H.; Chung, W.-S.; Jang, H.-J. Anti-obesity effect of polygalin C isolated from Polygala japonica Houtt. via suppression of the adipogenic and lipogenic factors in 3T3-L1 adipocytes. Int. J. Mol. Sci. 2021, 22, 10405. [Google Scholar] [CrossRef]
- Lillycrop, K.A.; Burdge, G.C. Maternal diet as a modifier of offspring epigenetics. J. Dev. Orig. Health Dis. 2015, 6, 88–95. [Google Scholar] [CrossRef]
- Silva, L.B.A.R.; Pinheiro-Castro, N.; Novaes, G.M.; Pascoal, G.D.F.L.; Ong, T.P. Bioactive food compounds, epigenetics and chronic disease prevention: Focus on early-life interventions with polyphenols. Food Res. Int. 2019, 125, 108646. [Google Scholar] [CrossRef]
- Ramazi, S.; Allahverdi, A.; Zahiri, J. Evaluation of post-translational modifications in histone proteins: A review on histone modification defects in developmental and neurological disorders. J. Biosci. 2020, 45, 135. [Google Scholar] [CrossRef]
- Zhou, Y.; Peng, J.; Jiang, S. Role of histone acetyltransferases and histone deacetylases in adipocyte differentiation and adipogenesis. Eur. J. Cell Biol. 2014, 93, 170–177. [Google Scholar] [CrossRef]
- Milagro, F.I.; Mansego, M.L.; De Miguel, C.; Martínez, J.A. Dietary factors, epigenetic modifications and obesity outcomes: Progresses and perspectives. Mol. Asp. Med. 2013, 34, 782–812. [Google Scholar] [CrossRef]
- Jiménez-Chillarón, J.C.; Díaz, R.; Martínez, D.; Pentinat, T.; Ramón-Krauel, M.; Ribó, S.; Plösch, T. The role of nutrition on epigenetic modifications and their implications on health. Biochimie 2012, 94, 2242–2263. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, M.; Wang, J.; Yang, B.; Jiang, Y. Antioxidant activity of methanolic extract of Emblica fruit (Phyllanthus emblica L.) from six regions in China. J. Food Compos. Anal. 2008, 21, 219–228. [Google Scholar] [CrossRef]
- Ahmad, B.; Hafeez, N.; Rauf, A.; Bashir, S.; Linfang, H.; Rehman, M.; Mubarak, M.S.; Uddin, M.S.; Bawazeer, S.; Shariati, M.A.; et al. Phyllanthus emblica: A comprehensive review of its therapeutic benefits. S. Afr. J. Bot. 2021, 138, 278–310. [Google Scholar] [CrossRef]
- Gaire, B.P.; Subedi, L. Phytochemistry, pharmacology and medicinal properties of Phyllanthus emblica Linn. Chin. J. Integr. Med. 2014, 1–8. [Google Scholar] [CrossRef]
- Goran, M.I. Energy metabolism and obesity. Med. Clin. N. Am. 2000, 84, 347–362. [Google Scholar] [CrossRef]
- Li, H.X.; Xiao, L.; Wang, C.; Gao, J.L.; Zhai, Y.G. Review: Epigenetic regulation of adipocyte differentiation and adipogenesis. J. Zhejiang Univ. Sci. B 2010, 11, 784–791. [Google Scholar] [CrossRef]
- Pham, T.X.; Lee, J. Dietary regulation of histone acetylases and deacetylases for the prevention of metabolic diseases. Nutrients 2012, 4, 1868–1886. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Chen, S.Y.; Lin, J.A.; Yen, G.C. Preventive effect of Indian gooseberry (Phyllanthus emblica L.) fruit extract on cognitive decline in high-fat diet (HFD)-fed rats. Mol. Nutr. Food Res. 2023, 67, e2200791. [Google Scholar] [CrossRef]
- Huang, C.-Z.; Tung, Y.-T.; Hsia, S.-M.; Wu, C.-H.; Yen, G.-C. The hepatoprotective effect of Phyllanthus emblica L. fruit on high fat diet-induced non-alcoholic fatty liver disease (NAFLD) in SD rats. Food Funct. 2017, 8, 842–850. [Google Scholar] [CrossRef]
- Kim, S.-J.; Nian, C.; McIntosh, C.H.S. Adipocyte expression of the glucose-dependent insulinotropic polypeptide receptor involves gene regulation by PPARγ and histone acetylation. J. Lipid Res. 2011, 52, 759–770. [Google Scholar] [CrossRef]
- Barilla, S.; Treuter, E.; Venteclef, N. Transcriptional and epigenetic control of adipocyte remodeling during obesity. Obesity (Silver Spring) 2021, 29, 2013–2025. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Schmidt, H.; Lai, B.; Ge, K. Transcriptional and epigenomic regulation of adipogenesis. Mol. Cell. Biol. 2019, 39, e00601-18. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-T.; Choi, H.-K.; Kim, S.H.; Chung, S.; Hur, H.J.; Park, J.H.; Chung, M.-Y. Hypolipidemic activity of Quercus acutissima fruit ethanol extract is mediated by inhibition of acetylation. J. Med. Food 2017, 20, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-W.; Friso, S. Epigenetics: A new bridge between nutrition and health. Adv. Nutr. 2010, 1, 8–16. [Google Scholar] [CrossRef]
- Jin, Q.; Yu, L.R.; Wang, L.; Zhang, Z.; Kasper, L.H.; Lee, J.E.; Wang, C.; Brindle, P.K.; Dent, S.Y.R.; Ge, K. Distinct roles of GCN5/PCAF-mediated H3K9ac and CBP/p300-mediated H3K18/27ac in nuclear receptor transactivation. EMBO J. 2011, 30, 249–262. [Google Scholar] [CrossRef]
- Ong, B.X.; Brunmeir, R.; Zhang, Q.; Peng, X.; Idris, M.; Liu, C.; Xu, F. Regulation of thermogenic adipocyte differentiation and adaptive thermogenesis through histone acetylation. Front. Endocrinol. 2020, 11, 95. [Google Scholar] [CrossRef]
- Abd Eldaim, M.A.; Matsuoka, S.; Okamatsu-Ogura, Y.; Kamikawa, A.; Ahmed, M.M.; Terao, A.; Nakajima, K.I.; Kimura, K. Retinoic acid modulates lipid accumulation glucose concentration dependently through inverse regulation of SREBP-1 expression in 3T3L1 adipocytes. Genes Cells 2017, 22, 568–582. [Google Scholar] [CrossRef]
- Ahmad, R.; Bahman, F.; Kochumon, S.P.; Jacob, T.K.; Alrashed, F.; Wilson, A.; Arefanian, H.; Thomas, R.S.; Akhter, N.; Al-Mansour, N.; et al. 162-OR: Sustained TNF-α transcription triggered in obesity through H3K9/K18 acetylation is associated with metabolic impairments. Diabetes 2024, 73, 162. [Google Scholar] [CrossRef]
- Ketema, E.B.; Lopaschuk, G.D. The role of acetylation in obesity-induced cardiac metabolic alterations. J. Pharm. Pharm. Sci. 2024, 27, 13080. [Google Scholar] [CrossRef]
- Ambele, M.A.; Dhanraj, P.; Giles, R.; Pepper, M.S. Adipogenesis: A complex interplay of multiple molecular determinants and pathways. Int. J. Mol. Sci. 2020, 21, 4283. [Google Scholar] [CrossRef]
- Kim, H.-J.; Lee, J.; Chung, M.-Y.; Park, S.H.; Park, J.H.; Choi, H.-K.; Hwang, J.-T. Tamarixetin abrogates adipogenesis through inhibiting p300/CBP-associated factor acetyltransferase activity in 3T3-L1 preadipocyte cells. J. Med. Food 2022, 25, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Totani, N.; Tateishi, S.; Takimoto, T.; Maeda, Y.; Sasaki, H. Gallic acid glycerol ester promotes weight-loss in rats. J. Oleo Sci. 2011, 60, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Behera, P.K.; Devi, S.; Mittal, N. Therapeutic potential of gallic acid in obesity: Considerable shift! Obes. Med. 2023, 37, 100473. [Google Scholar] [CrossRef]
- Das, N.D.; Das, A.; Chai, Y.G. The anti-oxidative and anti-inflammatory roles of gallic acid on transcriptional regulation. In Handbook on Gallic Acid; Nova Science Publishers, Inc.: New York, NY, USA, 2013. [Google Scholar]
- Kim, M.J.; Seong, A.R.; Yoo, J.Y.; Jin, C.H.; Lee, Y.H.; Kim, Y.J.; Lee, J.; Jun, W.J.; Yoon, H.G. Gallic acid, a histone acetyltransferase inhibitor, suppresses β-amyloid neurotoxicity by inhibiting microglial-mediated neuroinflammation. Mol. Nutr. Food Res. 2011, 55, 1798–1808. [Google Scholar] [CrossRef]
- Bora-Tatar, G.; Dayangaç-Erden, D.; Demir, A.S.; Dalkara, S.; Yelekçi, K.; Erdem-Yurter, H. Molecular modifications on carboxylic acid derivatives as potent histone deacetylase inhibitors: Activity and docking studies. Bioorg. Med. Chem. 2009, 17, 5219–5228. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Rt (min) | Proposed Compound | Precursor (m/z) [M−H]− | MS/MS Fragments |
|---|---|---|---|---|
| 1 | 0.63 | Mucic acid gallate | 361 | 209, 191, 147, 133, 85, 71, 59 |
| 2 | 0.94 | Mucic acid gallate | 723.1 [2M−H]− | 361, 209, 191 |
| 3 | 1.76 | Galloylquinic acid isomer | 687.1 | 343, 191, 147, 85 |
| 4 | 2.54 | Gallic acid | 169 | 169, 125, 124, 123, 107, 97, 95, 81, 79, 69, 67, 53 |
| 5 | 2.73 | Galloylquinic acid isomer | 687.1 | 687, 343, 191, 169, 147, 85 |
| 6 | 2.88 | Galloylquinic acid isomer | 687.1 | 687, 343, 237, 191, 169, 147, 85 |
| 7 | 3.12 | Gallic acid derivative | 673.1 | 387, 285, 169, 133, 125, 115 |
| 8 | 3.24 | Gallic acid derivative | 387.1 | 169, 125, 129 |
| 9 | 3.32 | Gallic acid derivative | 455.0 (387) | 387, 325, 209, 191, 173, 169, 151, 129, 125 |
| 10 | 3.45 | Digalloyl glucose | 483.1 | 483, 331, 313, 271, 241, 211, 169, 168, 151, 125, 124 |
| 11 | 3.64 | Digallic acid | 321 | 169, 125 |
| 12 | 3.81 | Trigalloyl glucose isomer | 635.1 | 123, 113, 93, 68, 25 |
| 13 | 3.93 | Trigalloyl glucose isomer | 635.1 | 483, 465, 423, 313, 271, 211, 169 |
| 14 | 3.98 | Digalloyl-HHDP glucose | 785.1 | 633, 615, 300, 463, 275 |
| 15 | 4.16 | unidentified ellagitannin | 953.6 | 300 |
| 16 | 4.21 | Gallic acid derivative | 965.1 | 387, 285, 169, 133, 125 |
| 17 | 4.45 | Gallic acid derivative | 197 | 169, 125, 124, 78 |
| 17 | 4.84 | Kaempferol-3-O-glucoside | 447.1 | 479, 314, 287, 223, 213, 163 |
| 19 | 5.29 | Kaempferol-3-O-deoxyhexoside | 431.1 | 285, 284, 255, 229, 227 |
| 20 | 5.62 | unidentified gallic acid | 461.1 | 313, 271, 241, 211, 169, 151, 125, 124 |
| 21 | 5.73 | unidentified gallic acid | 461.1 | 313, 211, 189, 169, 161, 151, 147, 125, 124, 123 |
| 22 | 5.82 | 613.1 | 465, 461, 313, 271, 211, 169 |
| Compound Name | Phenolic Acid (mg/g) |
|---|---|
| Gallic acid (GA) | 13.99 ± 0.07 |
| 3-Galloylquinic acid (GQA) | 10.71 ± 0.16 |
| Compound Name | IC50 (μM) |
|---|---|
| Gallic acid (GA) | 18.32 |
| 3-Galloylquinic acid (GQA) | 83.65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.K.; Lee, Y.G.; Lee, J.-I.; Kim, M.-S.; Park, J.-H.; Hwang, J.-T.; Chung, M.-Y. Phyllanthus emblica Prevents Adipogenesis by Regulating Histone Acetylation. Foods 2025, 14, 160. https://doi.org/10.3390/foods14020160
Park SK, Lee YG, Lee J-I, Kim M-S, Park J-H, Hwang J-T, Chung M-Y. Phyllanthus emblica Prevents Adipogenesis by Regulating Histone Acetylation. Foods. 2025; 14(2):160. https://doi.org/10.3390/foods14020160
Chicago/Turabian StylePark, Seon Kyeong, Yu Geon Lee, Jae-In Lee, Min-Sun Kim, Jae-Ho Park, Jin-Taek Hwang, and Min-Yu Chung. 2025. "Phyllanthus emblica Prevents Adipogenesis by Regulating Histone Acetylation" Foods 14, no. 2: 160. https://doi.org/10.3390/foods14020160
APA StylePark, S. K., Lee, Y. G., Lee, J.-I., Kim, M.-S., Park, J.-H., Hwang, J.-T., & Chung, M.-Y. (2025). Phyllanthus emblica Prevents Adipogenesis by Regulating Histone Acetylation. Foods, 14(2), 160. https://doi.org/10.3390/foods14020160