
Maintaining the Quality and Nutritional Integrity of Chilled Cordyceps sinensis: Comparative Effects and Mechanisms of Modified Atmosphere Packaging and UV-Based Interventions

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
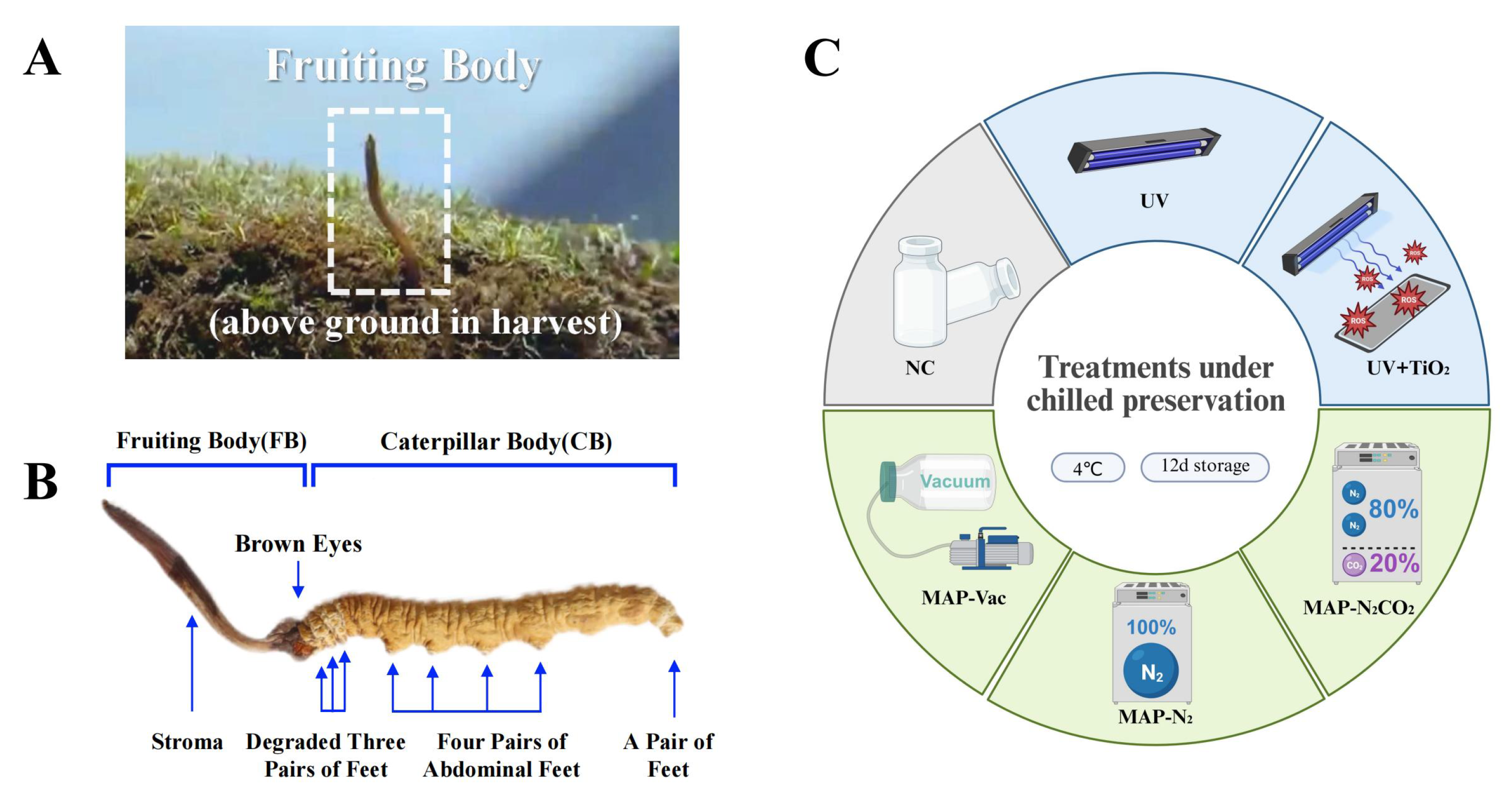
2.1.1. Fresh C. sinensis Selection and Grouping
2.1.2. C. sinensis Treatment
2.2. Methods
2.2.1. Color Quality and Visual Image
2.2.2. Sensory Evaluation Method
2.2.3. Weight Loss Measurement
2.2.4. Scanning Electron Microscopy (SEM)
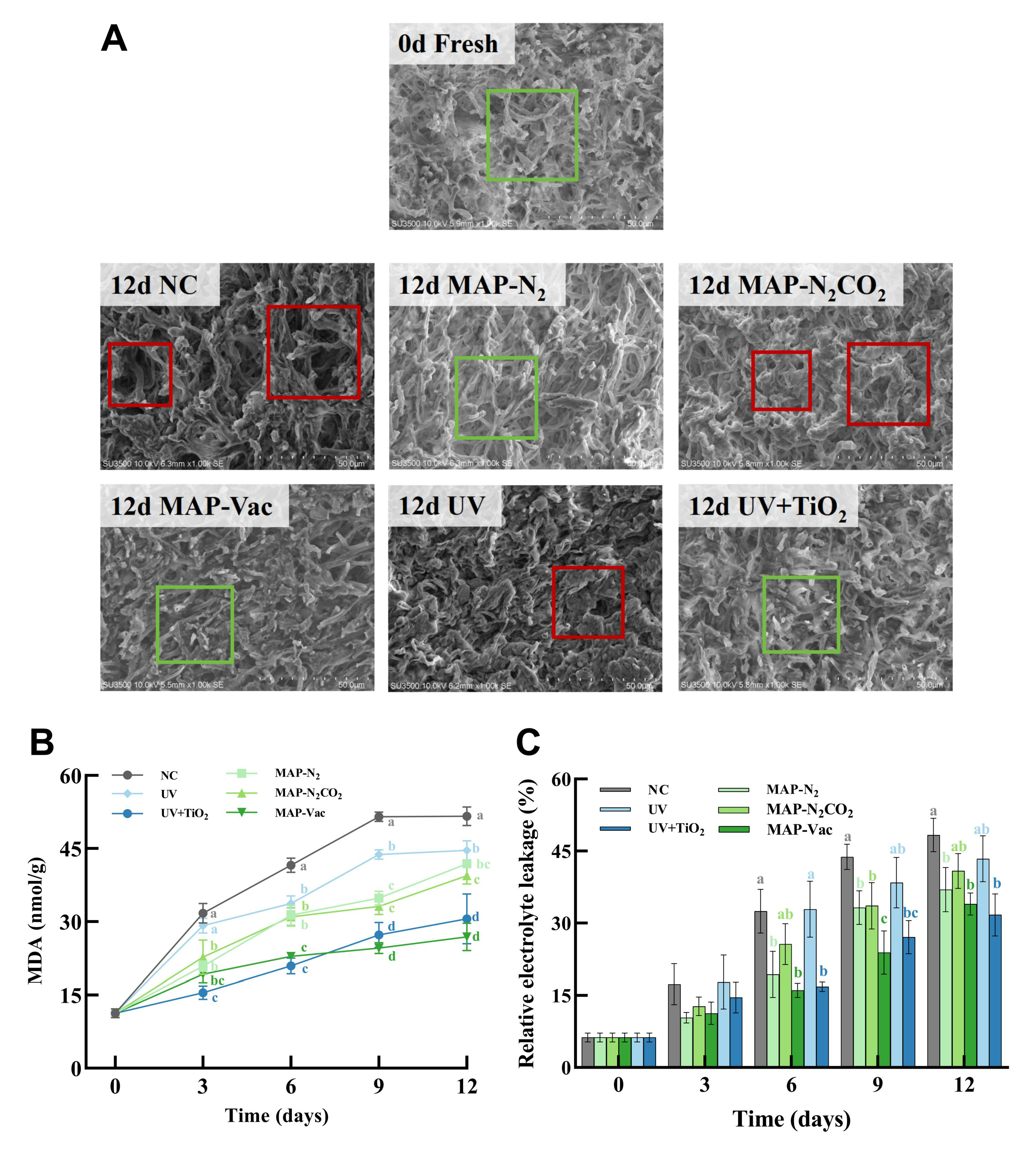
2.2.5. Malondialdehyde and Relative Electrolyte Leakage
2.2.6. Total Sugar, Amino Acid, and Nucleosides Content
2.2.7. Energy Status Assay
2.2.8. Measurement of Superoxide Dismutase, Catalase, Tyrosinase, and Polyphenol Oxidase Activities
2.2.9. Assay of Total Antioxidant Capacity
2.2.10. Bacterial Community Analysis Method
2.2.11. Statistical Analysis
3. Results
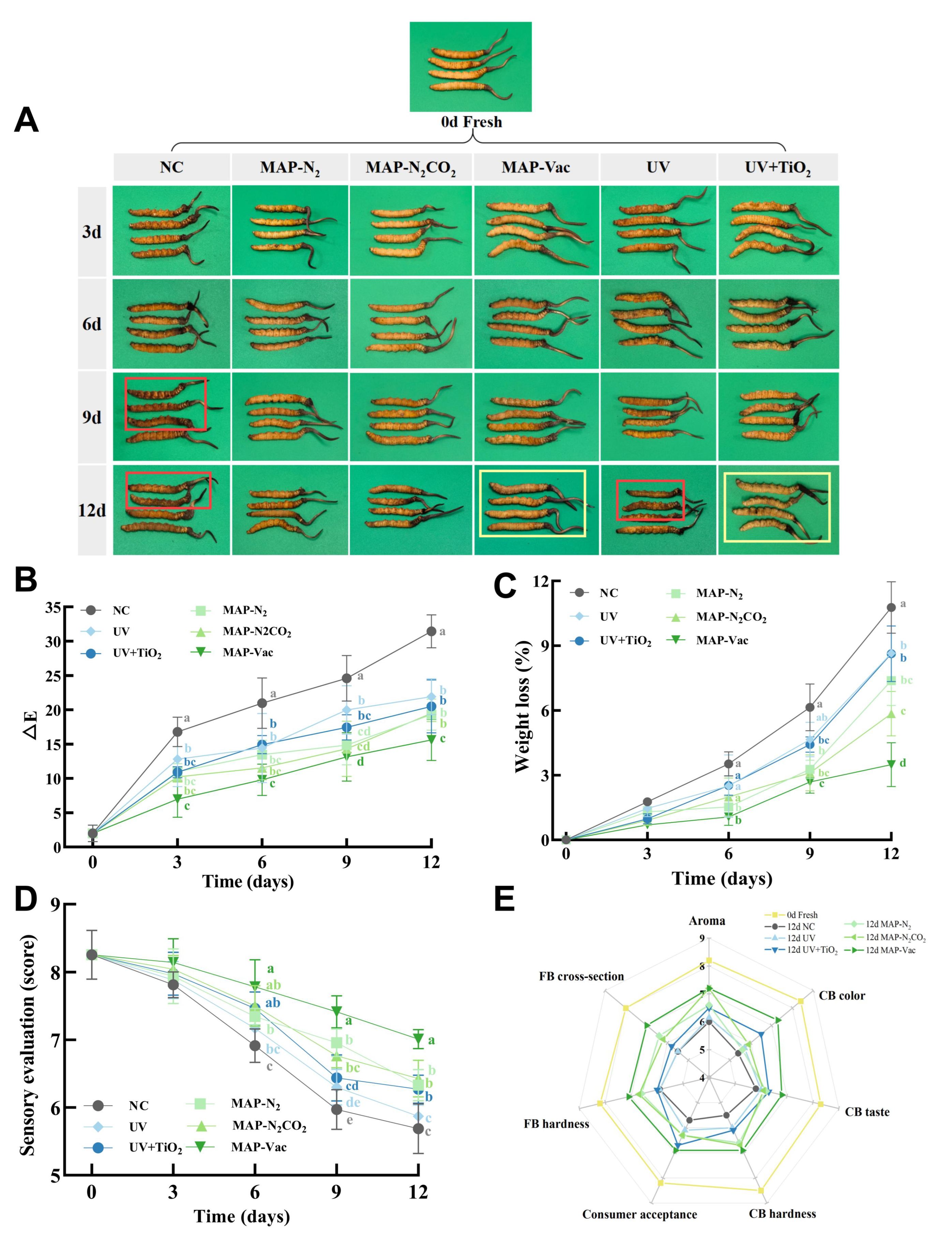
3.1. Apparent Quality Evaluation
3.1.1. Color Quality and Difference
3.1.2. Sensory Evaluation
3.1.3. Weight Loss
3.2. Microstructure Observation and Cell Integrity
3.2.1. C. sinensis Internal Hypha Structure
3.2.2. C. sinensis Hypha Cell Integrity
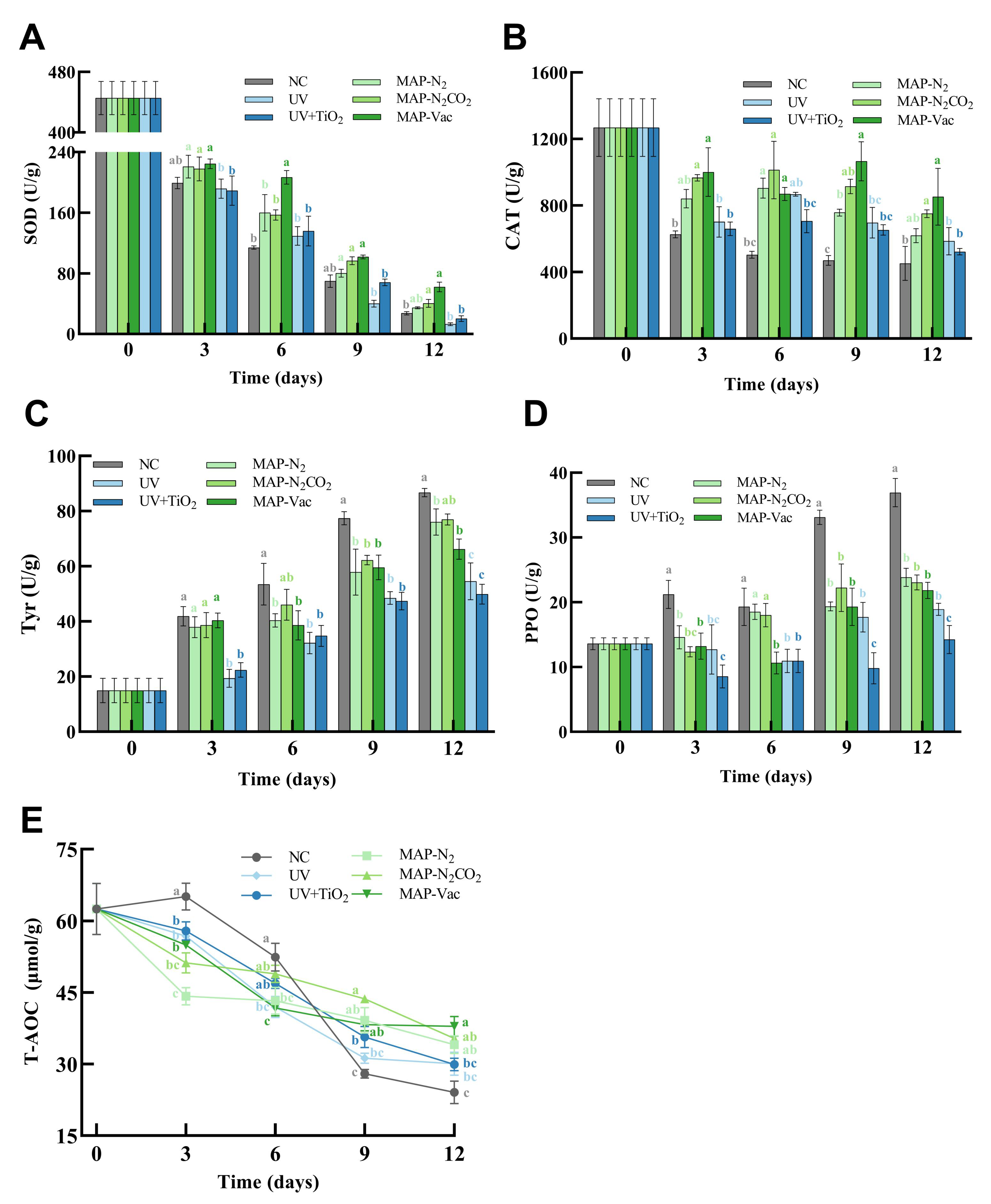
3.3. Antioxidant Capacity Evaluation
3.3.1. Antioxidant Enzyme Activities
3.3.2. Polyphenol Oxidase and Tyrosinase Enzyme Activities
3.3.3. Total Antioxidant Capacity
3.4. Nutritional Value Evaluation
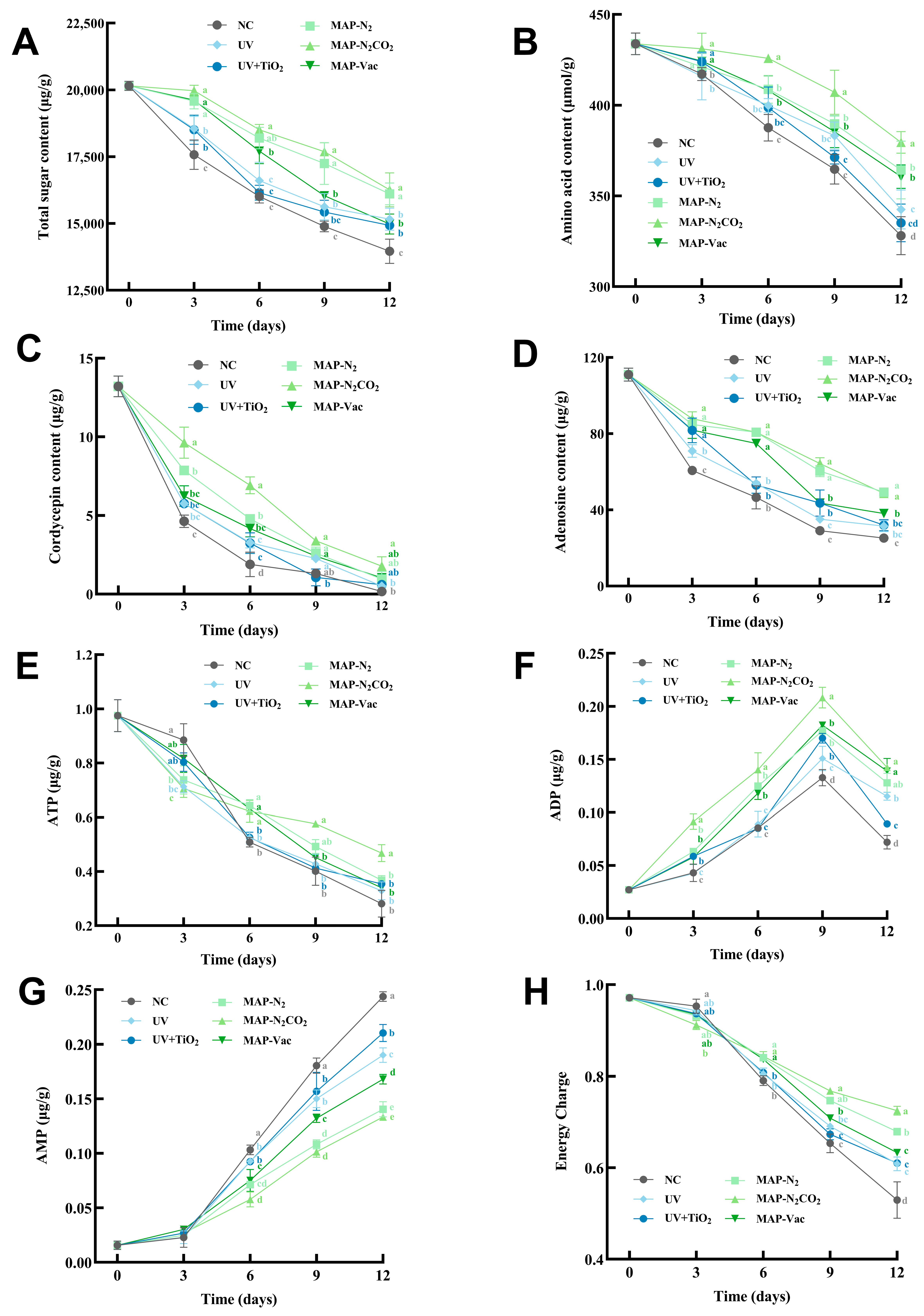
3.4.1. Dynamic Changes in Total Sugars and Total Amino Acid Content
3.4.2. Nucleosides Dynamics
3.4.3. ATP, ADP, AMP, and Energy Charge
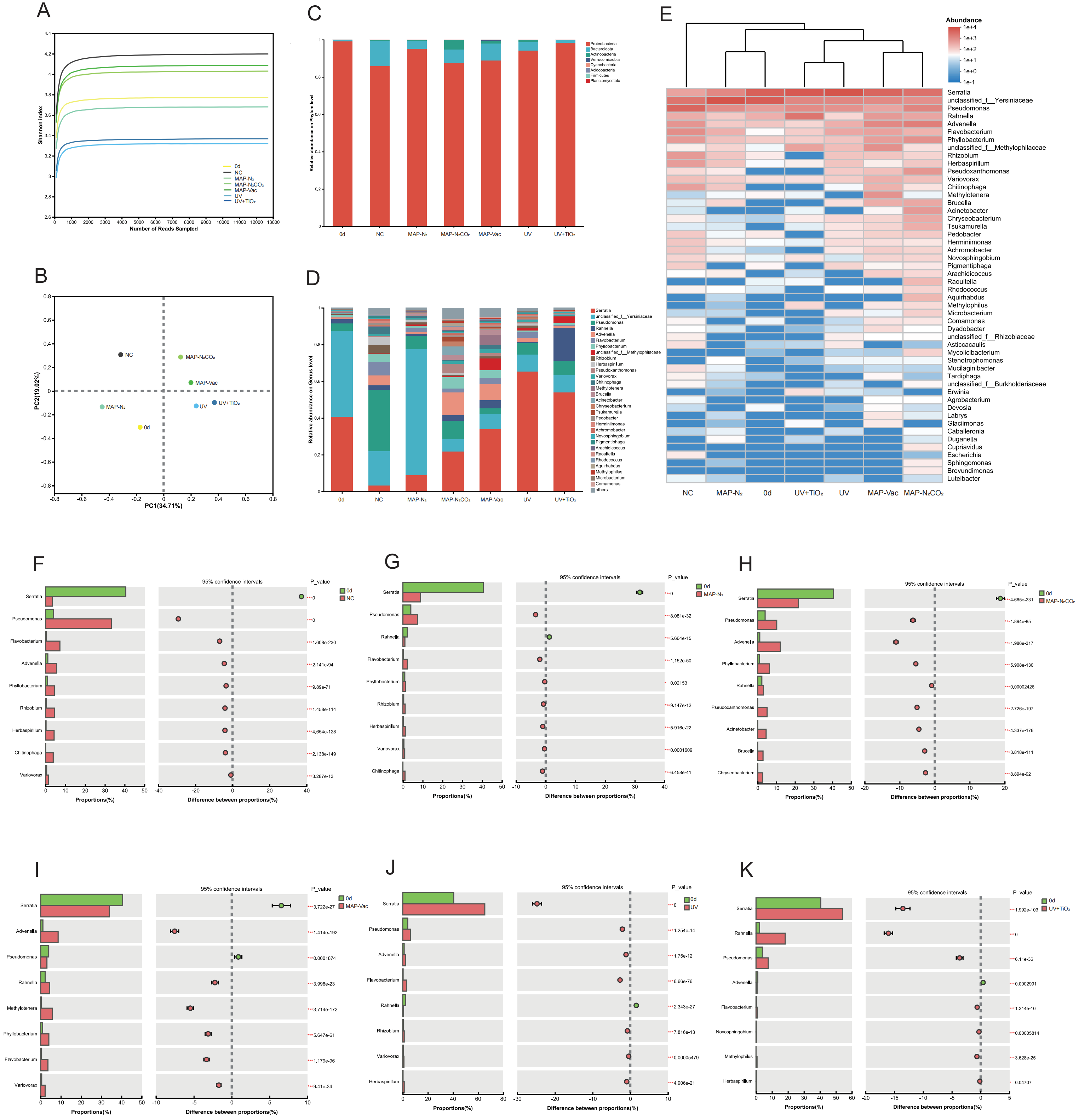
3.5. Bacterial Community Analysis
3.5.1. Bacterial Community Compositions
3.5.2. Spoilage-Associated Microbial Succession
3.6. Correlation and Difference Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADP | Adenosine diphosphate |
| AMP | Adenosine monophosphate |
| ATP | Adenosine triphosphate |
| CAT | Catalase |
| CB | Caterpillar body |
| C. sinensis | Cordyceps sinensis |
| FB | Fruiting body |
| MDA | Malondialdehyde |
| MAP | Modified atmosphere packaging |
| MAP-N2 | Modified atmosphere packaging with nitrogen |
| MAP-N2CO2 | Modified atmosphere packaging with nitrogen and carbon dioxide |
| MAP-Vac | Vacuum packaging |
| NC | Negative control |
| PCA | Principal component analysis |
| PCoA | Principal coordinates analysis |
| PPO | Polyphenol oxidase |
| REL | Relative electrolyte leakage |
| ROS | Reactive oxygen species |
| SEM | Scanning electron microscopy |
| SOD | Superoxide dismutase |
| T-AOC | Total antioxidant capacity |
| TiO2 | Titanium dioxide |
| TSC | Total sugar content |
| UV | Ultraviolet treatment |
| UV + TiO2 | Photocatalysis treatment |
References
- Liu, Y.; Wang, J.; Wang, W.; Zhang, H.; Zhang, X.; Han, C. The Chemical Constituents and Pharmacological Actions of C. sinensis. Evid.-Based Complement. Altern. Med. 2015, 2015, 575063. [Google Scholar]
- Lee, D.; Lee, W.-Y.; Jung, K.; Kwon, Y.S.; Kim, D.; Hwang, G.S.; Kim, C.-E.; Lee, S.; Kang, K.S. The Inhibitory Effect of Cordycepin on the Proliferation of MCF-7 Breast Cancer Cells, and its Mechanism: An Investigation Using Network Pharmacology-Based Analysis. Biomolecules 2019, 9, 414. [Google Scholar] [CrossRef] [PubMed]
- Shashidhar, M.G.; Giridhar, P.; Sankar, K.U.; Manohar, B. Bioactive principles from C. sinensis: A potent food supplement—A review. J. Funct. Foods 2013, 5, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Xiong, X.; Gao, S.; Dou, P.; Lv, H.; Tan, Y.; Hong, H.; Luo, Y. Enhancing Cordyceps Sinensis shelf life: The role of liquid nitrogen spray freezing in maintaining hypha structure and reducing metabolic degradation. Food Chem. 2025, 473, 142982. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.-J.; Pan, J.; Zhao, B.; Liang, J.; Wu, Z.-Y.; Yang, J.-J. Comparisons on enhancing the immunity of fresh and dry C. militaris in vivo and in vitro. J. Ethnopharmacol. 2013, 149, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Ren, H.; Chen, X.; Ma, F.; Yu, G.; Chen, M.; Jiang, F. Short-term anaerobic treatment combined with perforation mediated MAP on the quality of A. bisporus mushroom. Postharvest Biol. Technol. 2021, 176, 111518. [Google Scholar] [CrossRef]
- Tao, F.; Zhang, M.; Yu, H.-Q. Effect of vacuum cooling on physiological changes in the antioxidant system of mushroom under different storage conditions. J. Food. Eng. 2007, 79, 1302–1309. [Google Scholar] [CrossRef]
- Wang, G.; Ma, F.; Zeng, L.; Bai, Y.; Wang, H.; Xu, X.; Zhou, G. Modified atmosphere packaging decreased P. fragi cell metabolism and extracellular proteolytic activities on meat. Food Microbiol. 2018, 76, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, Y.; Wang, Y.; Bu, H.; Dong, T. Changes in cell wall metabolism and flavor qualities of mushrooms (A. bernardii) under EMAP treatments during storage. Food Packag. Shelf Life 2021, 29, 100732. [Google Scholar] [CrossRef]
- Feng, Y.; Xu, H.; Sun, Y.; Xia, R.; Hou, Z.; Li, Y.; Wang, Y.; Pan, S.; Li, L.; Zhao, C.; et al. Effect of light on quality of preharvest and postharvest edible mushrooms and its action mechanism: A review. Trends Food Sci. Tech. 2023, 139, 104119. [Google Scholar] [CrossRef]
- Zheng, Y.; Duan, L.; Li, J.; Zhang, P.; Jiang, Y.; Yang, X.; Li, X.; Jia, X. Photocatalytic titanium dioxide reduces postharvest decay of nectarine fruit packaged in different materials through modulating central carbon and energy metabolisms. Food Chem. 2023, 433, 137357. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wen, X.; Liang, S.; Sun, X.; Ma, H.; Zhang, Y.; Tan, Y.; Hong, H.; Luo, Y. Enhancing bighead carp cutting: Chilled storage insights and machine vision-based segmentation algorithm development. Food Chem. 2024, 450, 139280. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Jin, Y.; Jiang, R.; Xu, Y.; Zhang, Y.; Luo, Y.; Huang, J.; Wang, K.; Liu, Z. Dynamic changes in the metabolite profile and taste characteristics of Fu brick tea during the manufacturing process. Food Chem. 2021, 344, 128576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Gong, H.; Liu, Y. Effects of postharvest coating using chitosan combined with natamycin on physicochemical and microbial properties of sweet cherry during cold storage. Int. J. Biol. Macromol. 2022, 214, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Liu, H.; Li, L.; Wang, X.; Kitazawa, H.; Guo, Y. Improving the property of a reproducible bioplastic film of glutenin and its application in retarding senescence of postharvest A. bisporus. Food Biosci. 2022, 48, 101796. [Google Scholar] [CrossRef]
- Feng, L.; Zhang, N.; Wang, J.; Jiang, X.; Han, J.; Li, L.; Kitazawa, H.; Wang, X.; Chen, C.; Guo, Y. Sustained release of a novel bilayer packaging film loaded with binary microemulsion of melatonin/pummelo essential oil and its regulation of postharvest energy metabolism in A. bisporus. Food Control 2024, 161, 110396. [Google Scholar] [CrossRef]
- Ye, T.; Chen, X.; Chen, Z.; Yao, H.; Wang, Y.; Lin, L.; Lu, J. Quality and microbial community of high pressure shucked crab (Eriocheir sinensis) meat stored at 4 °C. J. Food Process Pres. 2021, 45, e15330. [Google Scholar] [CrossRef]
- Tian, Y.; Li, D.; Luo, W.; Zhu, Z.; Li, W.; Qian, Z.; Li, G.; Sun, D.-W. Rapid freezing using atomized liquid nitrogen spray followed by frozen storage below glass transition temperature for C. sinensis preservation: Quality attributes and storage stability. Lwt-Food Sci. Technol. 2020, 123, 109066. [Google Scholar] [CrossRef]
- Gao, M.; Feng, L.; Jiang, T. Browning inhibition and quality preservation of button mushroom (A. bisporus) by essential oils fumigation treatment. Food Chem. 2014, 149, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Feng, J.; Wei, W.; Ban, Z.; Xu, X.; Zhao, Y. Effect of 1-MCP and Controlled Atmosphere Storage on Preservation of Apricot (P. armeniaca L. cv. Saimaiti). Food Sci. 2013, 34, 287–290. [Google Scholar]
- Yinxin, L.; Can, Z.; Menglu, H.; Cui, S.; Jinping, C.; Jingyu, W.; Huang, L. Effect of cold atmospheric plasma on the gray mold rot of postharvest mulberry fruit. Food Control 2022, 137, 108906. [Google Scholar] [CrossRef]
- Ząbek, K.; Miciński, J.; Milewski, S.; Sobczak, A. Effect of modified atmosphere packaging and vacuum packaging on quality characteristics of lamb meat. Arch. Anim. Breed. 2021, 64, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Atallah, A.; El-Deeb, A.M.; Mohamed, E.N. Shelf-life of Domiati cheese under modified atmosphere packaging. J. Dairy Sci. 2021, 104, 8568–8581. [Google Scholar] [CrossRef] [PubMed]
- Mou, Z.-L.; Wang, C.-X.; Wei, W.; Zhao, Y.-T.; Wang, L.; Ji, S.-J.; Shan, W.; Kuang, J.-F.; Lu, W.-J.; Chen, Y.-L.; et al. Modified atmosphere packaging delays postharvest leaf senescence of Chinese flowering cabbage via suppressing chloroplast dismantling and alleviating mitochondrial oxidative damage. Food Packag. Shelf Life 2023, 39, 101136. [Google Scholar] [CrossRef]
- Yang, Q.; Hu, Y.; Wang, Y.; Xu, B.; Zhou, C.; Adhikari, B.; Liu, J.; Xu, T.; Wang, B. Atmosphere-controlled high-voltage electrospray for improving conductivity, flexibility, and antibacterial properties of chitosan films. Food Res. Int. 2025, 200, 115450. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Bashri, G.; Prasad, S.M.; Singh, V.P. Kinetin Alleviates UV-B-Induced Damage in S. lycopersicum: Implications of Phenolics and Antioxidants. J. Plant Growth Regul. 2019, 38, 831–841. [Google Scholar] [CrossRef]
- Liu, Q.; Cui, X.; Song, Z.; Kong, W.; Kang, Y.; Kong, W.; Ng, T.B. Coating shiitake mushrooms (L. edodes) with a polysaccharide from Oudemansiella radicata improves product quality and flavor during postharvest storage. Food Chem. 2021, 352, 129357. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Khan, A.S.; Nawaz, A.; Anjum, M.A.; Naz, S.; Ejaz, S.; Hussain, S. Aloe vera gel coating delays postharvest browning and maintains quality of harvested litchi fruit. Postharvest Biol. Technol. 2019, 157, 110960. [Google Scholar] [CrossRef]
- Cui, X.; Ding, Y.; Sun, C.; Li, X.; Yuan, S.; Guo, F.; Zeng, X.; Fan, X.; Sun, S. Improving the storage quality and suppressing off-flavor generation of winter jujube by precise micro-perforated MAP. Front. Plant Sci. 2024, 15, 1372638. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Jiang, P.; Chen, D.; Zhang, C.; Guo, F.; Nie, X.; Wu, D.; Fan, X.; Zhao, H. Improving the storage quality and antioxidant capacity of postharvest winter jujube by laser microporous modified atmosphere packaging. Sci. Hortic. 2024, 337, 113477. [Google Scholar] [CrossRef]
- Xu, C.; Wu, F.; Zou, Z.; Mao, L.; Lin, S. Discovery of the chemical constituents, structural characteristics, and pharmacological functions of Chinese caterpillar fungus. Open Chem. 2023, 21, 20220337. [Google Scholar] [CrossRef]
- Phan, C.-W.; Wang, J.-K.; Cheah, S.-C.; Naidu, M.; David, P.; Sabaratnam, V. A review on the nucleic acid constituents in mushrooms: Nucleobases, nucleosides and nucleotides. Crit. Rev. Biotechnol. 2018, 38, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.-C.; Zhan, Q.; Cai, L.; Wu, J.; Yang, L.; Sun, S.; Liang, H.; Li, Z. Investigation into the content change and distribution of active components in C. sinensis with growth cycle by direct TOF-SIMS detection. Microchem. J. 2021, 164, 106026. [Google Scholar] [CrossRef]
- Dulla, C.G.; Dobelis, P.; Pearson, T.; Frenguelli, B.G.; Staley, K.J.; Masino, S.A. Adenosine and ATP link PCO2 to cortical excitability via pH. Neuron 2005, 48, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Verástegui, L.L.; Osorio-Córdoba, J.; de León-Sánchez, F.D.; López-Ramos, A.M.; Pérez-Salvador, B.R.; Rivera-Cabrera, F.; Mendoza-Espinoza, J.A.; Pelayo-Zaldívar, C. Thermal treatment and modified atmosphere packaging delay chilling injury and preserve the quality of cactus stems (O. ficus-indica Mill.) cv Atlixco. J. Food Biochem. 2019, 43, e12773. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Qiu, Z.; Yin, M.; Zhang, L.; Zhao, Y.; Zhang, R.; Shi, W.; Wang, X. Key non-volatile taste components and characteristics in abdominal muscle of E. sinensis under various thermal processing methods. Food Chem. 2024, 464, 141902. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.L.; Sarti, A.C.; Di Virgilio, F. Extracellular nucleotides and nucleosides as signalling molecules. Immunol. Lett. 2019, 205, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Lin, H.; Chen, Y.; Zhang, H.; Wang, X.; Lin, M.; Chen, Y.; Wang, H.; Fan, Z.; Lin, Y. Respiratory and energy metabolisms participate in the disease occurrence of fresh Chinese olive caused by P. microspora. Postharvest Biol. Tec. 2023, 205, 112514. [Google Scholar] [CrossRef]
- Fang, D.; Wang, H.; Deng, Z.; Kimatu, B.M.; Pei, F.; Hu, Q.; Ma, N. Nanocomposite packaging regulates energy metabolism of mushrooms (F. filiformis) during cold storage: A study on mitochondrial proteomics. Postharvest Biol. Tec. 2022, 193, 112046. [Google Scholar] [CrossRef]
- Hou, F.; Yi, F.; Song, L.; Zhan, S.; Zhang, R.; Han, X.; Sun, X.; Liu, Z. Bacterial community dynamics and metabolic functions prediction in white button mushroom (A. bisporus) during storage. Food Res. Int. 2023, 171, 113077. [Google Scholar] [CrossRef] [PubMed]
- Pogorzelska-Nowicka, E.; Hanula, M.; Wojtasik-Kalinowska, I.; Stelmasiak, A.; Zalewska, M.; Półtorak, A.; Wierzbicka, A. Packaging in a High O2 or Air Atmospheres and in Microperforated Films Effects on Quality of Button Mushrooms Stored at Room Temperature. Agriculture 2020, 10, 479. [Google Scholar] [CrossRef]
- Bhavya, M.L.; Hebbar, H.U. Pulsed light processing of foods for microbial safety. Food Qual. Saf. 2017, 1, 187–201. [Google Scholar] [CrossRef]
- McKee, L.S.; Martínez-Abad, A.; Ruthes, A.C.; Vilaplana, F.; Brumer, H. Focused Metabolism of β-Glucans by the Soil Bacteroidetes Species C. pinensis. Appl. Environ. Microb. 2019, 85, e02231-18. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Huang, Y.; Sun, C.; Wang, D.; Xu, W. A Water-Dispersible Carboxylated Carbon Nitride Nanoparticles-Based Electrochemical Platform for Direct Reporting of Hydroxyl Radical in Meat. Foods 2021, 11, 40. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Aroma (S1) | CB Hardness (S2) | CB Color (S3) | CB Taste (S4) | FB Hardness (S5) | FB Cross Section (S6) | Consumer Acceptance (S7) | Sensory Score |
|---|---|---|---|---|---|---|---|---|
| 10–8 | Pleasant mushroom-like aroma | Very firm | Yellowish- white to golden-yellow | Sweet taste, lasting flavor | Very firm | Full of white content | Very satisfied | - |
| 8–6 | Mild mushroom-like aroma | Firm, less springy | Golden-yellow | Sweet taste, slightly weak and short-lived flavor | Firm, less springy | Quite full of white content | Satisfied | - |
| 6–4 | Mild and Neutral | Moderately firm | Brownish- yellow | Neutral and no off flavor | Moderately firm | Somewhat lacking white content | Acceptable | - |
| 4–2 | Slight off smell | Soft, lacks firmness | Tawny yellow | Bland or slightly off flavor | Soft, lacks firmness | Less white content | Unacceptable | - |
| 2–0 | Unpleasant, off odor | Very soft | Dark brown | Obvious off flavor | Very soft | No white content | Offensive | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.; Lv, H.; Lin, Y.; Xiong, X.; Tan, Y.; Hong, H.; Luo, Y. Maintaining the Quality and Nutritional Integrity of Chilled Cordyceps sinensis: Comparative Effects and Mechanisms of Modified Atmosphere Packaging and UV-Based Interventions. Foods 2025, 14, 2611. https://doi.org/10.3390/foods14152611
Huang T, Lv H, Lin Y, Xiong X, Tan Y, Hong H, Luo Y. Maintaining the Quality and Nutritional Integrity of Chilled Cordyceps sinensis: Comparative Effects and Mechanisms of Modified Atmosphere Packaging and UV-Based Interventions. Foods. 2025; 14(15):2611. https://doi.org/10.3390/foods14152611
Chicago/Turabian StyleHuang, Tianzhuo, Huanzhi Lv, Yubo Lin, Xin Xiong, Yuqing Tan, Hui Hong, and Yongkang Luo. 2025. "Maintaining the Quality and Nutritional Integrity of Chilled Cordyceps sinensis: Comparative Effects and Mechanisms of Modified Atmosphere Packaging and UV-Based Interventions" Foods 14, no. 15: 2611. https://doi.org/10.3390/foods14152611
APA StyleHuang, T., Lv, H., Lin, Y., Xiong, X., Tan, Y., Hong, H., & Luo, Y. (2025). Maintaining the Quality and Nutritional Integrity of Chilled Cordyceps sinensis: Comparative Effects and Mechanisms of Modified Atmosphere Packaging and UV-Based Interventions. Foods, 14(15), 2611. https://doi.org/10.3390/foods14152611