
CRISPR-Cas12a/Aurora Deoxyribozyme Cascade: A Label-Free Ultrasensitive Platform for Rapid Salmonella Detection

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Bacterial Culture and Extraction
2.3. RPA Amplification and Validation
2.4. Label-Free Detection of Salmonella by Aurora-Based CRISPR/Cas12a Sensor
2.5. Preparation of Spiked Samples
3. Results and Discussion
3.1. Novel Strategy for Highly Sensitive Detection of Salmonella typhimurium Based on Synergistic Action of CRISPR-Cas12a and Aurora Fluorescence System
3.2. Comprehensive Validation of the CRISPR-Cas12a/Aurora-Based Detection System for Salmonella typhimurium
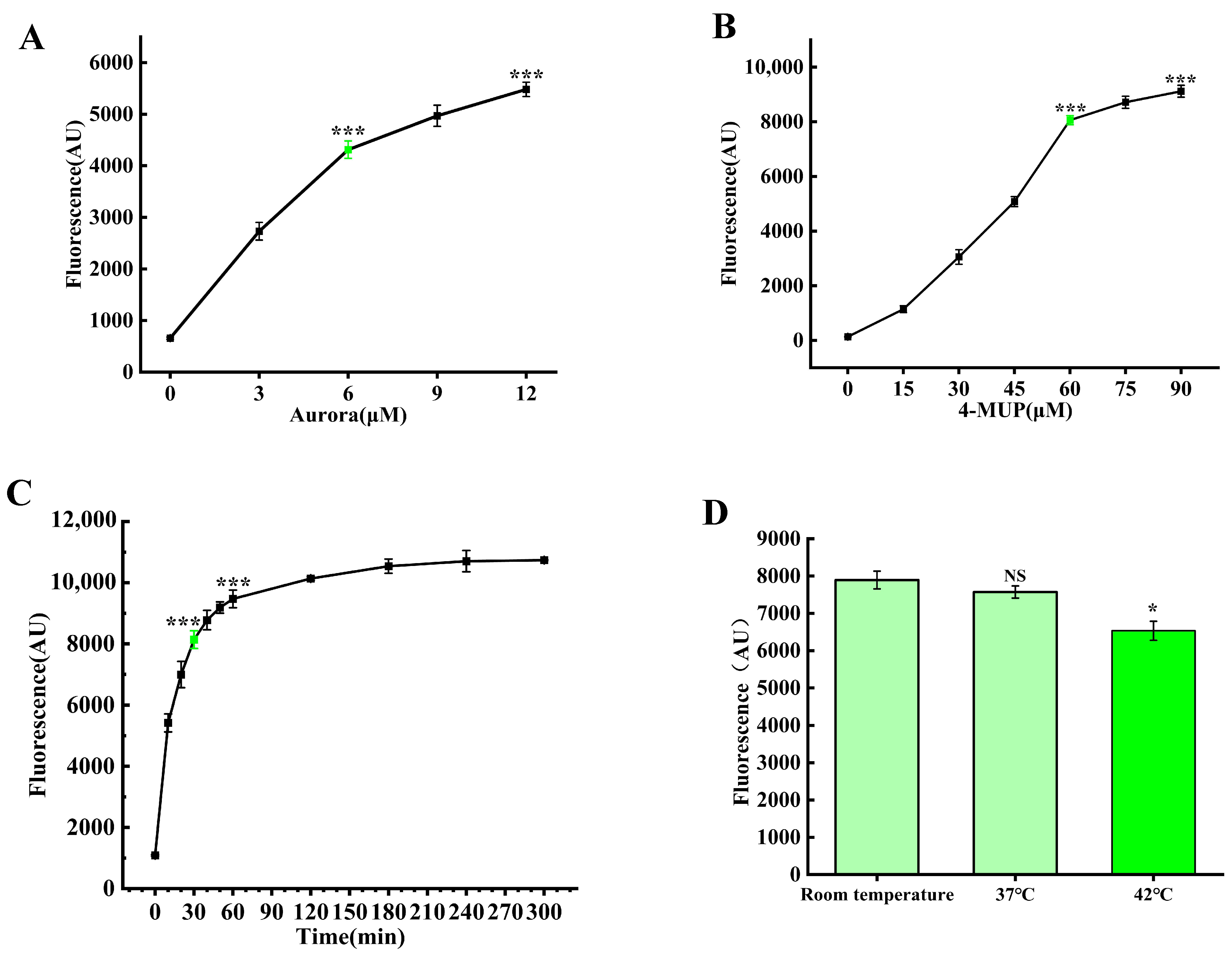
3.3. Optimization of Reaction Conditions
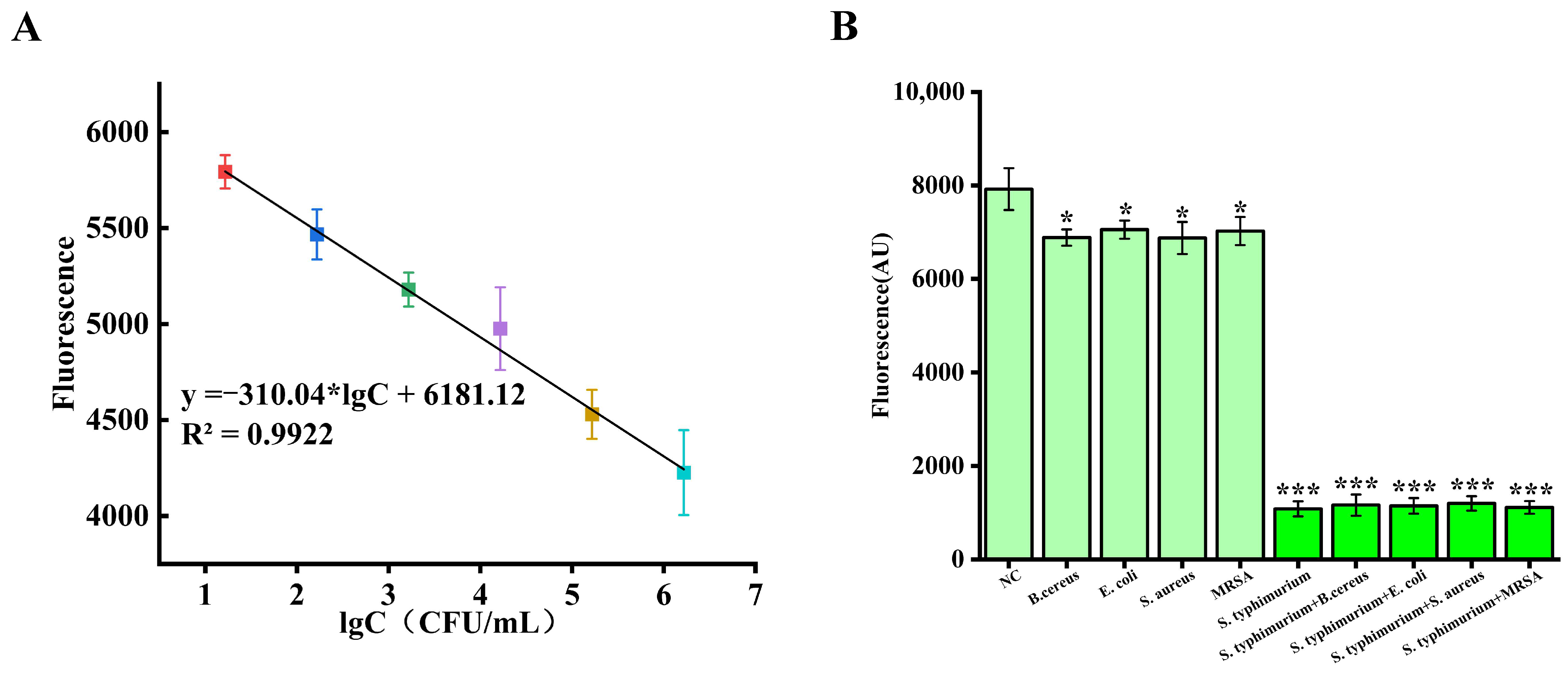
3.4. Label-Free Detection of Salmonella Using Aurora-Based CRISPR-Cas12a Sensors
3.5. Application in Real Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maskery, B.; Ochiai, R.L.; Lee, J.S.; Mogasale, V.V.; Ramani, E.; Kim, Y.E.; Park, J.K.; Wierzba, T.F. Burden of typhoid fever in low-income and middle-income countries: A systematic, literature-based update with risk-factor adjustment. Lancet Glob. Health 2014, 2, e570–e580. [Google Scholar]
- Li, G.; Huang, Y.; Duan, M.; Xing, K.; You, X.; Zhou, H.; Liu, Y.; Liu, C.; Liu, D.; Lai, W. Biosensing multiplexer based on immunochromatographic assay for rapid and high-throughput classification of Salmonella serogroups. Sens. Actuators B Chem. 2019, 282, 317–321. [Google Scholar] [CrossRef]
- Xiang, X.; Shang, Y.; Ye, Q.; Li, F.; Zhang, J.; Zhou, B.; Suo, H.; Chen, M.; Gu, Q.; Ding, Y.; et al. A Salmonella serogroup rapid identification system for food safety based on high throughput microfluidic chip combined with recombinase aided amplification. Sens. Actuators B Chem. 2022, 357, 131402. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, G.; Li, X.; Xu, Z.; Lei, H.; Shen, X. Development of rapid and easy detection of Salmonella in food matrics using RPA-CRISPR/Cas12a method. LWT 2022, 162, 113443. [Google Scholar] [CrossRef]
- Duan, M.; Li, B.; Zhao, Y.; Liu, Y.; Liu, Y.; Dai, R.; Li, X.; Jia, F. A CRISPR/Cas12a-mediated, DNA extraction and amplification-free, highly direct and rapid biosensor for Salmonella Typhimurium. Biosens. Bioelectron. 2023, 219, 114823. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O'Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease “Burden of Illness” Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jia, F.; Sun, Y.; Fang, W.; Li, Y.; Chen, J.; Fu, Y. An electrochemical sensing method based on CRISPR/Cas12a system and hairpin DNA probe for rapid and sensitive detection of Salmonella Typhimurium. Sens. Actuators B Chem. 2022, 369, 132301. [Google Scholar] [CrossRef]
- Chen, L.; Hu, M.; Zhou, X. Trends in developing one-pot CRISPR diagnostics strategies. Trends Biotechnol. 2024, 43, 98–110. [Google Scholar] [CrossRef]
- Chen, B.; Li, Y.; Xu, F.; Yang, X. Powerful CRISPR-based biosensing techniques and their integration with microfluidic platforms. Front. Bioeng. Biotechnol. 2022, 10, 851712. [Google Scholar] [CrossRef]
- Liu, P.; Lin, Y.; Zhuo, X.; Zeng, J.; Chen, B.; Zou, Z.; Liu, G.; Xiong, E.; Yang, R. Universal crRNA Acylation Strategy for Robust Photo-Initiated One-Pot CRISPR–Cas12a Nucleic Acid Diagnostics. Angew. Chem. Int. Ed. 2024, 63, e202401486. [Google Scholar] [CrossRef]
- Qin, C.; Liu, J.; Zhu, W.; Zeng, M.; Xu, K.; Ding, J.; Zhou, H.; Zhu, J.; Ke, Y.; Li, L.Y.; et al. One-pot visual detection of african swine fever virus using CRISPR-Cas12a. Front. Vet. Sci. 2022, 9, 962438. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Macaluso, N.C.; Pizzano, B.L.; Cash, M.N.; Spacek, J.; Karasek, J.; Miller, M.R.; Lednicky, J.A.; Dinglasan, R.R.; Salemi, M.; et al. A thermostable Cas12b from Brevibacillus leverages one-pot discrimination of SARS-CoV-2 variants of concern. EBioMedicine 2022, 77, 103926. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Duan, J.-J.; Wei, X.-Y.; Hu, H.; Wang, Y.-B.; Jia, P.-P.; Pei, D.-S. Generation and application of a novel high-throughput detection based on RPA-CRISPR technique to sensitively monitor pathogenic microorganisms in the environment. Sci. Total Environ. 2022, 838, 156048. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zong, N.; Ye, F.; Mei, Y.; Qu, J.; Jiang, X. Dual-CRISPR/Cas12a-assisted RT-RAA for ultrasensitive SARS-CoV-2 detection on automated centrifugal microfluidics. Anal. Chem. 2022, 94, 9603–9609. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Liu, S.; Xu, Y.; Li, A.; Wu, W.; Liang, M.; Niu, G.; Wang, Z.; Wang, T. CRISPR/Cas12a technology combined with RPA for rapid and portable SFTSV detection. Front. Microbiol. 2022, 13, 754995. [Google Scholar] [CrossRef]
- Hille, F.; Richter, H.; Wong, S.P.; Bratovič, M.; Ressel, S.; Charpentier, E. The biology of CRISPR-Cas: Backward and forward. Cell 2018, 172, 1239–1259. [Google Scholar] [CrossRef]
- Wright, A.V.; Nuñez, J.K.; Doudna, J.A. Biology and applications of CRISPR systems: Harnessing nature’s toolbox for genome engineering. Cell 2016, 164, 29–44. [Google Scholar] [CrossRef]
- Smith, C.W. Development of Nucleic Acid Diagnostics for Targeted and Non-Targeted Biosensing. Ph.D. Thesis, State University of New York, Albany, NY, USA, 2022. [Google Scholar]
- Yang, H.; Gao, P.; Rajashankar, K.R.; Patel, D.J. PAM-dependent target DNA recognition and cleavage by C2c1 CRISPR-Cas endonuclease. Cell 2016, 167, 1814–1828. [Google Scholar] [CrossRef]
- Sohail, M.; Xie, S.; Zhang, X.; Li, B. Methodologies in visualizing the activation of CRISPR/Cas: The last mile in developing CRISPR-Based diagnostics and biosensing–a review. Anal. Chim. Acta 2022, 1205, 339541. [Google Scholar] [CrossRef]
- Yan, F.; Wang, W.; Zhang, J. CRISPR-Cas12 and Cas13: The lesser known siblings of CRISPR-Cas9. Cell Biol. Toxicol. 2019, 35, 489–492. [Google Scholar] [CrossRef]
- Volek, M.; Kurfürst, J.; Drexler, M.; Svoboda, M.; Srb, P.; Veverka, V.; A Curtis, E. Aurora: A fluorescent deoxyribozyme for high-throughput screening. Nucleic Acids Res. 2024, 52, 9049–9061. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, C. Designing in vivo active DNAzymes. Nat. Chem. 2021, 13, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, S.; Sintim, H.O. Biomolecule detection with peroxidase-mimicking DNAzymes; expanding detection modality with fluorogenic compounds. Mol. Biosyst. 2009, 6, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wang, Q.; Chen, J.; Huang, L.; Zhang, H.; Rong, K.; Dong, S. Biomimetic design for enhancing the peroxidase mimicking activity of hemin. Nanoscale 2019, 11, 12603–12609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, Y.; Wu, Y.; Chang, Y.; Liu, M. CRISPR-Cas systems: From gene scissors to programmable biosensors. TrAC Trends Anal. Chem. 2021, 137, 116210. [Google Scholar] [CrossRef]
- Jing, S.; Liu, Q.; Jin, Y.; Li, B. Dimeric G-quadruplex: An effective nucleic acid scaffold for lighting up thioflavin T. Anal. Chem. 2020, 93, 1333–1341. [Google Scholar] [CrossRef]
- Zheng, Q.; Mikš-Krajnik, M.; Yang, Y.; Xu, W.; Yuk, H.-G. Real-time PCR method combined with immunomagnetic separation for detecting healthy and heat-injured Salmonella Typhimurium on raw duck wings. Int. J. Food Microbiol. 2014, 186, 6–13. [Google Scholar] [CrossRef]
- Wu, W.; Li, J.; Pan, D.; Li, J.; Song, S.; Rong, M.; Li, Z.; Gao, J.; Lu, J. Gold nanoparticle-based enzyme-linked antibody-aptamer sandwich assay for detection of Salmonella Typhimurium. ACS Appl. Mater. Interfaces 2014, 6, 16974–16981. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Q.; Zhou, M.; Li, C.; Yan, C.; Huang, L.; Qin, P. Development of a CRISPR/Cas9-integrated lateral flow strip for rapid and accurate detection of Salmonella. Food Control 2022, 142, 109203. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, K.; Yin, H.; Li, Q.; Wang, L.; Liu, Z. Detection of Salmonella and several common Salmonella serotypes in food by loop-mediated isothermal amplification method. Food Sci. Hum. Wellness 2015, 4, 75–79. [Google Scholar] [CrossRef]
- Jia, F.; Li, B.; He, Y.; Shen, Y.; Chen, J.; Li, X.; Li, Y. An amplification-free CRISPR-SERS biosensor for specific, sensitive and rapid detection of Salmonella Typhimurium in poultry. LWT 2023, 189, 115476. [Google Scholar] [CrossRef]
- Yin, L.; Duan, N.; Chen, S.; Yao, Y.; Liu, J.; Ma, L. Ultrasensitive pathogenic bacteria detection by a smartphone-read G-quadruplex-based CRISPR-Cas12a bioassay. Sens. Actuators B Chem. 2021, 347, 130586. [Google Scholar] [CrossRef]
- Cai, Q.; Wang, R.; Qiao, Z.; Yang, W. Single-digit Salmonella detection with the naked eye using bio-barcode immunoassay coupled with recombinase polymerase amplification and a CRISPR-Cas12a system. Analyst 2021, 146, 5271–5279. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | LOD (CFU/mL) | DNA Amplification | Time | Samples | References |
|---|---|---|---|---|---|
| PCR | 1 × 103 | PCR | 10 h | Raw duck wing | [28] |
| ELISA | 1 × 103 | _ | <3 h | Milk | [29] |
| Traditional CRISPR | 1×102 | PCR | <100 min | Milk | [30] |
| LAMP | 2 × 101 | LAMP | 45–70 min | Pork | [31] |
| Electrochemical CRISPR | 5.5 × 101 | PCR | <2.5 h | Chicken | [7] |
| CRISPR-SERS biosensor | 1.1 × 102 | - | 2 h | Chicken | [32] |
| smartphone-read G-quadruplex-based CRISPR-Cas12a bioassay | 1 | RPA | 3 h | Bear and Juice | [33] |
| BCA-RPA-Cas 12 a | 1 | RPA | 60 min | Milk | [34] |
| This work | 1.29 | RPA | 100 min | Chicken, Milk, Vegetable | This work |
| Samples | C(S. typhimurium) (CFU/mL) | Results (CFU/mL) | Recovery (%) | RSD (%) |
|---|---|---|---|---|
| Milk | 0 | ND a | — | — |
| Milk | 1.65 × 106 | 1.57 × 106 | 95.15 | 3.55 |
| Milk | 1.65 × 105 | 1.63 × 105 | 98.79 | 3.59 |
| Chicken | 0 | ND | — | — |
| Chicken | 1.65 × 106 | 1.49 × 106 | 93.74 | 3.60 |
| Chicken | 1.65 × 105 | 1.59 × 105 | 99.60 | 4.14 |
| Lettuce | 0 | ND | — | — |
| Lettuce | 1.65 × 106 | 1.50 × 106 | 90.91 | 4.72 |
| Lettuce | 1.65 × 105 | 1.64 × 105 | 99.40 | 3.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, C.; Tan, H.; Yu, Z.; Li, W.; Man, Y.; Zhang, Q. CRISPR-Cas12a/Aurora Deoxyribozyme Cascade: A Label-Free Ultrasensitive Platform for Rapid Salmonella Detection. Foods 2025, 14, 1892. https://doi.org/10.3390/foods14111892
Shi C, Tan H, Yu Z, Li W, Man Y, Zhang Q. CRISPR-Cas12a/Aurora Deoxyribozyme Cascade: A Label-Free Ultrasensitive Platform for Rapid Salmonella Detection. Foods. 2025; 14(11):1892. https://doi.org/10.3390/foods14111892
Chicago/Turabian StyleShi, Cong, Huimin Tan, Zhou Yu, Weilin Li, Yan Man, and Qinghai Zhang. 2025. "CRISPR-Cas12a/Aurora Deoxyribozyme Cascade: A Label-Free Ultrasensitive Platform for Rapid Salmonella Detection" Foods 14, no. 11: 1892. https://doi.org/10.3390/foods14111892
APA StyleShi, C., Tan, H., Yu, Z., Li, W., Man, Y., & Zhang, Q. (2025). CRISPR-Cas12a/Aurora Deoxyribozyme Cascade: A Label-Free Ultrasensitive Platform for Rapid Salmonella Detection. Foods, 14(11), 1892. https://doi.org/10.3390/foods14111892