

Variation in Intramuscular Fat Deposition of Goats and Sheep and Its Correlation with Gut Microbiota
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sample Collection
2.3. Determination of IMF Content, Concentrations of Blood Lipid, Enzyme Levels, and Fatty Acid Content
2.4. SrRNA Analysis
2.4.1. Microbial DNA Extractions and PCR Amplification
2.4.2. Library Construction and Sequencing
2.4.3. Processing of Sequencing Data
2.4.4. Alpha- and Beta-Diversity Analyses
2.4.5. Functional Prediction of the Microbial Genes
2.5. Statistical Analysis
3. Results
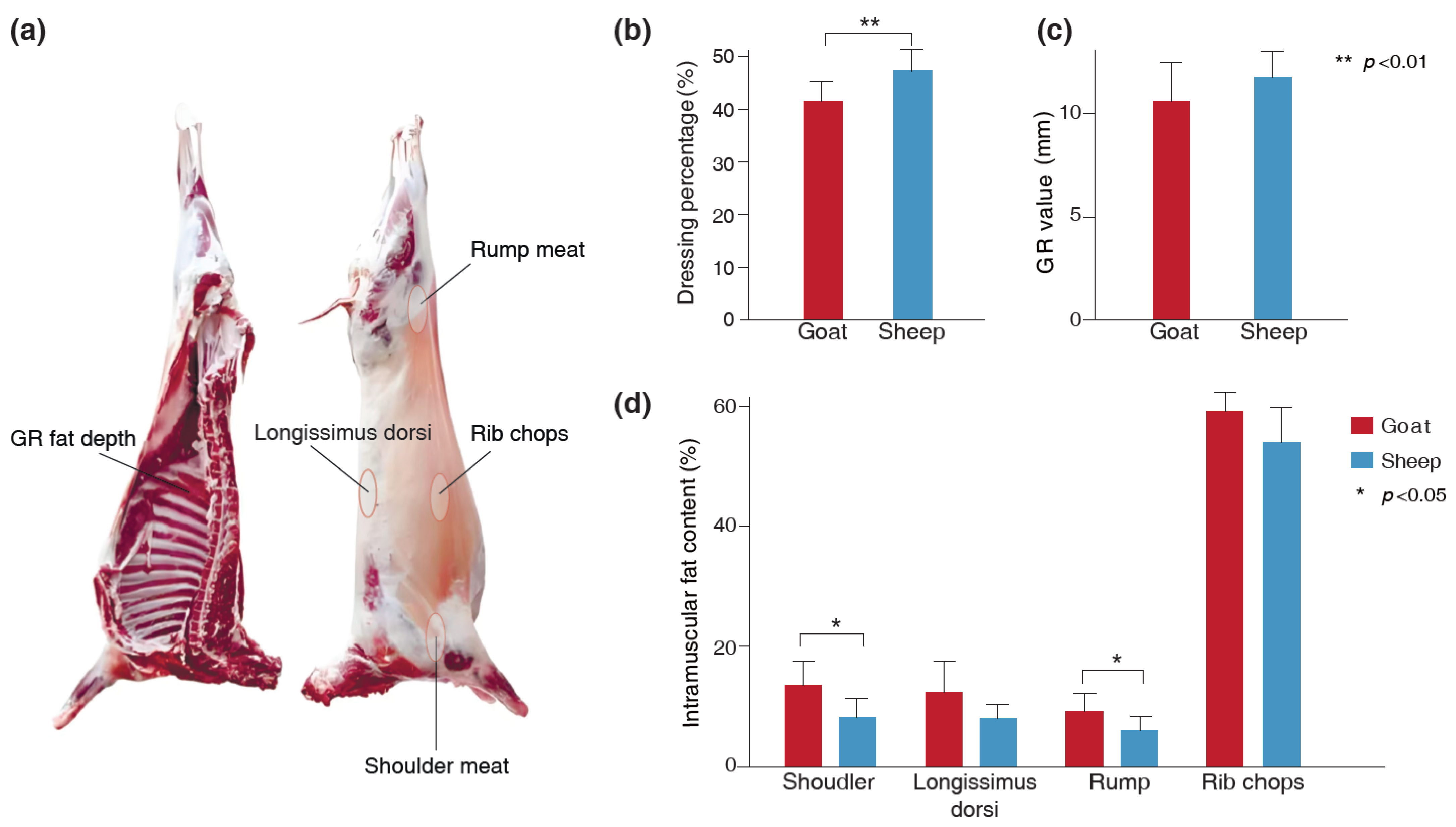
3.1. The Differences in Dressing Percentage, GR Value, and IMF Content of the Goats and Sheep
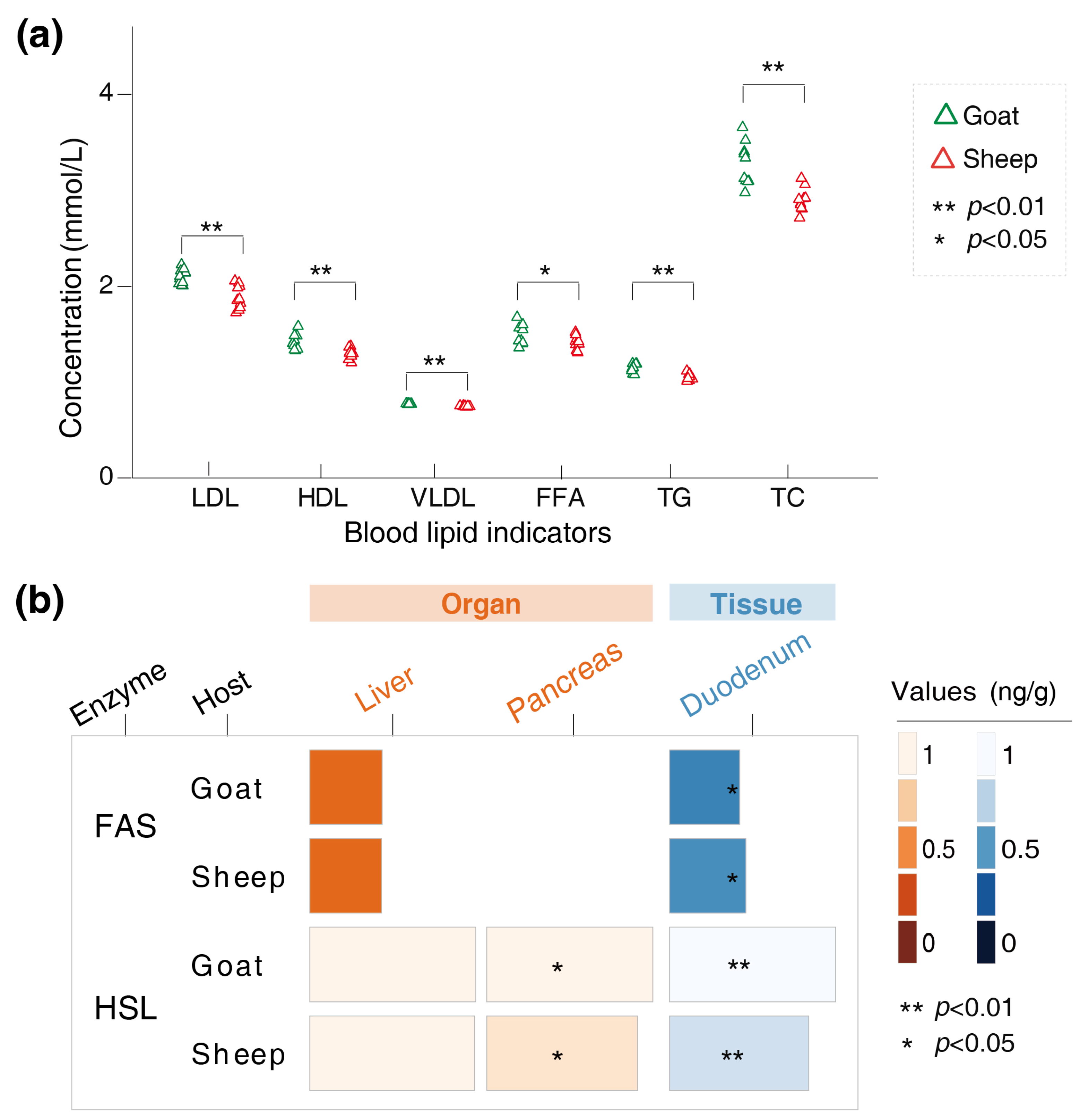
3.2. Variations in the Physiological Indicators of the Goats and Sheep
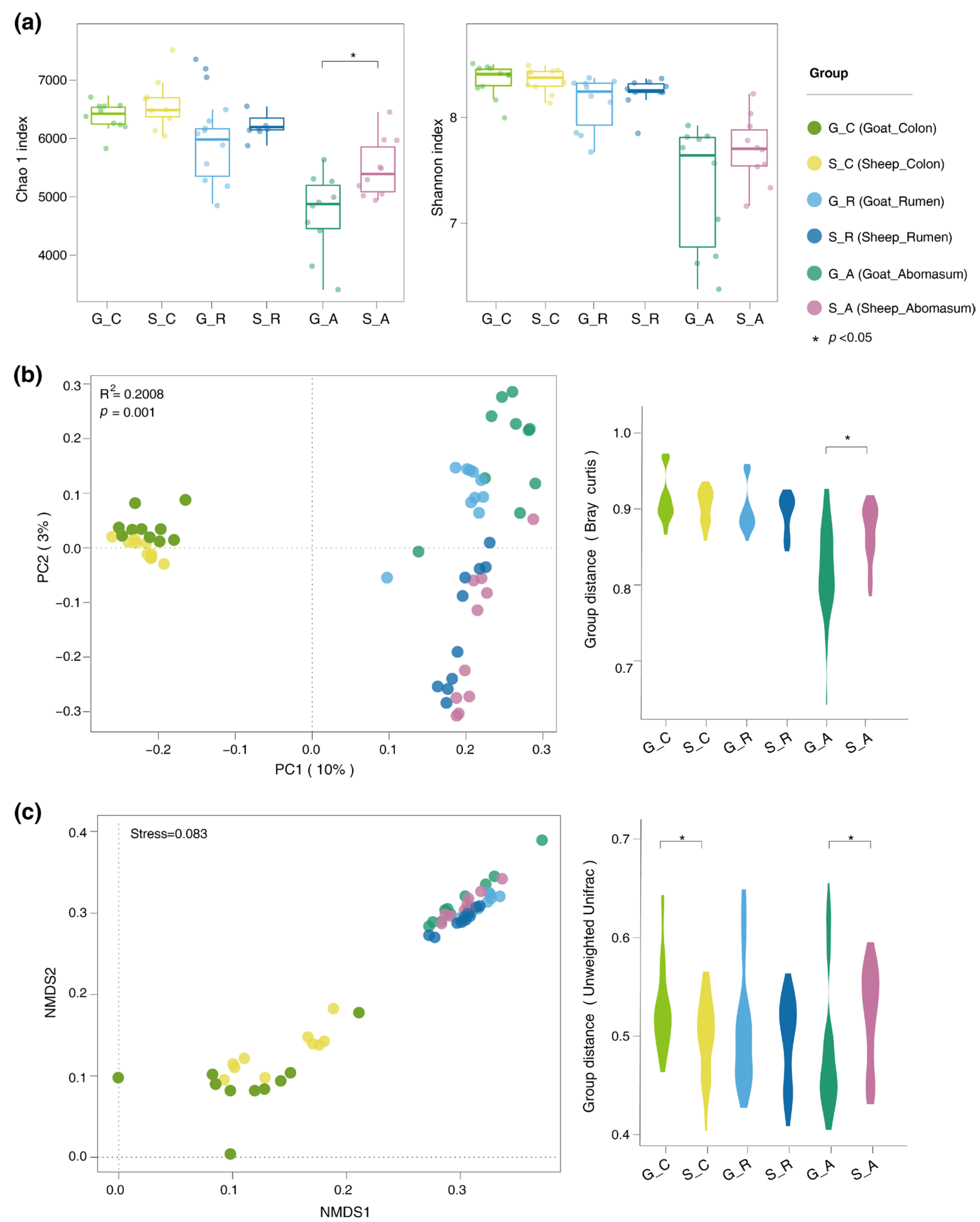
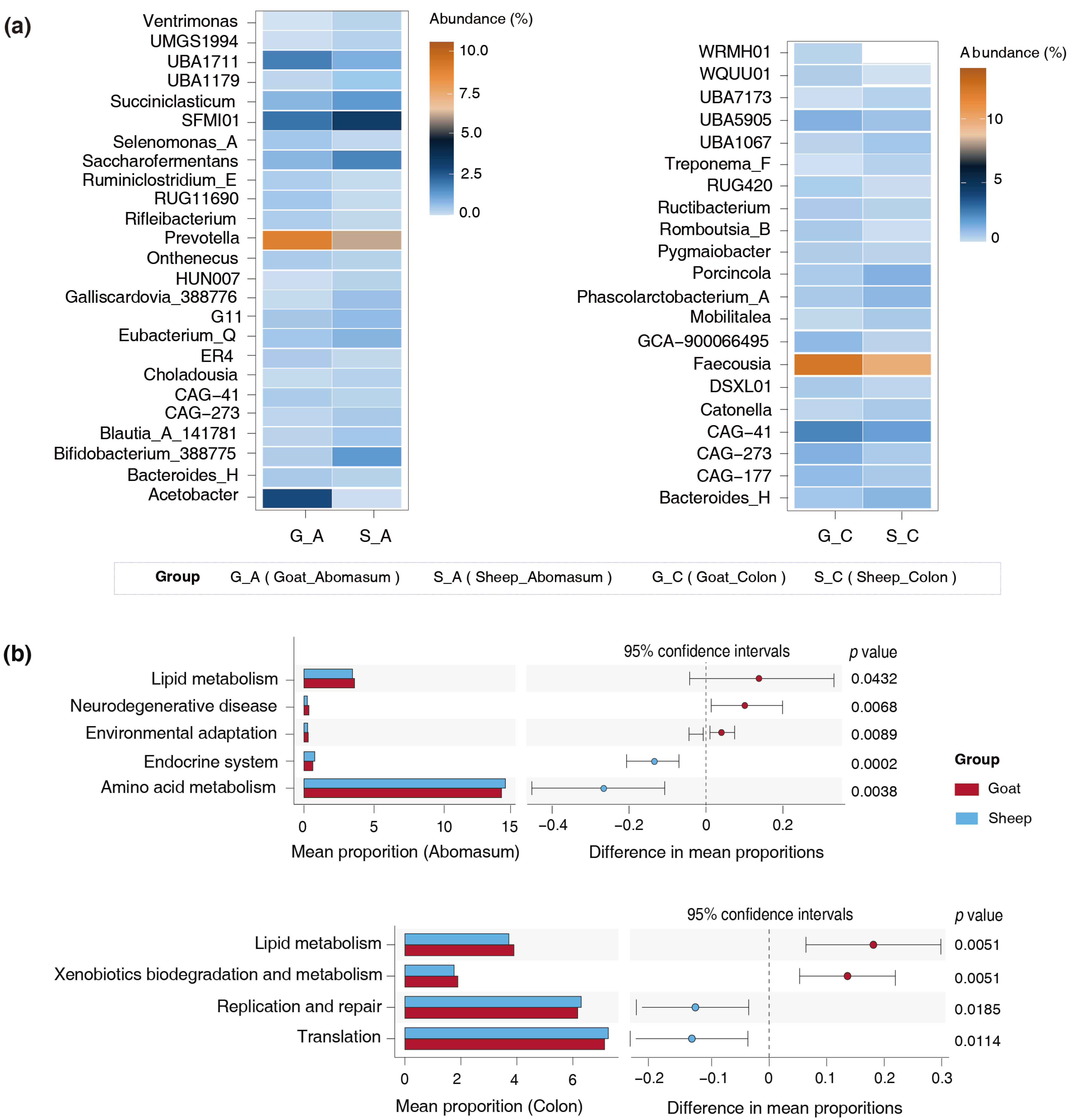
3.3. Differences in the Gut Microbiota of the Goats and Sheep
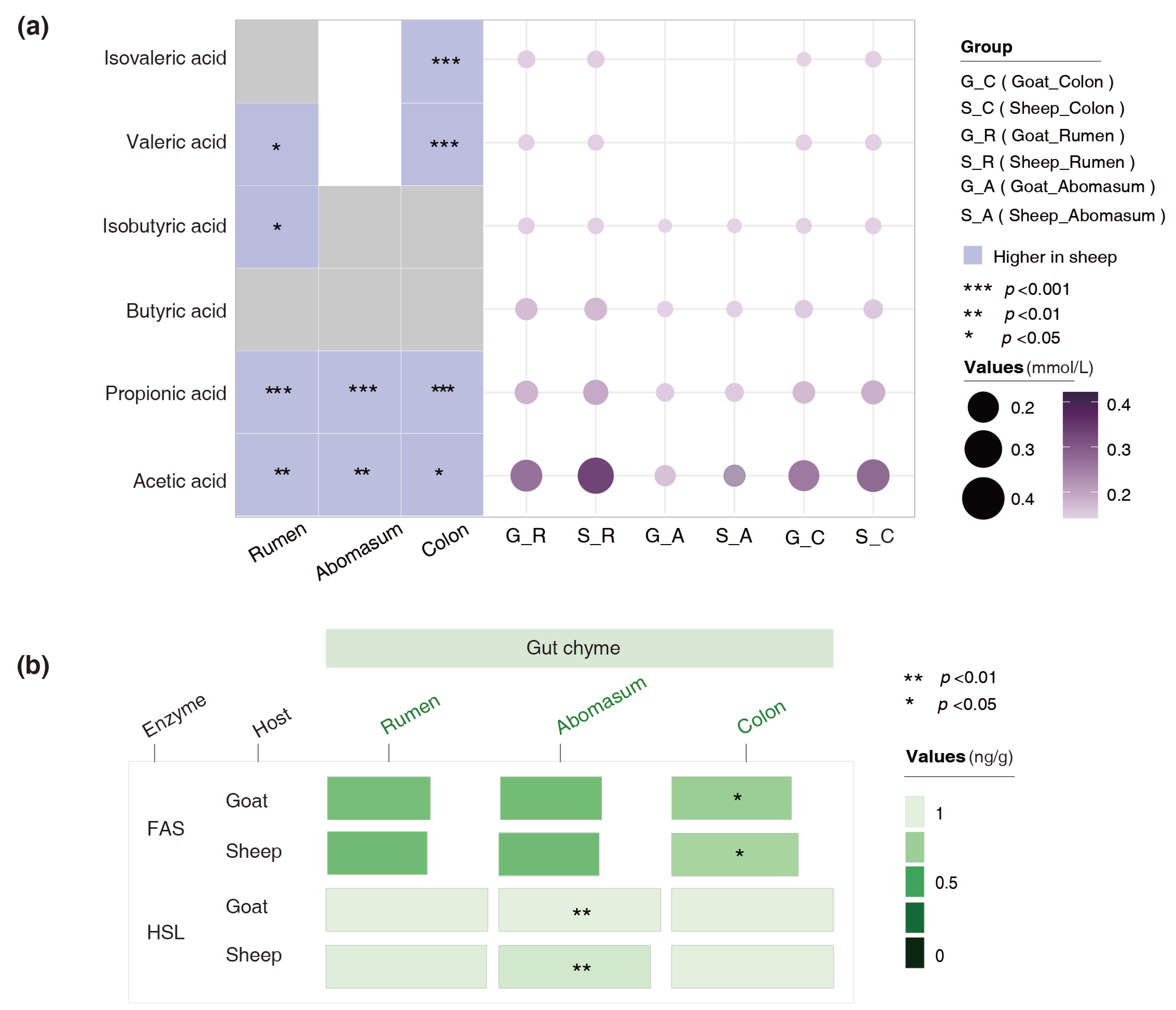
3.4. Variations in the Concentrations of SCFAs and Lipid-Metabolizing Enzymes in the Gut Chyme of the Goats and Sheep
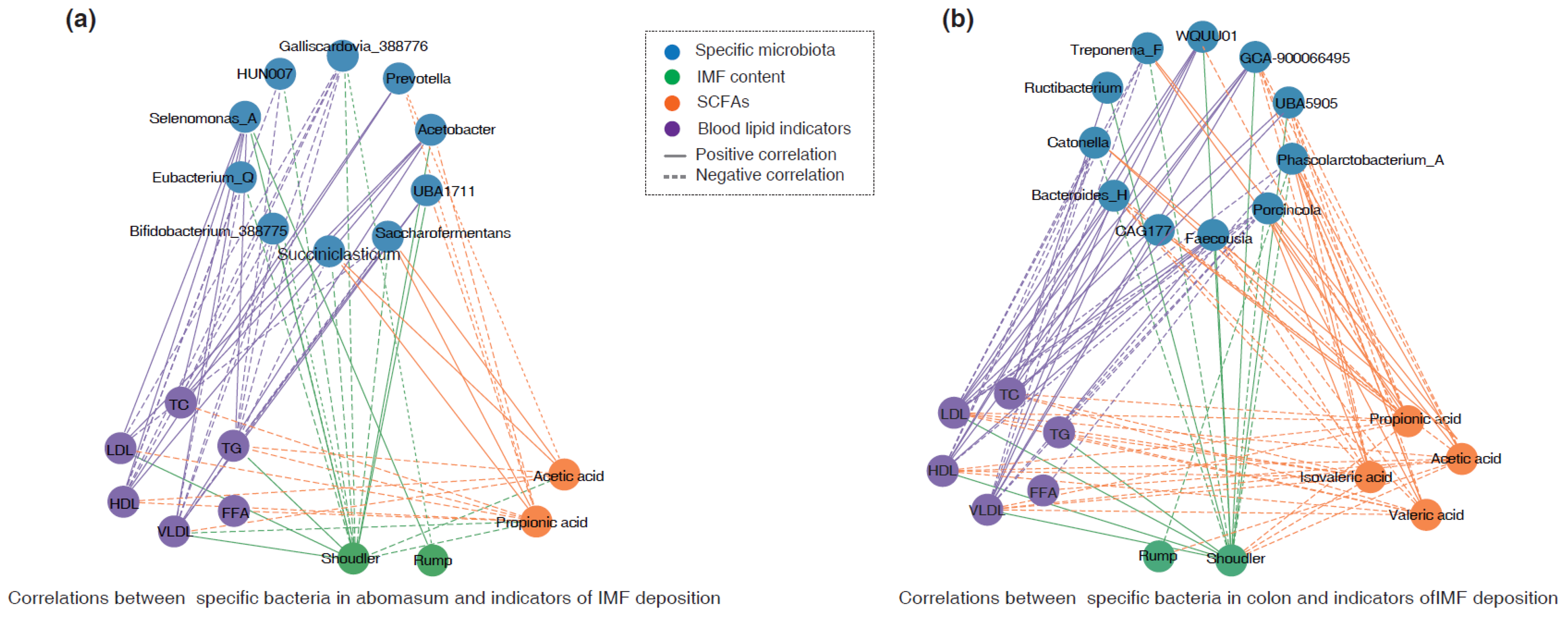
3.5. Association of IMF Deposition with Identified Bacteria and SCFAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. World Agriculture Statistics. 2024. Available online: http://www.fao.org/statistics (accessed on 19 May 2025).
- Kruk, Z.A.; Bottema, M.J.; Reyes-Veliz, L.; Forder, R.E.A.; Pitchford, W.S.; Bottema, C.D.K. Vitamin A and marbling attributes: Intramuscular fat hyperplasia effects in cattle. Meat Sci. 2018, 137, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Troy, D.J.; Tiwari, B.K.; Joo, S.T. Health Implications of Beef Intramuscular Fat Consumption. Korean J. Food Sci. Anim. Resour. 2016, 36, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality—A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, C.; Kong, Y.; Li, F.; Yue, X. Effects of intramuscular fat on meat quality and its regulation mechanism in Tan sheep. Front. Nutr. 2022, 9, 908355. [Google Scholar] [CrossRef]
- Cui, H.; Liu, L.; Liu, X.; Wang, Y.; Luo, N.; Tan, X.; Zhu, Y.; Liu, R.; Zhao, G.; Wen, J. A selected population study reveals the biochemical mechanism of intramuscular fat deposition in chicken meat. J. Anim. Sci. Biotechnol. 2022, 13, 54. [Google Scholar] [CrossRef]
- Wang, L.; Liu, S.; Zhang, S.; Wang, Y.; Zhou, Y.; Shan, T. Single-nucleus transcriptomics reveal the cytological mechanism of conjugated linoleic acids in regulating intramuscular fat deposition. Elife 2025, 13, RP99790. [Google Scholar] [CrossRef]
- Sun, J.; Xie, F.; Wang, J.; Luo, J.; Chen, T.; Jiang, Q.; Xi, Q.; Liu, G.E.; Zhang, Y. Integrated meta-omics reveals the regulatory landscape involved in lipid metabolism between pig breeds. Microbiome 2024, 12, 33. [Google Scholar] [CrossRef]
- Zeng, Y.; Mou, H.; He, Y.; Zhang, D.; Pan, X.; Zhou, L.; Shen, Y.; E, G. Effects of Key Rumen Bacteria and Microbial Metabolites on Fatty Acid Deposition in Goat Muscle. Animals 2024, 14, 22. [Google Scholar] [CrossRef]
- Zhao, X.; Ding, X.; Yang, Z.; Shen, Y.; Zhang, S.; Shi, S. Effects of Clostridium butyricum on thigh muscle lipid metabolism of broilers. Ital. J. Anim. Sci. 2018, 17, 1010–1020. [Google Scholar] [CrossRef]
- Zhang, T.; Ding, H.; Chen, L.; Lin, Y.; Gong, Y.; Pan, Z.; Zhang, G.; Xie, K.; Dai, G.; Wang, J. Antibiotic-Induced Dysbiosis of Microbiota Promotes Chicken Lipogenesis by Altering Metabolomics in the Cecum. Metabolites 2021, 11, 8. [Google Scholar] [CrossRef]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, Y.; Zuo, S.; Peng, S.; Wang, Z.; Zhang, Y.; Luo, H. Untargeted and Targeted Metabolomics Profiling of Muscle Reveals Enhanced Meat Quality in Artificial Pasture Grazing Tan Lambs via Rescheduling the Rumen Bacterial Community. J. Agric. Food Chem. 2021, 69, 846–858. [Google Scholar] [CrossRef] [PubMed]
- De Smet, S.; Vossen, E. Meat: The balance between nutrition and health—A review. Meat Sci. 2016, 120, 145–156. [Google Scholar] [CrossRef]
- Babour, A.; Wei, J.; Luoa, Y. Fatty Acid Composition of Muscles Tissues of Longdong Goat (Black and White Cashmere). J. Chromatog. Separat. Tech. 2018, 9, 2. [Google Scholar]
- Tangerman, A.; Nagengast, F.M. A gas chromatographic analysis of fecal short-chain fatty acids, using the direct injection method. Analyt. Biochem. 1996, 236, 1–8. [Google Scholar] [CrossRef]
- Choi, Y.; Jeong, J.; Kim, M.; Cha, S.; Han, K. Genomics, Backtracking identification techniques for predicting unclear bacterial taxonomy at species level: Molecular diagnosis-based bacterial classification. Genes 2025, 47, 503–508. [Google Scholar]
- Lin, H.; Lin, J.; Pan, T.; Li, T.; Jiang, H.; Fang, Y.; Wang, Y.; Wu, F.; Huang, J.; Zhang, H.; et al. Polymeric immunoglobulin receptor deficiency exacerbates autoimmune hepatitis by inducing intestinal dysbiosis and barrier dysfunction. Cell Death Dis. 2023, 14, 68. [Google Scholar] [CrossRef]
- Prasad, D.V.; Madhusudanan, S.; Jaganathan, S. uCLUST-a new algorithm for clustering unstructured data. ARPN J. Eng. Appl. Sci. 2015, 10, 2108–2117. [Google Scholar]
- McDonald, D.; Jiang, Y.; Balaban, M.; Cantrell, K.; Zhu, Q.; Gonzalez, A.; Morton, J.T.; Nicolaou, G.; Parks, D.H.; Karst, S.M.; et al. Greengenes2 unifies microbial data in a single reference tree. Nat. Biotechnol. 2024, 42, 715–718. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M. PICRUSt2 for prediction of metagenome functions. Nat. Biotech. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Env. Micro 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Thakur, N.; Mehrotra, D.; Bansal, A.; Bala, M.N.; Mehrotra, D.; Bansal, A.; Bala, M. Analysis and Implementation of the Bray–Curtis Distance-Based Similarity Measure for Retrieving Information from the Medical Repository. In Proceedings of the International Conference on Innovative Computing and Communications, Ostrava, Czech Republic, 21–22 March 2019; Lecture Notes in Networks and Systems. pp. 117–125. [Google Scholar]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.R.; Santra, A.; Karim, S.A. Carcass yield, composition and meat quality attributes of sheep and goat under semiarid conditions. Meat Sci. 2004, 66, 757–763. [Google Scholar] [CrossRef]
- Mazinani, M.; Rude, B. Population, world production and quality of sheep and goat products. Am. J. Anim. Vet. Sci. 2020, 15, 291–299. [Google Scholar] [CrossRef]
- Casey, N.H. Carcass and Growth Characteristics of Four South African Sheep Breeds and the Boer Goat. Ph.D. Thesis, Universiteit van Pretoria, Pretoria, South Africa, 1983. [Google Scholar]
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animals 2010, 4, 303–319. [Google Scholar] [CrossRef]
- Yan, J.; Liao, K.; Wang, T.; Mai, K.; Xu, W.; Ai, Q. Dietary Lipid Levels Influence Lipid Deposition in the Liver of Large Yellow Croaker (Larimichthys crocea) by Regulating Lipoprotein Receptors, Fatty Acid Uptake and Triacylglycerol Synthesis and Catabolism at the Transcriptional Level. PLoS ONE 2015, 10, e0129937. [Google Scholar] [CrossRef]
- Ge, K.; Ye, P.; Yang, L.; Kuang, J.; Chen, X.; Geng, Z. Comparison of slaughter performance, meat traits, serum lipid parameters and fat tissue between Chaohu ducks with high-and low-intramuscular fat content. Anim. Biotech. 2020, 31, 245–255. [Google Scholar] [CrossRef]
- Hu, R.; Zou, H.; Wang, H.; Wang, Z.; Wang, X.; Ma, J.; Shah, A.M.; Peng, Q.; Xue, B.; Wang, L.; et al. Dietary Energy Levels Affect Rumen Bacterial Populations that Influence the Intramuscular Fat Fatty Acids of Fattening Yaks (Bos grunniens). Animals 2020, 10, 1474. [Google Scholar] [CrossRef]
- Mabrouk, G.M.; Helmy, I.M.; Thampy, K.G.; Wakil, S.J. Acute hormonal control of acetyl-CoA carboxylase. The roles of insulin, glucagon, and epinephrine. J. Biol. Chem. 1990, 265, 6330–6338. [Google Scholar] [CrossRef]
- Menendez, J.A.; Lupu, R. Fatty acid synthase-catalyzed de novo fatty acid biosynthesis: From anabolic-energy-storage pathway in normal tissues to jack-of-all-trades in cancer cells. Arch. Immunol. Therap. Exp. 2004, 52, 414–426. [Google Scholar]
- Lowe, M.E. The triglyceride lipases of the pancreas. J. Lipid Res. 2002, 43, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Wen, H.; Zeng, L.B.; Jiang, M.; Wu, F.; Liu, W.; Yang, C.G. Changes in the activities and mRNA expression levels of lipoprotein lipase (LPL), hormone-sensitive lipase (HSL) and fatty acid synthetase (FAS) of Nile tilapia (Oreochromis niloticus) during fasting and re-feeding. Aquaculture 2013, 400, 29–35. [Google Scholar] [CrossRef]
- Den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARγ-Dependent Switch from Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef]
- Li, X.; Chen, H.; Guan, Y.; Li, X.; Lei, L.; Liu, J.; Yin, L.; Liu, G.; Wang, Z. Acetic acid activates the AMP-activated protein kinase signaling pathway to regulate lipid metabolism in bovine hepatocytes. PLoS ONE 2013, 8, e67880. [Google Scholar] [CrossRef]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.; Forrest, G.; et al. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS ONE 2012, 7, e35240. [Google Scholar] [CrossRef]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Li, C.; Tian, H.; Weng, X.; Lin, C.; Zhang, D.; Zhao, Y.; Li, X.; Cheng, J. Microbiomes, Rumen microbiome and fat deposition in sheep: Insights from a bidirectional mendelian randomization study. Npj Biofilms Microbiomes 2024, 10, 129. [Google Scholar] [CrossRef]
- Yin, Y.N.; Yu, Q.F.; Fu, N.; Liu, X.W.; Lu, F.G. Effects of four Bifidobacteria on obesity in high-fat diet induced rats. World J. Gastro. 2010, 16, 3394–3401. [Google Scholar] [CrossRef]
- Kim, G.; Yoon, Y.; Park, J.H.; Park, J.W.; Noh, M.G.; Kim, H.; Park, C.; Kwon, H.; Park, J.H.; Kim, Y.; et al. Bifidobacterial carbohydrate/nucleoside metabolism enhances oxidative phosphorylation in white adipose tissue to protect against diet-induced obesity. Microbiome 2022, 10, 188. [Google Scholar] [CrossRef]
- Begley, M.; Hill, C.; Gahan, C.G. Bile salt hydrolase activity in probiotics. Appl. Environ. Microbiol. 2006, 72, 1729–1738. [Google Scholar] [CrossRef]
- Maruta, H.; Fujii, Y.; Toyokawa, N.; Nakamura, S.; Yamashita, H. Effects of Bifidobacterium-Fermented Milk on Obesity: Improved Lipid Metabolism through Suppression of Lipogenesis and Enhanced Muscle Metabolism. Int. J. Mol. Sci. 2024, 25, 9934. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xia, Q.; Wu, T.; Shao, Y.; Wang, Y.; Jin, N.; Tian, P.; Wu, L.; Lu, X. Prophylactic treatment with Bacteroides uniformis and Bifidobacterium bifidum counteracts hepatic NK cell immune tolerance in nonalcoholic steatohepatitis induced by high fat diet. Gut Microbes 2024, 16, 2302065. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.D. Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut health. Gut Microbes 2020, 12, 1802866. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Li, Y.; Wang, X.; Yu, M.; Liu, X.; Zhang, H.; Meng, Q.; Majeed, U.; Jian, L.; Song, W.; et al. Positive interactions among Corynebacterium glutamicum and keystone bacteria producing SCFAs benefited T2D mice to rebuild gut eubiosis. Food Res. Int. 2023, 172, 113163. [Google Scholar] [CrossRef]
- Li, W.-J.; Tao, M.; Zhang, N.-F.; Deng, K.-D.; Diao, Q.-Y. Dietary fat supplement affected energy and nitrogen metabolism efficiency and shifted rumen fermentation toward glucogenic propionate production via enrichment of Succiniclasticum in male twin lambs. J. Integ. Agric. 2023, 24, 1285–1295. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Zhang, X.X.; Li, F.D.; Li, C.; Li, G.Z.; Zhang, D.Y.; Song, Q.Z.; Li, X.L.; Zhao, Y.; Wang, W.M. Characterization of the rumen microbiota and its relationship with residual feed intake in sheep. Animals 2021, 15, 100161. [Google Scholar] [CrossRef]
- van Gylswyk, N.O. Succiniclasticum ruminis gen. nov., sp. nov., a ruminal bacterium converting succinate to propionate as the sole energy-yielding mechanism. Int. J. System. Bacteriol. 1995, 45, 297–300. [Google Scholar] [CrossRef]
- Wang, B.; Luo, Y.; Wang, Y.; Wang, D.; Hou, Y.; Yao, D.; Tian, J.; Jin, Y. Rumen bacteria and meat fatty acid composition of Sunit sheep reared under different feeding regimens in China. J. Sci. Food Agric. 2021, 101, 1100–1110. [Google Scholar] [CrossRef]
- Zhou, G.; Liang, X.; He, X.; Li, J.; Tian, G.; Liu, Y.; Wang, X.; Chen, Y.; Yang, Y. Compound enzyme preparation supplementation improves the production performance of goats by regulating rumen microbiota. Appl. Microbiol. Biotechnol. 2023, 107, 7287–7299. [Google Scholar] [CrossRef]
- Liu, H.; Peng, W.; Mao, K.; Yang, Y.; Wu, Q.; Wang, K.; Zeng, M.; Han, X.; Han, J.; Zhou, H. The Changes in Fecal Bacterial Communities in Goats Offered Rumen-Protected Fat. Microorganisms 2024, 12, 822. [Google Scholar] [CrossRef]
- Espiritu, H.M.; Valete, E.J.P.; Mamuad, L.L.; Jung, M.; Paik, M.J.; Lee, S.S.; Cho, Y.I. Metabolic Footprint of Treponema phagedenis and Treponema pedis Reveals Potential Interaction Towards Community Succession and Pathogenesis in Bovine Digital Dermatitis. Pathogens 2024, 13, 796. [Google Scholar] [CrossRef] [PubMed]
- Jiao, A.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Luo, Y.; Luo, J.; Mao, X.; Chen, D. Short chain fatty acids could prevent fat deposition in pigs via regulating related hormones and genes. Food Funct. 2020, 11, 1845–1855. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Li, S.; Hou, J.; Tang, Z.; Shi, B.; Luo, Y.; Wang, J.; Zhao, F. Variation in Intramuscular Fat Deposition of Goats and Sheep and Its Correlation with Gut Microbiota. Foods 2025, 14, 1885. https://doi.org/10.3390/foods14111885
Yang L, Li S, Hou J, Tang Z, Shi B, Luo Y, Wang J, Zhao F. Variation in Intramuscular Fat Deposition of Goats and Sheep and Its Correlation with Gut Microbiota. Foods. 2025; 14(11):1885. https://doi.org/10.3390/foods14111885
Chicago/Turabian StyleYang, Lei, Shaobin Li, Jiagong Hou, Zhisheng Tang, Bingang Shi, Yuzhu Luo, Jiqing Wang, and Fangfang Zhao. 2025. "Variation in Intramuscular Fat Deposition of Goats and Sheep and Its Correlation with Gut Microbiota" Foods 14, no. 11: 1885. https://doi.org/10.3390/foods14111885
APA StyleYang, L., Li, S., Hou, J., Tang, Z., Shi, B., Luo, Y., Wang, J., & Zhao, F. (2025). Variation in Intramuscular Fat Deposition of Goats and Sheep and Its Correlation with Gut Microbiota. Foods, 14(11), 1885. https://doi.org/10.3390/foods14111885