
Improving the Efficiency of Viability-qPCR with Lactic Acid Enhancer for the Selective Detection of Live Pathogens in Foods
 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Conditions
2.2. Testing the Sublethal Effect of Lactic Acid on the Strain E. coli ATCC 8739
2.3. Preparation of E. coli Suspensions and Heat Treatment
2.4. Pre-Treatment of Cell Suspensions with Lactic Acid
2.5. PMA Treatment and Cross-Linking
2.6. Extraction of DNA and qPCR
2.7. Artificially Inoculated Food Assays
2.8. Statistical Analysis
3. Results
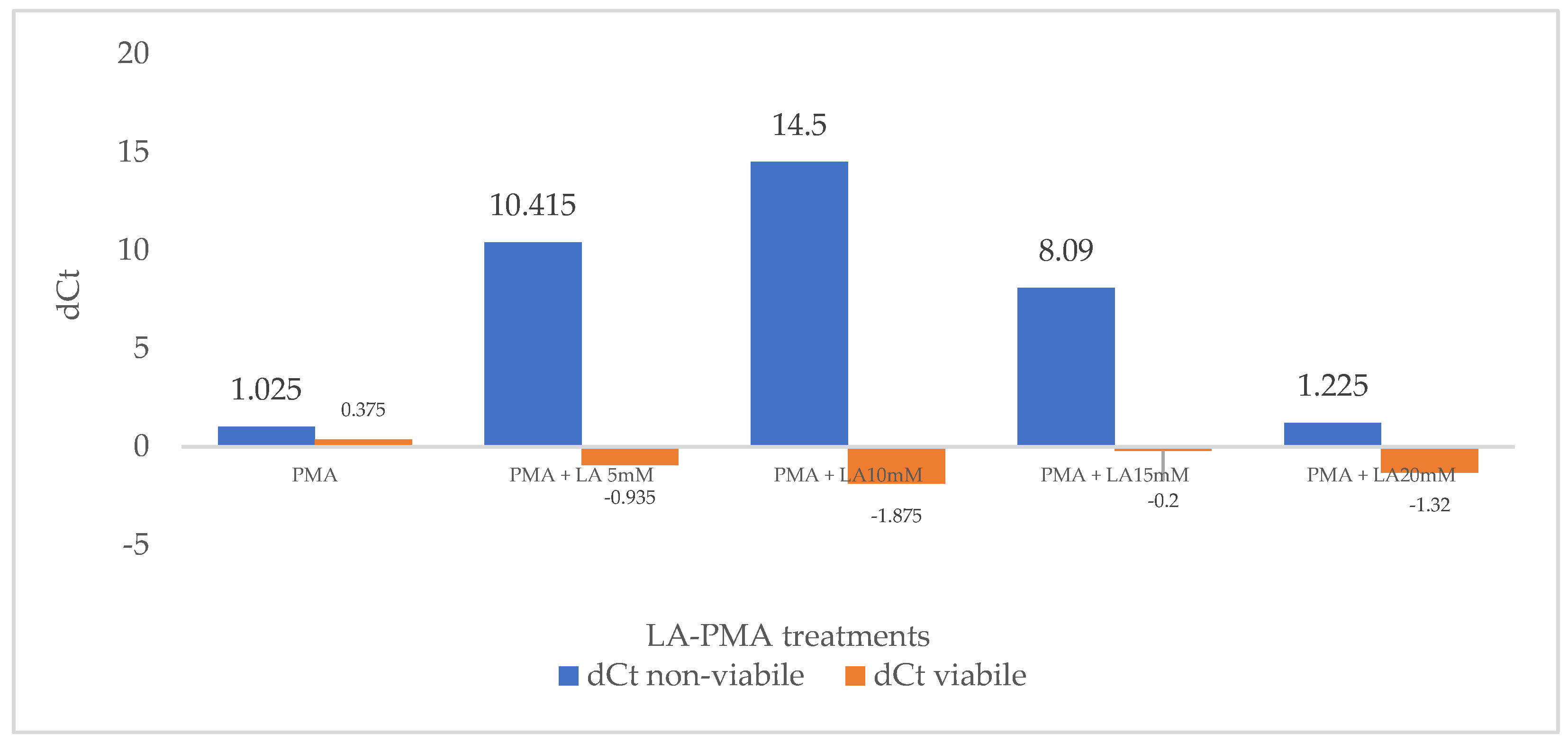
3.1. Effect of LA and PMA Treatments on qPCR Amplification from Viable and Non-Viable Bacterial Cell Suspensions
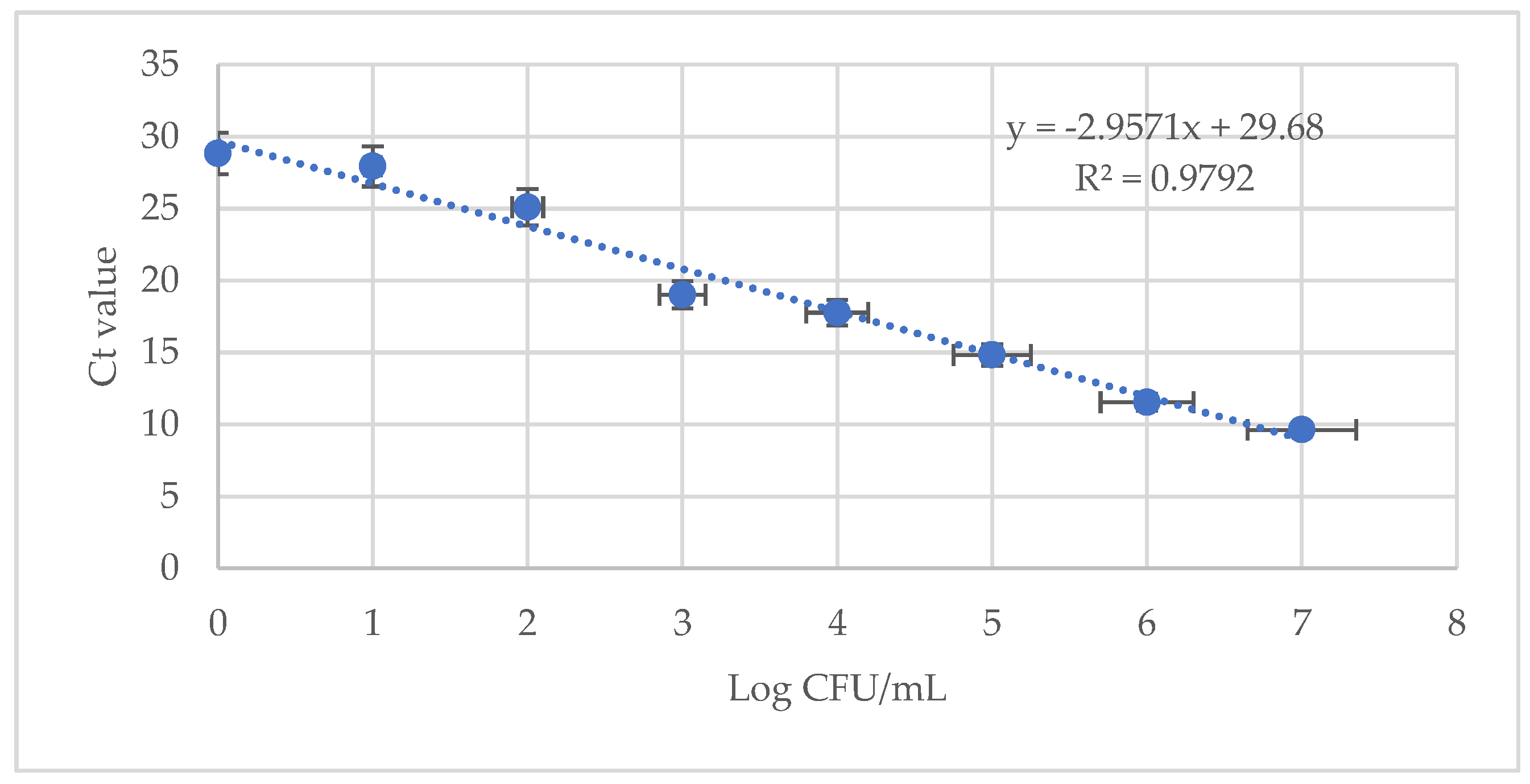
3.2. Limit of Detection for LA-PMA qPCR Assay in a Background of Dead Bacteria
3.3. Detection of E. coli ATCC 8739 via LA-PMA-qPCR Assay in Artificially Inoculated Milk
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pires, S.M.; Desta, B.N.; Mughini-Gras, L.; Mmbaga, B.T.; Fayemi, O.E.; Salvador, E.M.; Gobena, T.; Majowicz, S.E.; Hald, T.; Hoejskov, P.S.; et al. Burden of foodborne diseases: Think global, act local. Curr. Opin. Food Sci. 2021, 39, 152–159. [Google Scholar] [CrossRef]
- Gourama, H. Foodborne pathogens. In Food Safety Engineering; Springer: Berlin/Heidelberg, Germany, 2020; pp. 25–49. [Google Scholar]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The population genetics of pathogenic Escherichia coli. Nat. Rev. Microbiol. 2021, 19, 37–54. [Google Scholar] [CrossRef] [PubMed]
- Rani, A.; Ravindran, V.B.; Surapaneni, A.; Mantri, N.; Ball, A.S. Trends in point-of-care diagnosis for Escherichia coli O157: H7 in food and water. Int. J. Food Microbiol. 2021, 349, 109233. [Google Scholar] [CrossRef] [PubMed]
- Singha, S.; Thomas, R.; Viswakarma, J.N.; Gupta, V.K. Foodborne illnesses of Escherichia coli O157origin and its control measures. J. Food Sci. Technol. 2023, 60, 1274–1283. [Google Scholar] [CrossRef]
- Nocker, A.; Cheung, C.Y.; Camper, A.K. Comparison of propidium monoazide with ethidium monoazide for differentiation of live vs. dead bacteria by selective removal of DNA from dead cells. J. Microbiol. Methods 2006, 67, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Nocker, A.; Sossa-Fernandez, P.; Burr, M.D.; Camper, A.K. Use of propidium monoazide for live/dead distinction in microbial ecology. Appl. Environ. Microbiol. 2007, 73, 5111–5117. [Google Scholar] [CrossRef] [PubMed]
- Dinu, L.D.; Bach, S. Detection of viable but non-culturable Escherichia coli O157: H7 from vegetable samples using quantitative PCR with propidium monoazide and immunological assays. Food Control 2013, 31, 268–273. [Google Scholar] [CrossRef]
- Rudi, K.; Moen, B.; Drømtorp, S.M.; Holck, A.L. Use of ethidium monoazide and PCR in combination for quantification of viable and dead cells in complex samples. Appl. Environ. Microbiol. 2005, 71, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Elizaquível, P.; Sánchez, G.; Aznar, R. Application of propidium monoazide quantitative PCR for selective detection of live Escherichia coli O157: H7 in vegetables after inactivation by essential oils. Int. J. Food Microbiol. 2012, 159, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, X.; Fan, X.; Xu, J.; Liu, Q.; Wu, Z.; Pan, D. PMA-qPCR method for the selective quantitation of viable lactic acid bacteria in fermented milk. Front. Microbiol. 2022, 13, 984506. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, W.; Zhao, H.; Luo, Y.; Wan, M.; Li, Y. A new PMA-qPCR method for rapid and accurate detection of viable bacteria and spores of marine-derived Bacillus velezensis B-9987. J. Microbiol. Methods 2022, 199, 106537. [Google Scholar] [CrossRef] [PubMed]
- Okada, A.; Tsuchida, M.; Rahman, M.M.; Inoshima, Y. Two-Round Treatment with Propidium Monoazide Completely Inhibits the Detection of Dead Campylobacter Spp. Cells by Quantitative PCR. Front. Microbiol. 2022, 13, 801961. [Google Scholar] [CrossRef] [PubMed]
- Infante, N.; Rodríguez, R.; Bartolo, Y.; Sánchez, O.; Sanz, I.; Bermeo, L.; Morató, J. Biofunctionalization of Cork with Moringa oleifera Seeds and Use of PMA Staining and qPCR to Detect Viability of Escherichia coli. Water 2021, 13, 2731. [Google Scholar] [CrossRef]
- Rey, M.D.; Cap, M.; Favre, L.C.; Rodríguez Racca, A.; Dus Santos, M.J.; Vaudagna, S.R.; Mozgovoj, M. Evaluation of PMA-qPCR methodology to detect and quantify viable Shiga toxin-producing Escherichia coli in beef burgers. J. Food Process. Preserv. 2021, 45, e15338. [Google Scholar] [CrossRef]
- Do, T.N.; Lao, T.D.; Le, T.A. Establishment of PMA Real-Time PCR Method to Detect Viable Cells of Listeria monocytogenes and Salmonella spp. in Milk and Dairy Products. Asian J. Pharm. Res. Health Care 2021, 20, 146–156. [Google Scholar]
- Truchado, P.; Gil, M.I.; Kostic, T.; Allende, A. Optimization and validation of a PMA qPCR method for Escherichia coli quantification in primary production. Food Control 2016, 62, 150–156. [Google Scholar] [CrossRef]
- Zeng, D.; Chen, Z.; Jiang, Y.; Xue, F.; Li, B. Advances and challenges in viability detection of foodborne pathogens. Front. Microbiol. 2016, 7, 1833. [Google Scholar] [CrossRef] [PubMed]
- Van Holm, W.; Ghesquière, J.; Boon, N.; Verspecht, T.; Bernaerts, K.; Zayed, N.; Chatzigiannidou, I.; Teughels, W. A viability quantitative PCR dilemma: Are longer amplicons better? Appl. Environ. Microbiol. 2021, 87, e02653-20. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.L.; Levin, R.E. Discrimination of viable and dead Vibrio vulnificus after refrigerated and frozen storage using EMA, sodium deoxycholate and real-time PCR. J. Microbiol. Methods 2009, 79, 184–188. [Google Scholar] [CrossRef]
- Yang, X.; Badoni, M.; Gill, C.O. Use of propidium monoazide and quantitative PCR for differentiation of viable Escherichia coli from E. coli killed by mild or pasteurizing heat treatments. Food Microbiol. 2011, 28, 1478–1482. [Google Scholar] [CrossRef]
- Nkuipou-Kenfack, E.; Engel, H.; Fakih, S.; Nocker, A. Improving efficiency of viability-PCR for selective detection of live cells. J. Microbiol. Methods 2013, 93, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Liang, T.; Zhan, Z.; Liu, R.; Li, F.; Xu, H. Rapid and simultaneous quantification of viable Escherichia coli O157: H7 and Salmonella spp. in milk through multiplex real-time PCR. J. Dairy Sci. 2017, 100, 8804–8813. [Google Scholar] [CrossRef] [PubMed]
- Canh, V.D.; Kasuga, I.; Furumai, H.; Katayama, H. Viability RT-qPCR combined with sodium deoxycholate pre-treatment for selective quantification of infectious viruses in drinking water samples. Food Environ. Virol. 2019, 11, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ye, C.; Xu, H.; Aguilar, Z.P.; Xiong, Y.; Lai, W.; Wei, H. Development of an SD-PMA-mPCR assay with internal amplification control for rapid and sensitive detection of viable Salmonella spp., Shigella spp. and Staphylococcus aureus in food products. Food Control 2015, 57, 314–320. [Google Scholar] [CrossRef]
- Dong, L.; Liu, H.; Meng, L.; Xing, M.; Wang, J.; Wang, C.; Chen, H.; Zheng, N. Quantitative PCR coupled with sodium dodecyl sulfate and propidium monoazide for detection of viable Staphylococcus aureus in milk. J. Dairy Sci. 2018, 101, 4936–4943. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Liu, H.; Meng, L.; Xing, M.; Lan, T.; Gu, M.; Zheng, N.; Wang, C.; Chen, H.; Wang, J. Quantitative PCR coupled with sodium dodecyl sulfate and propidium monoazide for detection of culturable Escherichia coli in milk. J. Dairy Sci. 2019, 102, 6914–6919. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Gao, Y.; Miya, S.; Kuda, T.; Kimura, B. Discrimination of live and dead cells of Escherichia coli using propidium monoazide after sodium dodecyl sulfate treatment. Food Control 2017, 71, 79–82. [Google Scholar] [CrossRef]
- Wang, H.; Gill, C.O.; Yang, X. Use of sodium lauroyl sarcosinate (sarkosyl) in viable real-time PCR for enumeration of Escherichia coli. J. Microbiol. Methods 2014, 98, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Xiong, J.; Nyaruaba, R.; Li, J.; Muturi, E.; Liu, H.; Yu, J.; Yang, H.; Wei, H. Rapid determination of infectious SARS-CoV-2 in PCR-positive samples by SDS-PMA assisted RT-qPCR. Sci. Total Environ. 2021, 797, 149085. [Google Scholar] [CrossRef] [PubMed]
- Zi, C.; Zeng, D.; Ling, N.; Dai, J.; Xue, F.; Jiang, Y.; Li, B. An improved assay for rapid detection of viable Staphylococcus aureus cells by incorporating surfactant and PMA treatments in qPCR. BMC Microbiol. 2018, 18, 132. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xin, H.; Li, S.F. Multiplex PMA–qPCR Assay with internal amplification control for simultaneous detection of viable Legionella pneumophila, Salmonella typhimurium, and Staphylococcus aureus in environmental waters. Environ. Sci. Technol. 2015, 49, 14249–14256. [Google Scholar] [CrossRef] [PubMed]
- Alakomi, H.L.; Paananen, A.; Suihko, M.L.; Helander, I.M.; Saarela, M. Weakening effect of cell permeabilizers on gram-negative bacteria causing biodeterioration. Appl. Environ. Microbiol. 2006, 72, 4695–4703. [Google Scholar] [CrossRef] [PubMed]
- Alakomi, H.L.; Skytta, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic acid permeabilizes gram-negative bacteria by disrupting the outer membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef]
- Barbau-Piednoir, E.; Denayer, S.; Botteldoorn, N.; Dierick, K.; De Keersmaecker, S.C.; Roosens, N.H. Detection and discrimination of five E. coli pathotypes using a combinatory SYBR® Green qPCR screening system. Appl. Microbiol. Biotechnol. 2018, 102, 3267–3285. [Google Scholar] [CrossRef] [PubMed]
- Biotium Site. Available online: https://biotium.com/free-sample/ (accessed on 15 January 2024).
- Wang, L.; Li, Y.; Mustapha, A. Detection of viable Escherichia coli O157:H7 by ethidium monoazide real-time PCR. J. Appl. Microbiol. 2009, 107, 1719–1728. [Google Scholar] [CrossRef]
- Pan, Y.O.; Breidt, F., Jr. Enumeration of viable Listeria monocytogenes cells by real-time PCR with propidium monoazide and ethidium monoazide in the presence of dead cells. Appl. Environ. Microbiol. 2007, 73, 8028–8031. [Google Scholar] [CrossRef]
- Lee, A.S.; Lamanna, O.K.; Ishida, K.; Hill, E.; Nguyen, A.; Hsieh, M.H. A novel propidium monoazide-based PCR assay can measure viable uropathogenic E. coli in vitro and in vivo. Front. Cell. Infect. Microbiol. 2022, 12, 794323. [Google Scholar] [CrossRef]
- Ouf, J.M.; Yuan, Y.; Singh, P.; Mustapha, A. Detection of viable but nonculturable Escherichia coli O157:H7 in ground beef by propidium monoazide real-time PCR. Detection of Viable but Nonculturable Escherichia coli O157:H7 in Ground Beef by Propidium Monoazide real-time PCR. Int. J. Agric. Sci. Food Technol. 2017, 3, 26–31. [Google Scholar]
- Taskin, B.; Gozen, A.G.; Duran, M. Selective quantification of viable Escherichia coli bacteria in biosolids by quantitative PCR with propidium monoazide modification. Appl. Environ. Microbiol. 2011, 77, 4329–4335. [Google Scholar] [CrossRef]
- Golpayegani, A.; Douraghi, M.; Rezaei, F.; Alimohammadi, M.; Nodehi, R.N. Propidium monoazide–quantitative polymerase chain reaction (PMA-qPCR) assay for rapid detection of viable and viable but non-culturable (VBNC) Pseudomonas aeruginosa in swimming pools. J. Environ. Health Sci. Eng. 2019, 17, 407–416. [Google Scholar] [CrossRef]
- Cechova, M.; Beinhauerova, M.; Babak, V.; Kralik, P. A viability assay combining palladium compound treatment with quantitative PCR to detect viable Mycobacterium avium subsp. paratuberculosis cells. Sci. Rep. 2022, 12, 4769. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wei, Y.; Shang, N.; Li, P. Synergistic inhibition of plantaricin E/F and lactic acid against Aeromonas hydrophila LPL-1 reveals the novel potential of class IIb bacteriocin. Front. Microbiol. 2022, 13, 774184. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Li, T.; Xie, X.; Feng, Y.; Chen, Z.; Yang, H.; Wu, C.; Deng, S.; Liu, Y. PLGA-Based Drug Delivery Systems for Remotely Triggered Cancer Therapeutic and Diagnostic Applications. Front. Bioeng. Biotechnol. 2020, 8, 381. [Google Scholar] [CrossRef]
- Labrador, M.; Giménez-Rota, C.; Rota, C. Real-time PCR method combined with a matrix lysis procedure for the quantification of Listeria monocytogenes in meat products. Foods 2021, 10, 735. [Google Scholar] [CrossRef] [PubMed]
- Vamanu, E.; Pop, M.I.; Pop, I.O.; Mishra, S.K.; Singh, K.S. Therapeutic potential of functional product based on wild edible mushrooms. AgroLife Sci. J. 2023, 12, 229–235. [Google Scholar] [CrossRef]
- Hodzic, E.; Glavinic, A.; Wademan, C. A novel approach for simultaneous detection of the most common food-borne pathogens by multiplex qPCR. Biomol. Biomed. 2023, 23, 640. [Google Scholar] [CrossRef]
- Suther, C.; Moore, M.D. Quantification and discovery of PCR inhibitors found in food matrices commonly associated with foodborne viruses. Food Sci. Hum. Wellness 2019, 8, 351–355. [Google Scholar] [CrossRef]
- Siala, M.; Barbana, A.; Smaoui, S.; Hachicha, S.; Marouane, C.; Kammoun, S.; Gdoura, R.; Messadi-Akrout, F. Screening and detecting Salmonella in different food matrices in Southern Tunisia using a combined enrichment/real-time PCR method: Correlation with conventional culture method. Front. Microbiol. 2017, 8, 2416. [Google Scholar] [CrossRef] [PubMed]
- Løvdal, T.; Hovda, M.B.; Björkblom, B.; Møller, S.G. Propidium monoazide combined with real-time quantitative PCR underestimates heat-killed Listeria innocua. J. Microbiol. Methods 2011, 85, 164–169. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinu, L.-D.; Al-Zaidi, Q.J.; Matache, A.G.; Matei, F. Improving the Efficiency of Viability-qPCR with Lactic Acid Enhancer for the Selective Detection of Live Pathogens in Foods. Foods 2024, 13, 1021. https://doi.org/10.3390/foods13071021
Dinu L-D, Al-Zaidi QJ, Matache AG, Matei F. Improving the Efficiency of Viability-qPCR with Lactic Acid Enhancer for the Selective Detection of Live Pathogens in Foods. Foods. 2024; 13(7):1021. https://doi.org/10.3390/foods13071021
Chicago/Turabian StyleDinu, Laura-Dorina, Quthama Jasim Al-Zaidi, Adelina Georgiana Matache, and Florentina Matei. 2024. "Improving the Efficiency of Viability-qPCR with Lactic Acid Enhancer for the Selective Detection of Live Pathogens in Foods" Foods 13, no. 7: 1021. https://doi.org/10.3390/foods13071021
APA StyleDinu, L.-D., Al-Zaidi, Q. J., Matache, A. G., & Matei, F. (2024). Improving the Efficiency of Viability-qPCR with Lactic Acid Enhancer for the Selective Detection of Live Pathogens in Foods. Foods, 13(7), 1021. https://doi.org/10.3390/foods13071021