

The Investigation of Changes in Bacterial Community of Pasteurized Milk during Cold Storage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Treatment of Milk Samples
2.2. Culture-Based Microbial Identification
2.3. DNA Extraction and Pyrosequencing 16S rRNA
2.4. The Examination of Bacterial Communities by High-Throughput Sequencing Analysis
2.5. Statistical Analysis
3. Results
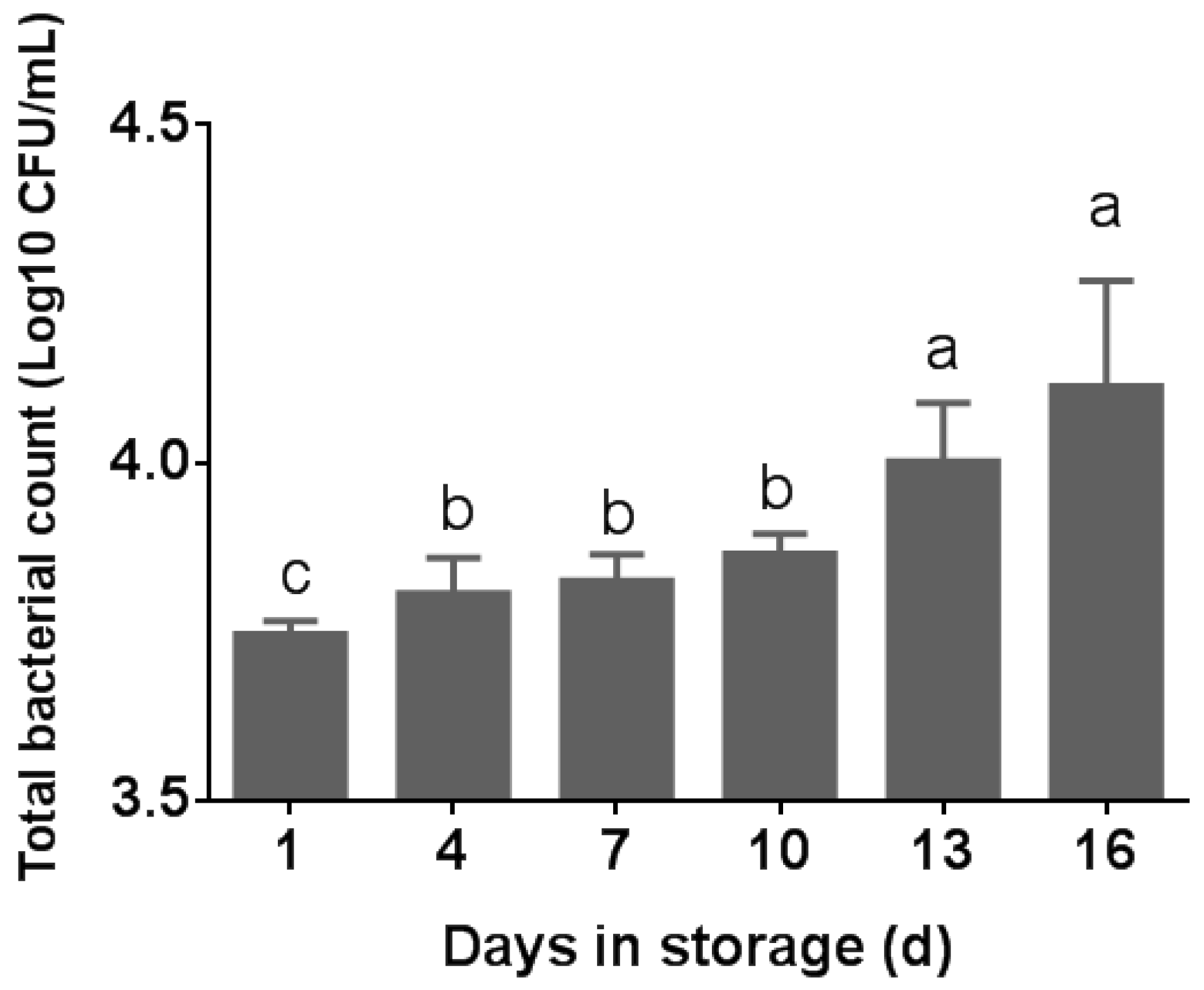
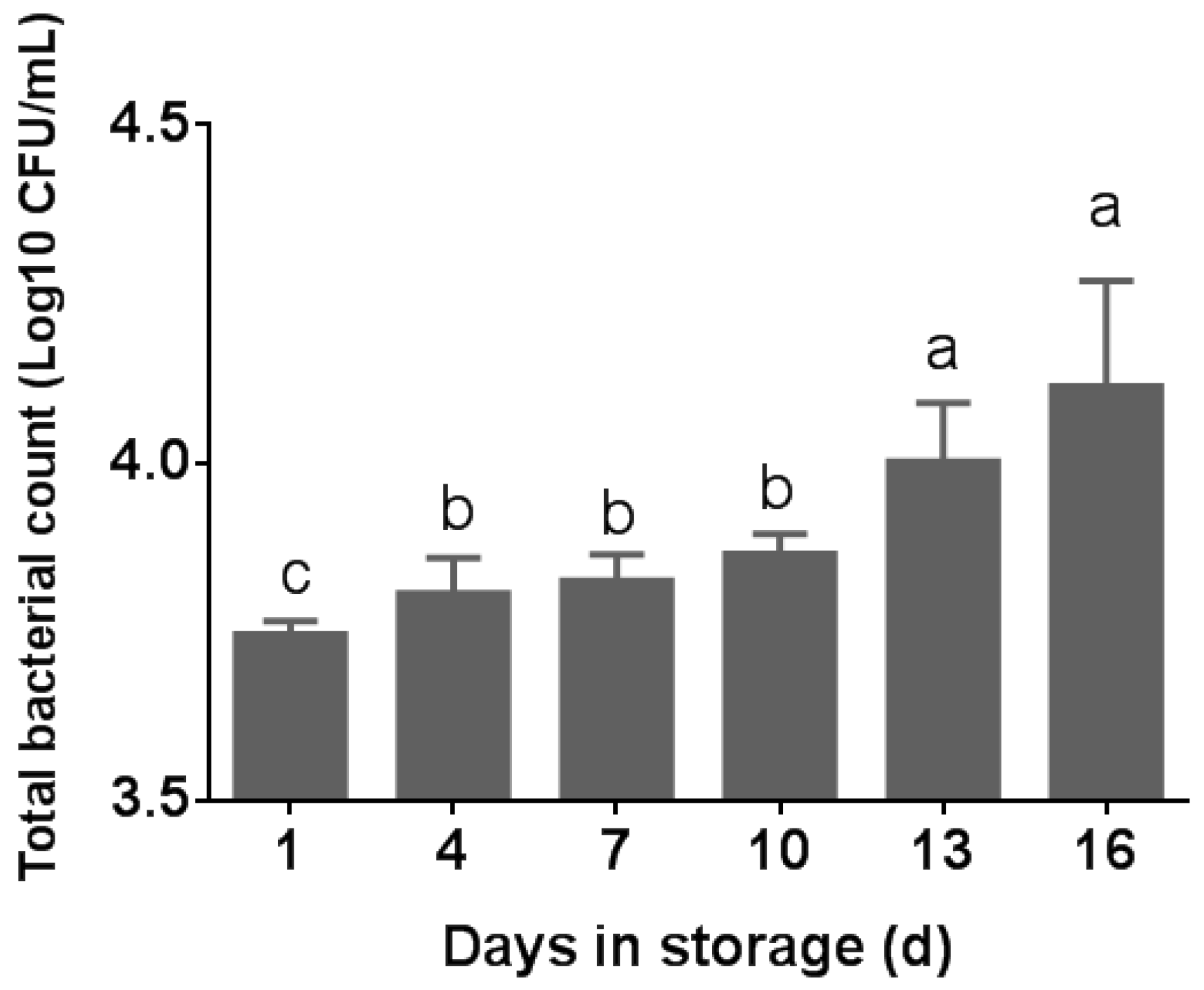
3.1. Total Bacterial Counts in Pasteurized Milk
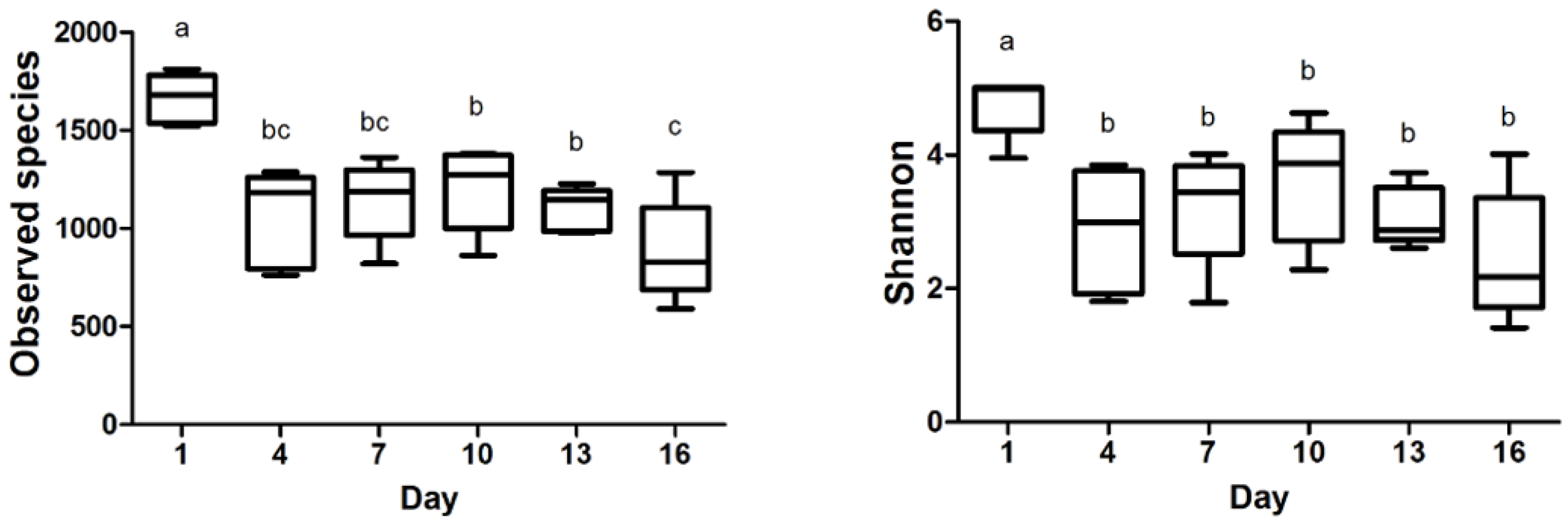
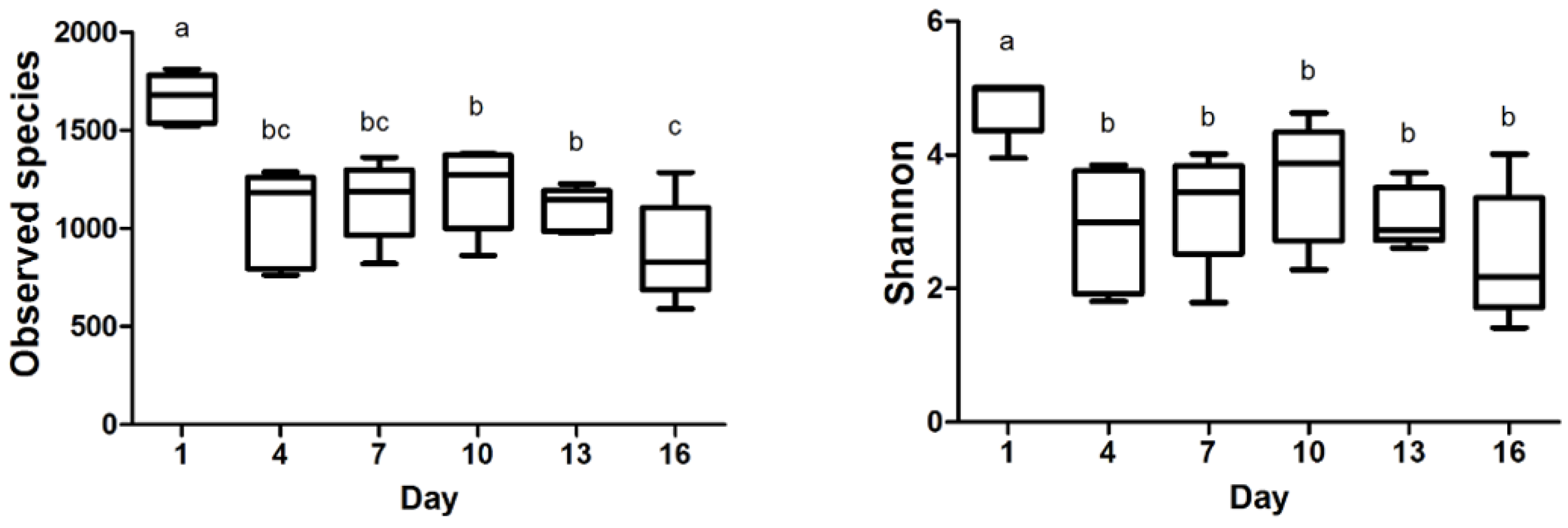
3.2. Bacterial Diversity
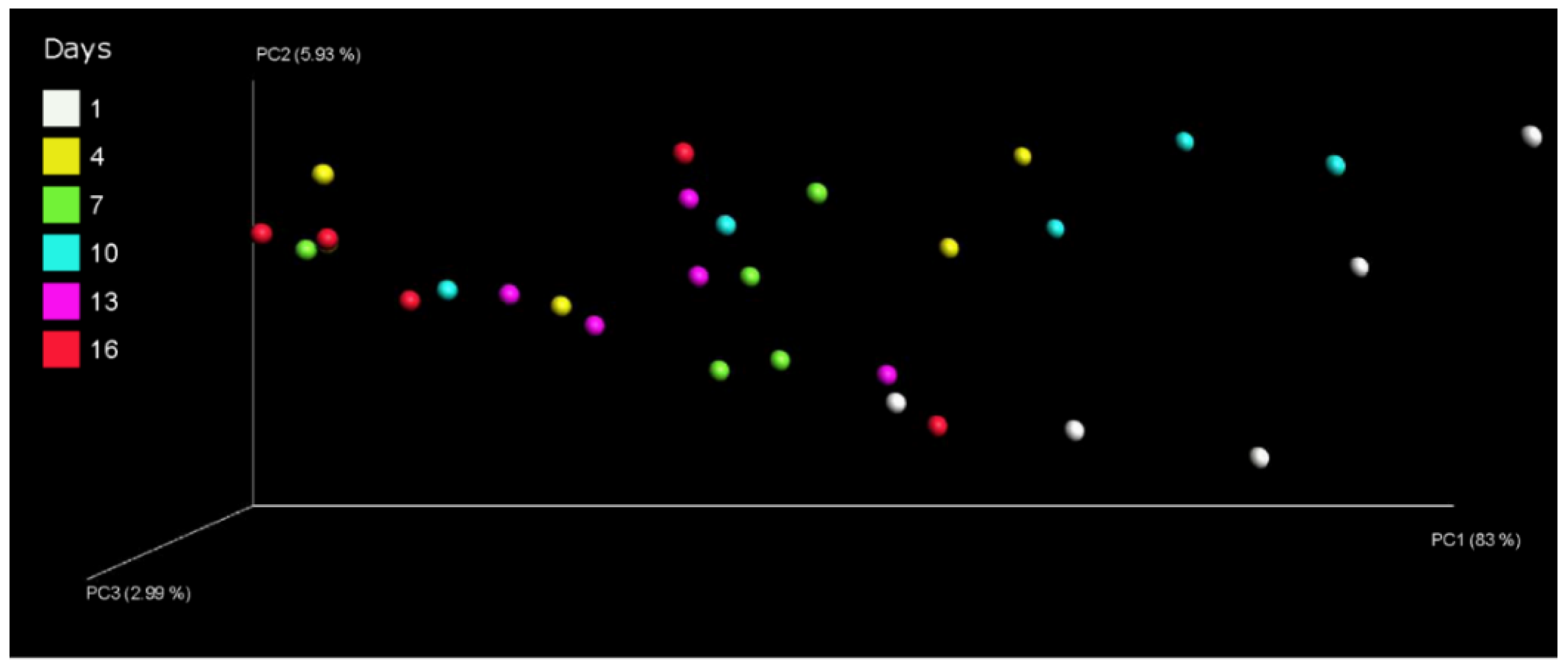
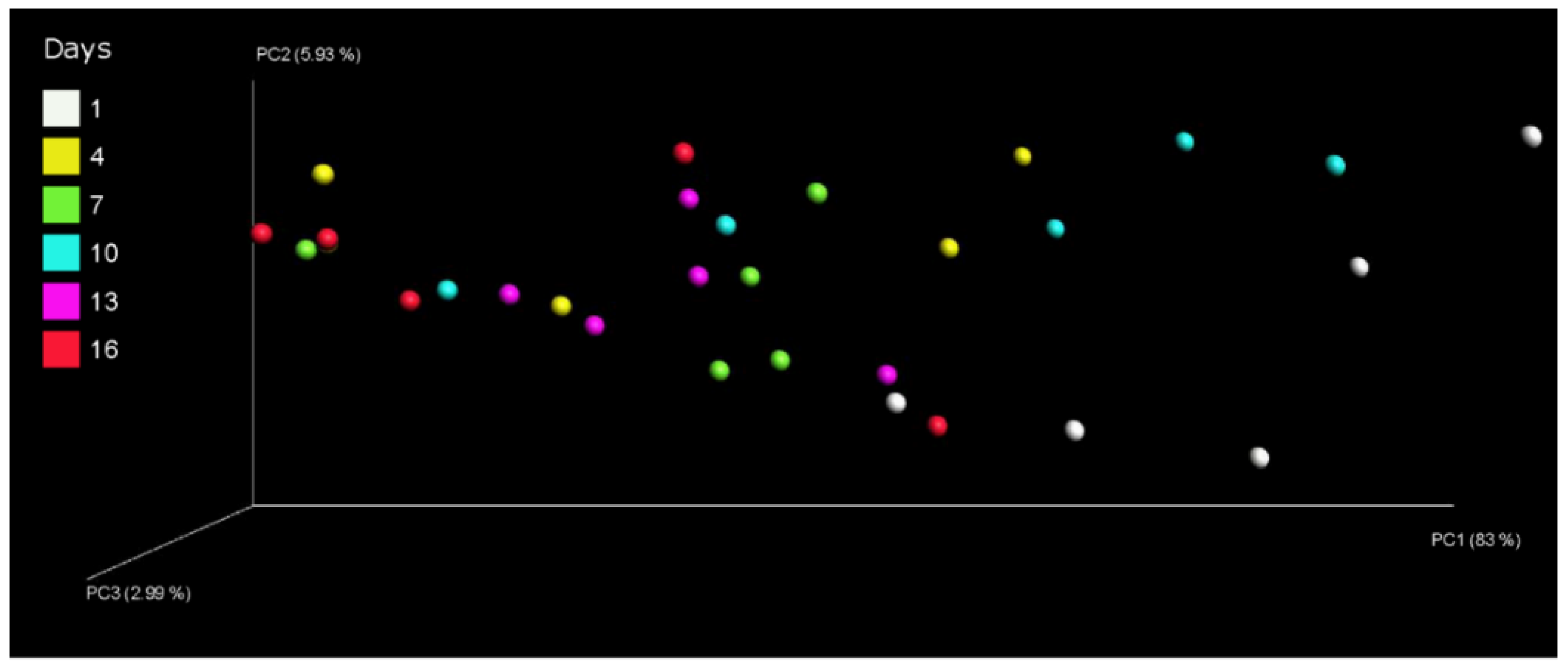
3.3. Effect of Storage Time on the Bacterial Communities in Pasteurized Milk
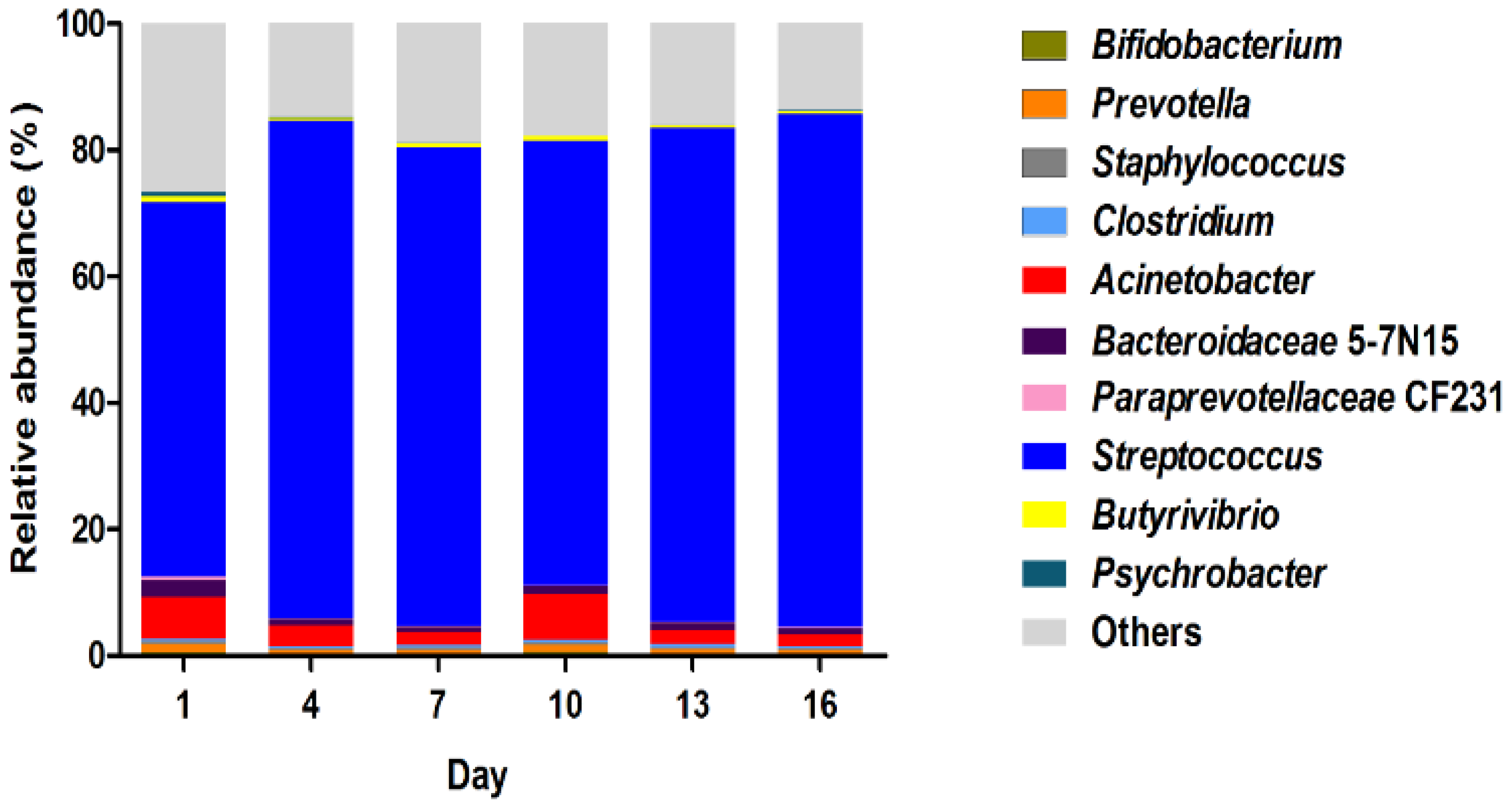
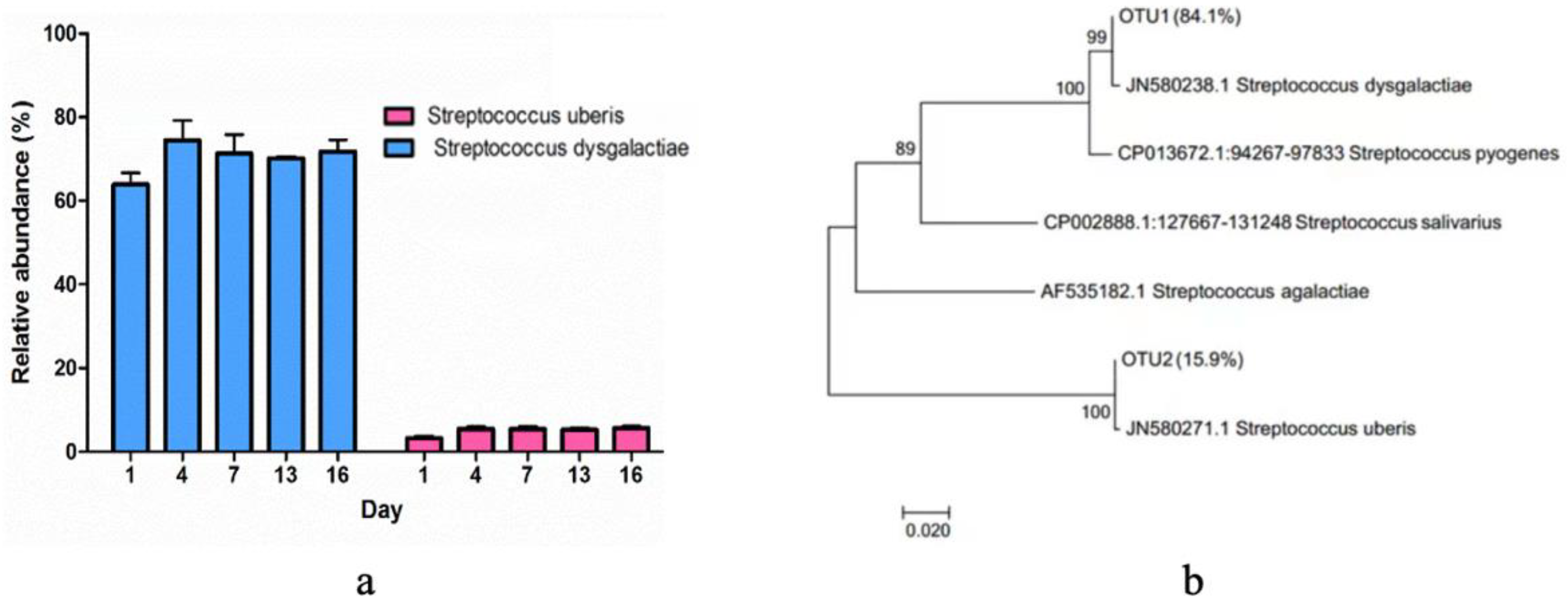
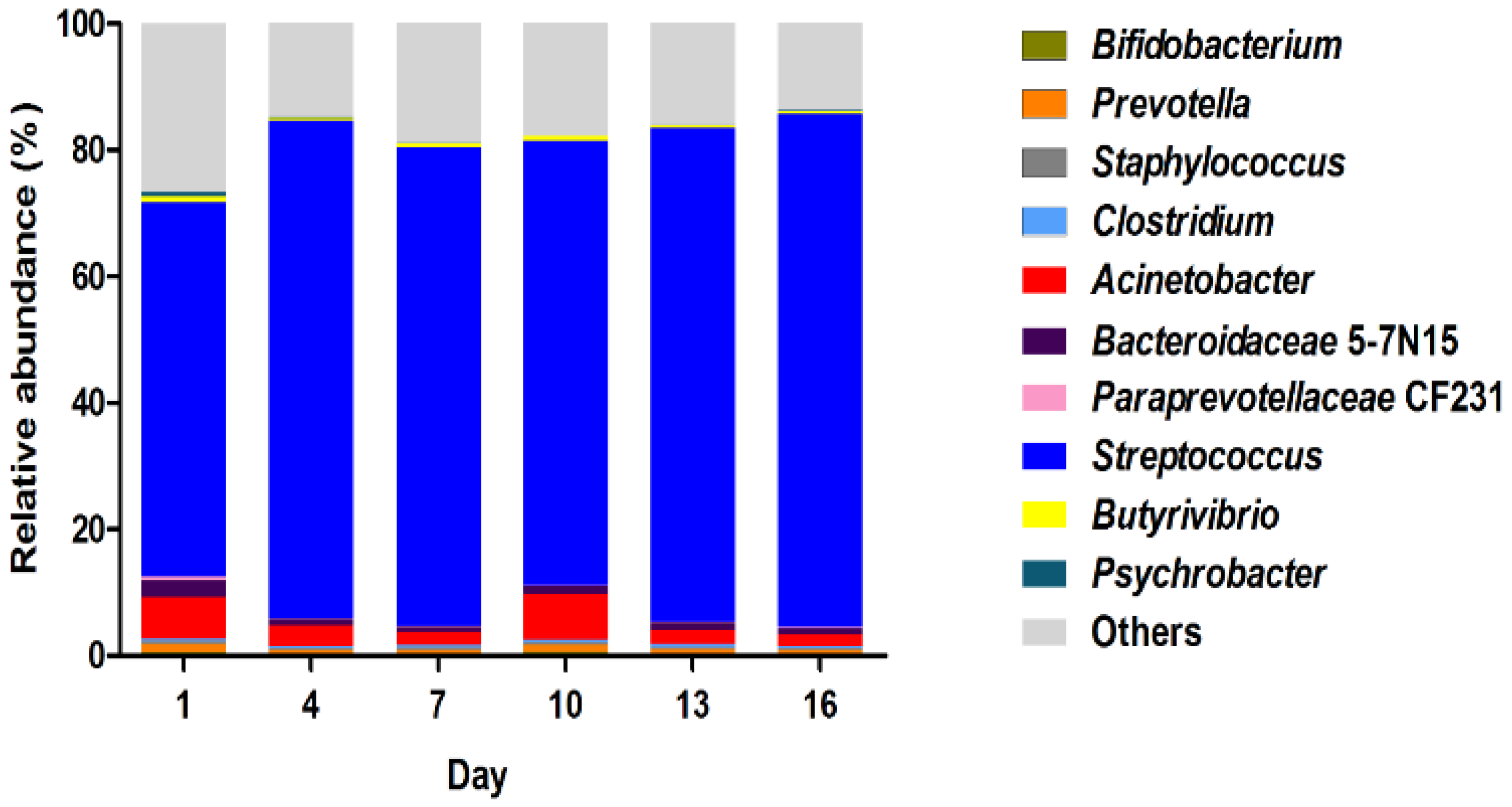
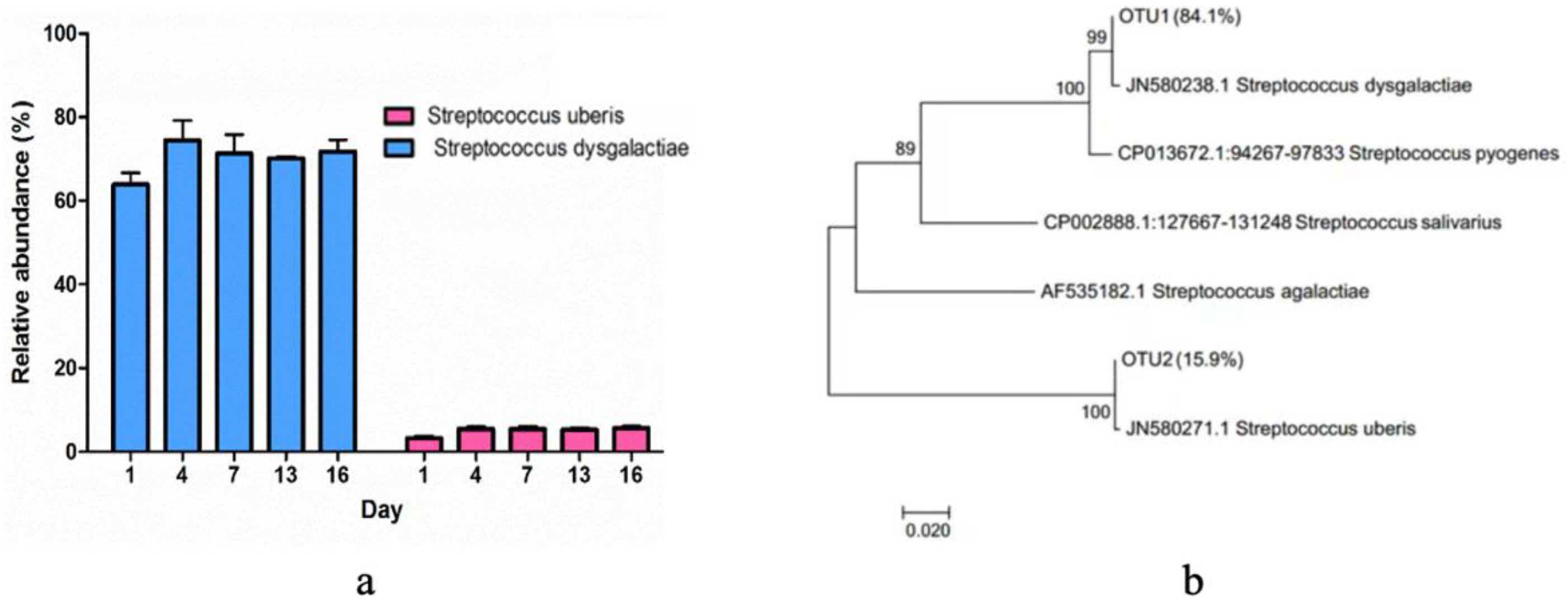
3.4. The Composition of Bacterial Community
4. Discussion
4.1. Effects of Storage on Bacteria in Pasteurized Milk
4.2. Effects of Storage on the Bacterial Community Diversity of Pasteurized Milk
4.3. The Bacterial Community Composition in Pasteurized Milk during the Storage
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bosica, S.; Chiaverini, A.; De Angelis, M.E.; Petrini, A.; Averaimo, D.; Martino, M.; Rulli, M.; Saletti, M.A.; Cantelmi, M.C.; Ruggeri, F.; et al. Severe Streptococcus equi Subspecies zooepidemicus Outbreak from Unpasteurized Dairy Product Consumption, Italy. Emerg. Infect. Dis. 2023, 29, 1020–1024. [Google Scholar] [CrossRef] [PubMed]
- Porcellato, D.; Aspholm, M.; Skeie, S.B.; Monshaugen, M.; Brendehaug, J.; Mellegard, H. Microbial diversity of consumption milk during processing and storage. Int. J. Food Microbiol. 2018, 266, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.; Liu, Y.; Yang, S.; Liu, Y.; Shi, H.; Yue, X.; Wu, R.; Wu, J. High-throughput sequencing provides new insights into the roles and implications of core microbiota present in pasteurized milk. Food Res. Int. 2020, 137, 109586. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Weng, P.; Wu, Z.; Liu, Y. Extending the Shelf Life of Raw Milk and Pasteurized Milk with Plantaricin FB-2. Foods 2023, 12, 608. [Google Scholar] [CrossRef] [PubMed]
- Endara, P.; Wiedmann, M.; Adalja, A. Consumer willingness to pay for shelf life of high temperature, short time pasteurized fluid milk: Implications for smart labeling and food waste reduction. J. Dairy Sci. 2023, 106, 5940–5957. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Chieffi, D.; Fanelli, F.; Logrieco, A.F.; Cho, G.S.; Kabisch, J.; Bohnlein, C.; Franz, C. Microbial quality and safety of milk and milk products in the 21st century. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2013–2049. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, Y.; You, C.; Ren, J.; Chen, W.; Zheng, H.; Liu, Z. Variation in Raw Milk Microbiota Throughout 12 Months and the Impact of Weather Conditions. Sci. Rep. 2018, 8, 2371. [Google Scholar] [CrossRef]
- Rossvoll, E.; Ronning, H.T.; Granum, P.E.; Moretro, T.; Hjerpekjon, M.R.; Langsrud, S. Toxin production and growth of pathogens subjected to temperature fluctuations simulating consumer handling of cold cuts. Int. J. Food Microbiol. 2014, 185, 82–92. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, Y.; Jay-Russell, M.; Lemay, D.G.; Mills, D.A. Reservoirs of antimicrobial resistance genes in retail raw milk. Microbiome 2020, 8, 99. [Google Scholar] [CrossRef]
- Ranvir, S.; Sharma, R.; Gandhi, K.; Mann, B. Assessment of physico-chemical changes in UHT milk during storage at different temperatures. J. Dairy Res. 2020, 87, 243–247. [Google Scholar] [CrossRef]
- Lott, T.T.; Wiedmann, M.; Martin, N.H. Shelf-life storage temperature has a considerably larger effect than high-temperature, short-time pasteurization temperature on the growth of spore-forming bacteria in fluid milk. J. Dairy Sci. 2023, 106, 3838–3855. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, C.; Ye, H.; Wang, Z.; Wu, X.; Han, Y.; Xu, B. Changes in the microbial communities in vacuum-packaged smoked bacon during storage. Food Microbiol. 2019, 77, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.; Normanno, G.; Capozzi, L.; Del Sambro, L.; Di Fato, L.; Miccolupo, A.; Di Taranto, P.; Caruso, M.; Petruzzi, F.; Ali, A.; et al. High Genetic Diversity and Virulence Potential in Bacillus cereus sensu lato Isolated from Milk and Cheeses in Apulia Region, Southern Italy. Foods 2023, 12, 1548. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.Y.; Zhao, S.G.; Zheng, N.; Li, S.L.; Zhang, Y.D.; Liu, H.M.; McKillip, J.; Wang, J.Q. Short communication: Microbiological quality of raw cow milk and its association with herd management practices in Northern China. J. Dairy Sci. 2017, 100, 4294–4299. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Zhao, S.; Zheng, N.; Bu, D.; Beckers, Y.; Denman, S.E.; McSweeney, C.S.; Wang, J. Differences in Ureolytic Bacterial Composition between the Rumen Digesta and Rumen Wall Based on ureC Gene Classification. Front. Microbiol. 2017, 8, 385. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- Du, B.; Meng, L.; Wu, H.; Yang, H.; Liu, H.; Zheng, N.; Zhang, Y.; Zhao, S.; Wang, J. Source Tracker Modeling Based on 16S rDNA Sequencing and Analysis of Microbial Contamination Sources for Pasteurized Milk. Front. Nutr. 2022, 9, 845150. [Google Scholar] [CrossRef]
- Edwards, K.M.; Badiger, A.; Heldman, D.R.; Klein, M.S. Metabolomic Markers of Storage Temperature and Time in Pasteurized Milk. Metabolites 2021, 11, 419. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.H.; Ranieri, M.L.; Wiedmann, M.; Boor, K.J. Reduction of pasteurization temperature leads to lower bacterial outgrowth in pasteurized fluid milk during refrigerated storage: A case study. J. Dairy Sci. 2012, 95, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.; Trmcic, A.; Martin, N.H.; Wiedmann, M.; Murphy, S.I. Development of a Monte Carlo simulation model to predict pasteurized fluid milk spoilage due to post-pasteurization contamination with gram-negative bacteria. J. Dairy Sci. 2022, 105, 1978–1998. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Cui, C.; Li, X.; Yan, J.; Sun, E.; Wang, C.; Guo, H.; Hao, Y. Prevalence, antimicrobial susceptibility, and antibiotic resistance gene transfer of Bacillus strains isolated from pasteurized milk. J. Dairy Sci. 2023, 106, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Yu, S.; Yu, P.; Liu, C.; Kong, L.; Feng, Z.; et al. Prevalence, Virulence Genes, Antimicrobial Susceptibility, and Genetic Diversity of Bacillus cereus Isolated from Pasteurized Milk in China. Front. Microbiol. 2018, 9, 533. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Chen, J.; Fei, P.; Feng, H.; Wang, Y.; Ali, M.A.; Li, S.; Jing, H.; Yang, W. Prevalence, molecular characterization, and antibiotic susceptibility of Bacillus cereus isolated from dairy products in China. J. Dairy Sci. 2020, 103, 3994–4001. [Google Scholar] [CrossRef]
- Porcellato, D.; Aspholm, M.; Skeie, S.B.; Mellegard, H. Application of a novel amplicon-based sequencing approach reveals the diversity of the Bacillus cereus group in stored raw and pasteurized milk. Food Microbiol. 2019, 81, 32–39. [Google Scholar] [CrossRef]
- Masiello, S.N.; Kent, D.; Martin, N.H.; Schukken, Y.H.; Wiedmann, M.; Boor, K.J. Longitudinal assessment of dairy farm management practices associated with the presence of psychrotolerant Bacillales spores in bulk tank milk on 10 New York State dairy farms. J. Dairy Sci. 2017, 100, 8783–8795. [Google Scholar] [CrossRef]
- Kobayashi, T.; Azuma, T.; Yasokawa, D.; Yamaki, S.; Yamazaki, K. Spore Heat Resistance and Growth Ability at Refrigeration Temperatures of Bacillus spp. and Paenibacillus spp. Biocontrol Sci. 2021, 26, 147–155. [Google Scholar] [CrossRef]
- Oikonomou, G.; Addis, M.F.; Chassard, C.; Nader-Macias, M.E.F.; Grant, I.; Delbes, C.; Bogni, C.I.; Le Loir, Y.; Even, S. Milk Microbiota: What Are We Exactly Talking About? Front. Microbiol. 2020, 11, 60. [Google Scholar] [CrossRef]
- Falentin, H.; Rault, L.; Nicolas, A.; Bouchard, D.S.; Lassalas, J.; Lamberton, P.; Aubry, J.M.; Marnet, P.G.; Le Loir, Y.; Even, S. Bovine Teat Microbiome Analysis Revealed Reduced Alpha Diversity and Significant Changes in Taxonomic Profiles in Quarters with a History of Mastitis. Front. Microbiol. 2016, 7, 480. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Kable, M.E.; Marco, M.L. Impact of DNA Sequencing and Analysis Methods on 16S rRNA Gene Bacterial Community Analysis of Dairy Products. mSphere 2018, 3, e00410-18. [Google Scholar] [CrossRef] [PubMed]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The complex microbiota of raw milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yu, Z.; Wang, J.; Ho, H.; Yang, Y.; Fan, R.; Du, Q.; Jiang, H.; Han, R. Prevalence, Drug Resistance, and Virulence Genes of Potential Pathogenic Bacteria in Pasteurized Milk of Chinese Fresh Milk Bar. J. Food Prot. 2021, 84, 1863–1867. [Google Scholar] [CrossRef]
- Phuektes, P.; Browning, G.F.; Anderson, G.; Mansell, P.D. Multiplex polymerase chain reaction as a mastitis screening test for Staphylococcus aureus, Streptococcus agalactiae, Streptococcus dysgalactiae and Streptococcus uberis in bulk milk samples. J. Dairy Res. 2003, 70, 149–155. [Google Scholar] [CrossRef]
- Nakata, H.; Imamura, Y.; Saha, S.; Lobo, R.E.; Kitahara, S.; Araki, S.; Tomokiyo, M.; Namai, F.; Hiramitsu, M.; Inoue, T.; et al. Partial Characterization and Immunomodulatory Effects of Exopolysaccharides from Streptococcus thermophilus SBC8781 during Soy Milk and Cow Milk Fermentation. Foods 2023, 12, 2374. [Google Scholar] [CrossRef]
- McHugh, A.J.; Feehily, C.; Fenelon, M.A.; Gleeson, D.; Hill, C.; Cotter, P.D. Tracking the Dairy Microbiota from Farm Bulk Tank to Skimmed Milk Powder. mSystems 2020, 5, 10-1128. [Google Scholar] [CrossRef]
- Sun, L.; Lundh, A.; Hojer, A.; Bernes, G.; Nilsson, D.; Johansson, M.; Hetta, M.; Gustafsson, A.H.; Saeden, K.H.; Dicksved, J. Milking system and premilking routines have a strong effect on the microbial community in bulk tank milk. J. Dairy Sci. 2022, 105, 123–139. [Google Scholar] [CrossRef]
- Al-Farsi, M.; Al-Gharibi, I.; Al-Abri, A.; Al-Humaimi, A.; Al-Nabhani, F.; Al-Hashmi, H.; Al-Sarmi, K.; Al-Shibli, S. Evaluating the shelf-life of pasteurized milk in Oman. Heliyon 2021, 7, e06555. [Google Scholar] [CrossRef]
- Li, S.N.; Tang, S.H.; Ren, R.; Gong, J.X.; Chen, Y.M. Metabolomic profile of milk fermented with Streptococcus thermophilus cocultured with Bifidobacterium animalis ssp. lactis, Lactiplantibacillus plantarum, or both during storage. J. Dairy Sci. 2021, 104, 8493–8505. [Google Scholar] [CrossRef]
- Cho, G.S.; Li, B.; Rostalsky, A.; Fiedler, G.; Rosch, N.; Igbinosa, E.; Kabisch, J.; Bockelmann, W.; Hammer, P.; Huys, G.; et al. Diversity and Antibiotic Susceptibility of Acinetobacter Strains From Milk Powder Produced in Germany. Front. Microbiol. 2018, 9, 536. [Google Scholar] [CrossRef] [PubMed]
- Minst, K.; Martlbauer, E.; Miller, T.; Meyer, C. Short communication: Streptococcus species isolated from mastitis milk samples in Germany and their resistance to antimicrobial agents. J. Dairy Sci. 2012, 95, 6957–6962. [Google Scholar] [CrossRef] [PubMed]
- Crippa, B.L.; Rodrigues, M.X.; Tomazi, T.; Yang, Y.; de Oliveira Rocha, L.; Bicalho, R.C.; Silva, N.C.C. Virulence factors, antimicrobial resistance and phylogeny of bovine mastitis-associated Streptococcus dysgalactiae. J. Dairy Res. 2023, 90, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Wente, N.; Kromker, V. Streptococcus dysgalactiae-Contagious or Environmental? Animals 2020, 10, 2185. [Google Scholar] [CrossRef] [PubMed]
- Lan, R.; Zhou, Y.; Wang, Z.; Fu, S.; Gao, Y.; Gao, X.; Zhang, J.; Han, X.; Phouthapane, V.; Xu, Y.; et al. Reduction of ROS-HIF1alpha-driven glycolysis by taurine alleviates Streptococcus uberis infection. Food Funct. 2022, 13, 1774–1784. [Google Scholar] [CrossRef]
- Mihklepp, K.; Kivirand, K.; Juronen, D.; Lookene, A.; Rinken, T. Immunodetection of Streptococcus uberis pathogen in raw milk. Enzyme Microb. Technol. 2019, 130, 109360. [Google Scholar] [CrossRef]
- Achek, R.; El-Adawy, H.; Hotzel, H.; Tomaso, H.; Ehricht, R.; Hamdi, T.M.; Azzi, O.; Monecke, S. Short communication: Diversity of staphylococci isolated from sheep mastitis in northern Algeria. J. Dairy Sci. 2020, 103, 890–897. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, X.; Wu, S.; Du, Q.; Min, L. The Investigation of Changes in Bacterial Community of Pasteurized Milk during Cold Storage. Foods 2024, 13, 451. https://doi.org/10.3390/foods13030451
Lan X, Wu S, Du Q, Min L. The Investigation of Changes in Bacterial Community of Pasteurized Milk during Cold Storage. Foods. 2024; 13(3):451. https://doi.org/10.3390/foods13030451
Chicago/Turabian StyleLan, Xinyi, Shuyan Wu, Qijing Du, and Li Min. 2024. "The Investigation of Changes in Bacterial Community of Pasteurized Milk during Cold Storage" Foods 13, no. 3: 451. https://doi.org/10.3390/foods13030451
APA StyleLan, X., Wu, S., Du, Q., & Min, L. (2024). The Investigation of Changes in Bacterial Community of Pasteurized Milk during Cold Storage. Foods, 13(3), 451. https://doi.org/10.3390/foods13030451