
Mass Spectrometry-Based Metabolomics Investigation on Two Different Seaweeds Under Arsenic Exposure

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Arsenic Concentration Determination in Seaweed Samples
2.2.1. Sample Preparation
2.2.2. Calibration Procedure
2.2.3. ICP-MS Analysis
2.2.4. Method Assurance
2.3. Metabolite Extraction
2.4. Non-Targeted Metabolomics by LCMS-QTOF
2.5. Preprocessing of MS Data
2.6. Statistical Analysis
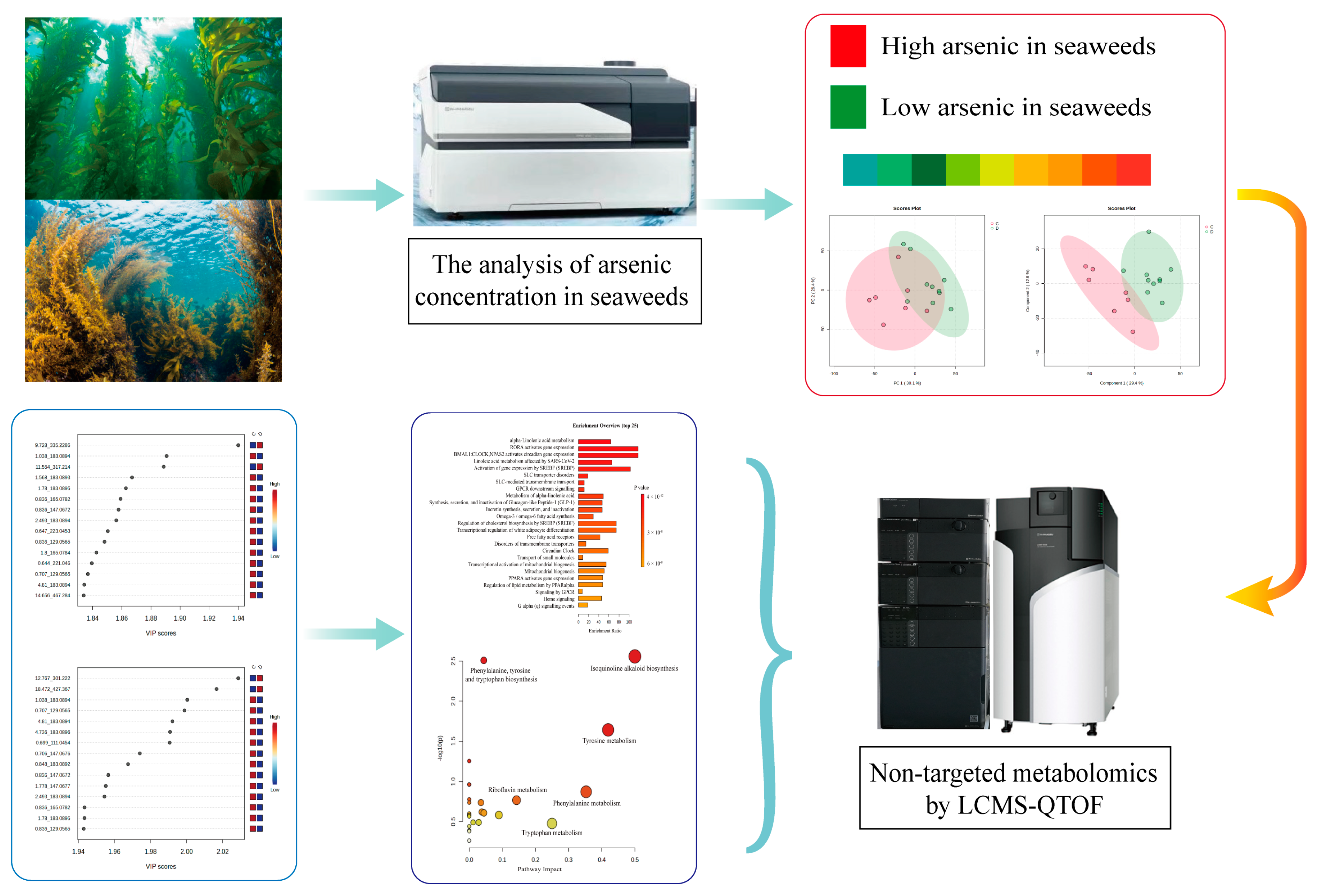
3. Results and Discussion
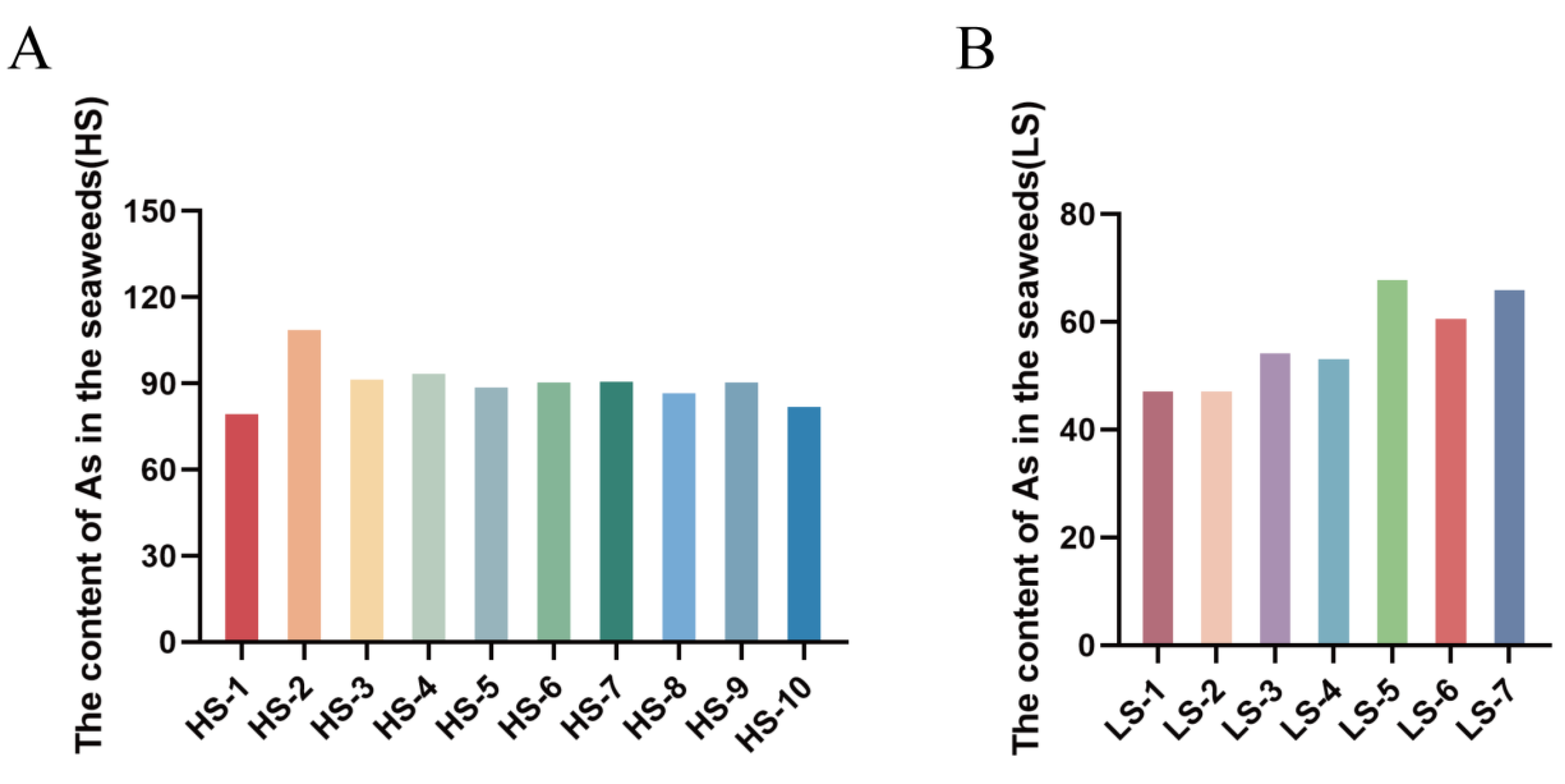
3.1. Arsenic Concentration
3.2. Metabolomic Analysis Based on UPLC-Q-TOF/MS
3.2.1. Metabolic Profiles
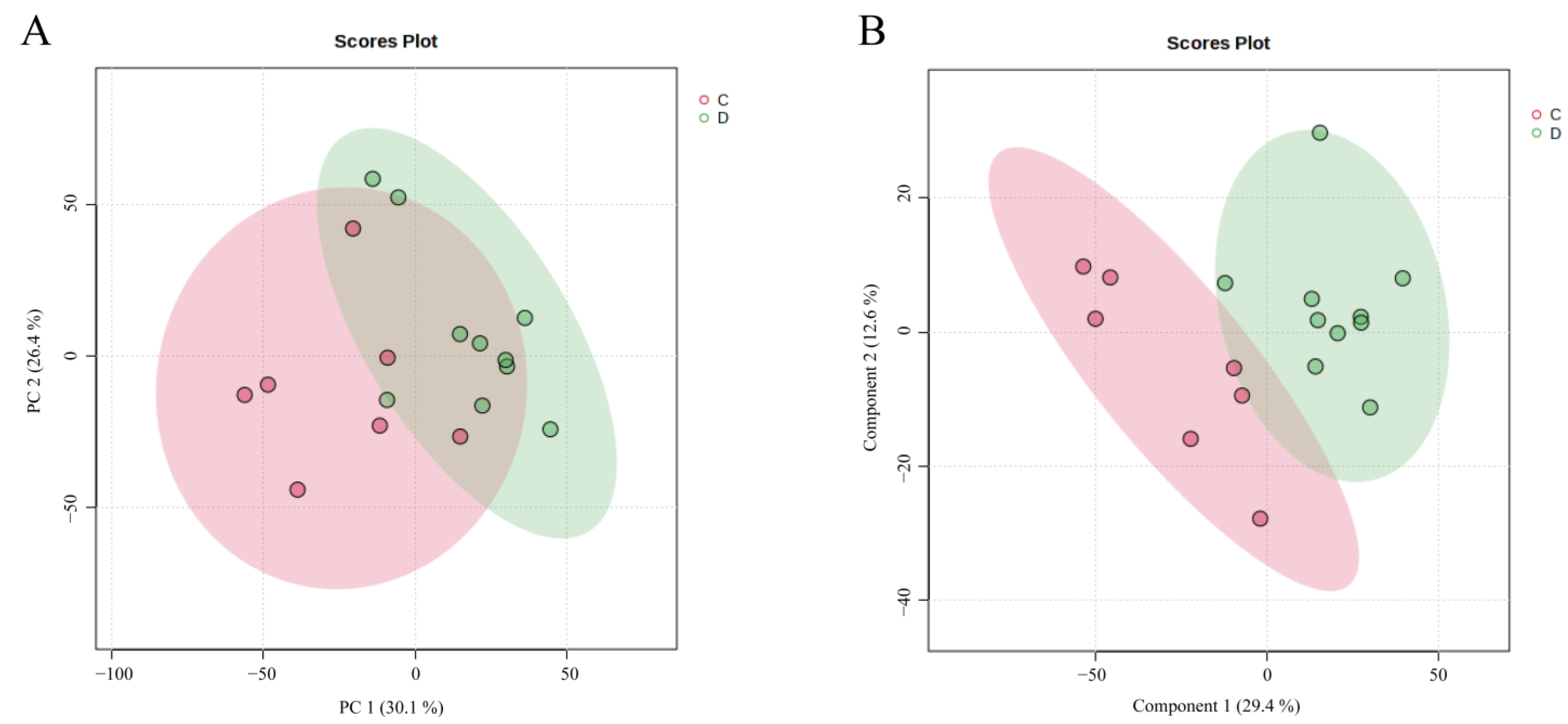
3.2.2. PCA and PLS-DA Models
3.3. Metabolic Pathway
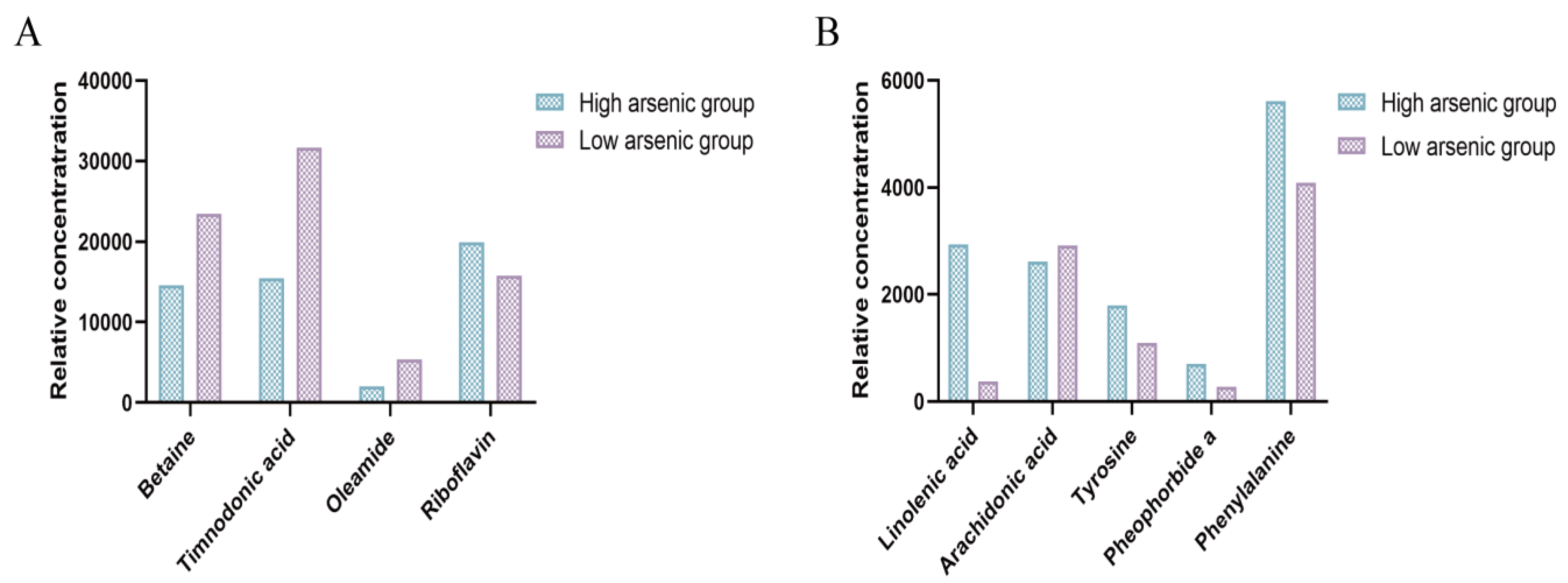
3.4. Targeted Profiles
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Q.Y.; Costa, M. Arsenic: A Global Environmental Challenge. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 47–63. [Google Scholar] [CrossRef]
- Zhang, W.; Guo, Z.; Zhou, Y.; Liu, H.; Zhang, L. Biotransformation and detoxification of inorganic arsenic in Bombay oyster Saccostrea cucullata. Aquat. Toxicol. 2015, 158, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.; Shi, Y.; Wu, D.; Ye, H.; Feng, D.; Huang, D.; Li, S.; Fang, C. Characteristics of arsenic speciation in mainly cultured shellfish from Sanmen Bay, Zhejiang Province, China. Mar. Pollut. Bull. 2023, 197, 115793. [Google Scholar] [CrossRef] [PubMed]
- Ratnaike, R.N. Acute and chronic arsenic toxicity. Postgrad. Med. J. 2003, 79, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; He, Y.; Pan, H.; Liu, H.; Mehmood, K.; Tang, Z.; Hu, L. Toxicity of inorganic arsenic to animals and its treatment strategies. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2023, 271, 109654. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, A.; Sathyapalan, T.; Moallem, S.A.; Sahebkar, A. Counteracting arsenic toxicity: Curcumin to the rescue? J. Hazard. Mater. 2020, 400, 123160. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; He, Y.; Guo, Z.; Wu, Q.; Li, S.; Zhu, Z.; Grimi, N.; Xiao, J. Species-specific arsenic species and health risk assessment in seaweeds from tropic coasts of South China Sea. Ecotoxicol. Environ. Saf. 2023, 267, 115634. [Google Scholar] [CrossRef]
- Escudero-Lourdes, C. Toxicity mechanisms of arsenic that are shared with neurodegenerative diseases and cognitive impairment: Role of oxidative stress and inflammatory responses. Neurotoxicology 2016, 53, 223–235. [Google Scholar] [CrossRef]
- Sidhu, M.S.; Desai, K.P.; Lynch, H.N.; Rhomberg, L.R.; Beck, B.D.; Venditti, F.J. Mechanisms of action for arsenic in cardiovascular toxicity and implications for risk assessment. Toxicology 2015, 331, 78–99. [Google Scholar] [CrossRef]
- Huq, M.E.; Fahad, S.; Shao, Z.; Sarven, M.S.; Khan, I.A.; Alam, M.; Saeed, M.; Ullah, H.; Adnan, M.; Saud, S.; et al. Arsenic in a groundwater environment in Bangladesh: Occurrence and mobilization. J. Environ. Manag. 2020, 262, 110318. [Google Scholar] [CrossRef]
- Ahmad, S.A.; Khan, M.H.; Haque, M. Arsenic contamination in groundwater in Bangladesh: Implications and challenges for healthcare policy. Risk Manag. Healthc. Policy 2018, 11, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Nigra, A.E.; Olmedo, P.; Grau-Perez, M.; O’Leary, R.; O’Leary, M.; Fretts, A.M.; Umans, J.G.; Best, L.G.; Francesconi, K.A.; Goessler, W.; et al. Dietary determinants of inorganic arsenic exposure in the Strong Heart Family Study. Environ. Res. 2019, 177, 108616. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Bi, R.; Musil, S.; Pétursdóttir, Á.H.; Luo, B.; Zhao, P.; Tan, X.; Jia, Y. Arsenic species and their health risks in edible seaweeds collected along the Chinese coastline. Sci. Total Environ. 2022, 847, 157429. [Google Scholar] [CrossRef]
- GB2762-2022; National Food Safety Standard of Maximum Levels of Contaminants in Foods. National Health Commission of the Peolple’s Republic of China: Beijing, China, 2022.
- Luvonga, C.; Rimmer, C.A.; Yu, L.L.; Lee, S.B. Determination of total arsenic and hydrophilic arsenic species in seafood. J. Food Compos. Anal. 2020, 96, 103729. [Google Scholar] [CrossRef] [PubMed]
- Molin, M.; Ulven, S.M.; Meltzer, H.M.; Alexander, J. Arsenic in the human food chain, biotransformation and toxicology--Review focusing on seafood arsenic. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. (GMS) 2015, 31, 249–259. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, J.; Sun, Y.; Chen, S.; Chen, J.; Fu, F. Bio-accessibility and bio-availability evaluation of each arsenic species existing in various edible seaweeds in vitro and in vivo for arsenic risk assessment. Sci. Total Environ. 2024, 946, 174247. [Google Scholar] [CrossRef] [PubMed]
- Preetha, J.S.Y.; Arun, M.; Vidya, N.; Kowsalya, K.; Halka, J.; Ondrasek, G. Biotechnology Advances in Bioremediation of Arsenic: A Review. Molecules 2023, 28, 1474. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.K.; Chang, J.S. Bioremediation of heavy metals using microalgae: Recent advances and mechanisms. Bioresour. Technol. 2020, 303, 122886. [Google Scholar] [CrossRef]
- Liang, P.; Liu, R. Speciation analysis of inorganic arsenic in water samples by immobilized nanometer titanium dioxide separation and graphite furnace atomic absorption spectrometric determination. Anal. Chim. Acta 2007, 602, 32–36. [Google Scholar] [CrossRef]
- Yu, D.X.; Guo, S.; Zhang, X.; Yan, H.; Mao, S.W.; Wang, J.M.; Zhou, J.Q.; Yang, J.; Yuan, Y.W.; Duan, J.A. Combining stable isotope, multielement and untargeted metabolomics with chemometrics to discriminate the geographical origins of ginger (Zingiber officinale Roscoe). Food Chem. 2023, 426, 136577. [Google Scholar] [CrossRef]
- Zhong, P.; Wei, X.; Li, X.; Wei, X.; Wu, S.; Huang, W.; Koidis, A.; Xu, Z.; Lei, H. Untargeted metabolomics by liquid chromatography-mass spectrometry for food authentication: A review. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2455–2488. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Ren, Q.Q.; Lai, Y.H.; Peng, M.Y.; Zhang, J.; Yang, L.T.; Huang, Z.R.; Chen, L.S. Metabolomics combined with physiology and transcriptomics reveals how Citrus grandis leaves cope with copper-toxicity. Ecotoxicol. Environ. Saf. 2021, 223, 112579. [Google Scholar] [CrossRef]
- Jensen, M.; Poulsen, R.; Langebæk, R.; Jenssen, B.M.; Moe, J.; Ciesielski, T.M.; Dietz, R.; Sonne, C.; Madsen, J.; Hansen, M. The metabolome of pink-footed goose: Heavy metals and lipid metabolism. Environ. Res. 2023, 231 Pt 1, 116043. [Google Scholar] [CrossRef]
- Bonanno, G.; Orlando-Bonaca, M. Chemical elements in Mediterranean macroalgae. A review. Ecotoxicol. Environ. Saf. 2018, 148, 44–71. [Google Scholar] [CrossRef]
- Marschner, K.; Musil, S.; Mikšík, I.; Dědina, J. Investigation of hydride generation from arsenosugars—Is it feasible for speciation analysis? Anal. Chim. Acta 2018, 1008, 8–17. [Google Scholar] [CrossRef]
- Kim, K.B.; Nam, Y.A.; Kim, H.S.; Hayes, A.W.; Lee, B.M. α-Linolenic acid: Nutraceutical, pharmacological and toxicological evaluation. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2014, 70, 163–178. [Google Scholar] [CrossRef]
- Kim, O.Y.; Song, J. Important roles of linoleic acid and α-linolenic acid in regulating cognitive impairment and neuropsychiatric issues in metabolic-related dementia. Life Sci. 2024, 337, 122356. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Aleman, G.H.; Castro, V.; Londaitsbehere, A.; Gutierrez-Rodríguez, M.; Garaigorta, U.; Solano, R.; Gastaminza, P. SARS-CoV-2 Fears Green: The Chlorophyll Catabolite Pheophorbide A Is a Potent Antiviral. Pharmaceuticals 2021, 14, 1048. [Google Scholar] [CrossRef] [PubMed]
- Valera, M.J.; Olivera, V.; Pérez, G.; Boido, E.; Dellacassa, E.; Carrau, F. Impact of phenylalanine on Hanseniaspora vineae aroma metabolism during wine fermentation. Int. J. Food Microbiol. 2024, 415, 110631. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, Y.; Li, Q.; Li, J.; Zhang, D.; Zhang, Y.; Li, Y.; Li, X.; Xu, P.; Bai, L.; et al. Multi-omics analysis reveals phenylalanine enhance mitochondrial function and hypoxic endurance via LKB1/AMPK activation. J. Transl. Med. 2024, 22, 920. [Google Scholar] [CrossRef]
- Bi, D.; Shi, M.; Hu, Q.; Wang, H.; Lou, D.; Zhang, A.; Hu, Y. LC/MS/MS-Based Liver Metabolomics to Identify Chronic Liver Injury Biomarkers Following Exposure to Arsenic in Rats. Biol. Trace Elem. Res. 2022, 200, 4355–4369. [Google Scholar] [CrossRef]
- Patel, M.; Parida, A.K. Salinity alleviates arsenic stress-induced oxidative damage via antioxidative defense and metabolic adjustment in the root of the halophyte Salvadora persica. Planta 2023, 258, 109. [Google Scholar] [CrossRef]
- Melo, P.C.; Collela, C.F.; Sousa, T.; Pacheco, D.; Cotas, J.; Gonçalves, A.M.M.; Bahcevandziev, K.; Pereira, L. Seaweed-Based Products and Mushroom β-Glucan as Tomato Plant Immunological Inducers. Vaccines 2020, 8, 524. [Google Scholar] [CrossRef]
- Suwannasom, N.; Kao, I.; Pruß, A.; Georgieva, R.; Bäumler, H. Riboflavin: The Health Benefits of a Forgotten Natural Vitamin. Int. J. Mol. Sci. 2020, 21, 950. [Google Scholar] [CrossRef]
- Thakur, K.; Tomar, S.K.; Singh, A.K.; Mandal, S.; Arora, S. Riboflavin and health: A review of recent human research. Crit. Rev. Food Sci. Nutr. 2017, 57, 3650–3660. [Google Scholar] [CrossRef]
- von Schacky, C. Importance of EPA and DHA Blood Levels in Brain Structure and Function. Nutrients 2021, 13, 1074. [Google Scholar] [CrossRef]
- Bito, T.; Teng, F.; Watanabe, F. Bioactive Compounds of Edible Purple Laver Porphyra sp. (Nori). J. Agric. Food Chem. 2017, 65, 10685–10692. [Google Scholar] [CrossRef]
- Kumar, N.; Singh, D.K.; Bhushan, S.; Jamwal, A. Mitigating multiple stresses in Pangasianodon hypophthalmus with a novel dietary mixture of selenium nanoparticles and Omega-3-fatty acid. Sci. Rep. 2021, 11, 19429. [Google Scholar] [CrossRef] [PubMed]
- Szczuko, M.; Kikut, J.; Komorniak, N.; Bilicki, J.; Celewicz, Z.; Ziętek, M. The Role of Arachidonic and Linoleic Acid Derivatives in Pathological Pregnancies and the Human Reproduction Process. Int. J. Mol. Sci. 2020, 21, 9628. [Google Scholar] [CrossRef] [PubMed]
- Kovács, D.; Camera, E.; Póliska, S.; Cavallo, A.; Maiellaro, M.; Dull, K.; Gruber, F.; Zouboulis, C.C.; Szegedi, A.; Törőcsik, D. Linoleic Acid Induced Changes in SZ95 Sebocytes-Comparison with Palmitic Acid and Arachidonic Acid. Nutrients 2023, 15, 3315. [Google Scholar] [CrossRef]
- El-Ghiaty, M.A.; Alqahtani, M.A.; El-Mahrouk, S.R.; Isse, F.A.; Alammari, A.H.; El-Kadi, A.O.S. Alteration of Hepatic Cytochrome P450 Expression and Arachidonic Acid Metabolism by Arsenic Trioxide (ATO) in C57BL/6 Mice. Biol. Trace Elem. Res. 2024. [Google Scholar] [CrossRef]
- Hernández-Díaz, C.; Juárez-Oropeza, M.A.; Mascher, D.; Pavón, N.; Regla, I.; Paredes-Carbajal, M.C. Effects of Oleamide on the Vasomotor Responses in the Rat. Cannabis Cannabinoid Res. 2020, 5, 42–50. [Google Scholar] [CrossRef]
- Zhao, G.; He, F.; Wu, C.; Li, P.; Li, N.; Deng, J.; Zhu, G.; Ren, W.; Peng, Y. Betaine in Inflammation: Mechanistic Aspects and Applications. Front. Immunol. 2018, 9, 1070. [Google Scholar] [CrossRef]
- Dobrijević, D.; Pastor, K.; Nastić, N.; Özogul, F.; Krulj, J.; Kokić, B.; Bartkiene, E.; Rocha, J.M.; Kojić, J. Betaine as a Functional Ingredient: Metabolism, Health-Promoting Attributes, Food Sources, Applications and Analysis Methods. Molecules 2023, 28, 4824. [Google Scholar] [CrossRef]
- Fu, Y.; Du, H.; Wang, P.; Yin, N.; Cai, X.; Geng, Z.; Li, Y.; Cui, Y. Effects of foods and food components on the in vitro bioaccessibility of total arsenic and arsenic species from Hizikia fusiforme seaweed. Sci. Total Environ. 2023, 900, 165775. [Google Scholar] [CrossRef]
- Wang, T.; Fu, X.; Chen, Q.; Patra, J.K.; Wang, D.; Wang, Z.; Gai, Z. Arachidonic Acid Metabolism and Kidney Inflammation. Int. J. Mol. Sci. 2019, 20, 3683. [Google Scholar] [CrossRef]
- Taddei, M.L.; Pardella, E.; Pranzini, E.; Raugei, G.; Paoli, P. Role of tyrosine phosphorylation in modulating cancer cell metabolism. Biochim. Biophys. Acta. Rev. Cancer 2020, 1874, 188442. [Google Scholar] [CrossRef]
- Du, X.; Zhang, J.; Zhang, X.; Schramm, K.W.; Nan, B.; Huang, Q.; Tian, M.; Shen, H. Persistence and reversibility of arsenic-induced gut microbiome and metabolome shifts in male rats after 30-days recovery duration. Sci. Total Environ. 2021, 776, 145972. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Ball, R.O.; Filler, R.M.; Moore, A.M.; Pencharz, P.B. Phenylalanine and tyrosine metabolism in neonates receiving parenteral nutrition differing in pattern of amino acids. Pediatr. Res. 1998, 44, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Min, X.; Zhu, T.; Hu, X.; Hou, C.; He, J.; Liu, X. Transcriptome and Metabolome Analysis of Isoquinoline Alkaloid Biosynthesis of Coptis chinensis in Different Years. Genes 2023, 14, 2232. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Batch No. | Location | Source |
|---|---|---|---|
| 1 | HS-1 | Zhejiang | Origin |
| 2 | HS-2 | Shandong | TCM market |
| 3 | HS-3 | Shandong | TCM market |
| 4 | HS-4 | Zhejiang | TCM market |
| 5 | HS-5 | Shandong | TCM market |
| 6 | HS-6 | Zhejiang | TCM market |
| 7 | HS-7 | Zhejiang | TCM market |
| 8 | HS-8 | Shandong | TCM market |
| 9 | HS-9 | Shandong | TCM market |
| 10 | HS-10 | Shandong | TCM market |
| 11 | LS-1 | Shandong | Origin |
| 12 | LS-2 | Zhejiang | Origin |
| 13 | LS-3 | Zhejiang | Origin |
| 14 | LS-4 | Zhejiang | Origin |
| 15 | LS-5 | Zhejiang | Origin |
| 16 | LS-6 | Shandong | Origin |
| 17 | LS-7 | Zhejiang | TCM market |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.-s.; Gong, S.; Xie, S.-m.; Chen, A.-z.; Jin, H.-y.; Liu, J.; Wang, Q.; Kang, S.; Li, P.; Wei, F.; et al. Mass Spectrometry-Based Metabolomics Investigation on Two Different Seaweeds Under Arsenic Exposure. Foods 2024, 13, 4055. https://doi.org/10.3390/foods13244055
Guo Y-s, Gong S, Xie S-m, Chen A-z, Jin H-y, Liu J, Wang Q, Kang S, Li P, Wei F, et al. Mass Spectrometry-Based Metabolomics Investigation on Two Different Seaweeds Under Arsenic Exposure. Foods. 2024; 13(24):4055. https://doi.org/10.3390/foods13244055
Chicago/Turabian StyleGuo, Yuan-sheng, Shuo Gong, Si-min Xie, An-zhen Chen, Hong-yu Jin, Jing Liu, Qi Wang, Shuai Kang, Ping Li, Feng Wei, and et al. 2024. "Mass Spectrometry-Based Metabolomics Investigation on Two Different Seaweeds Under Arsenic Exposure" Foods 13, no. 24: 4055. https://doi.org/10.3390/foods13244055
APA StyleGuo, Y.-s., Gong, S., Xie, S.-m., Chen, A.-z., Jin, H.-y., Liu, J., Wang, Q., Kang, S., Li, P., Wei, F., Zuo, T.-t., & Ma, S.-c. (2024). Mass Spectrometry-Based Metabolomics Investigation on Two Different Seaweeds Under Arsenic Exposure. Foods, 13(24), 4055. https://doi.org/10.3390/foods13244055