
Investigation of Efficient Pullulan Synthesis Utilizing Huangjiu Lees as a Substrate


Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Media
2.2. Pre-Treatment of Raw Materials for Huangjiu Lees
2.3. Cultivation of A. pullulans
2.4. Determination of the Yield and Molecular Weight of Pullulan
2.5. Adaptive Evolution
2.6. Transcriptome Sequencing and Analysis
2.7. Fermentation Process Optimization
2.8. Primer Design and Gene Expression Verification
2.9. Statistical Analysis
3. Results and Discussion
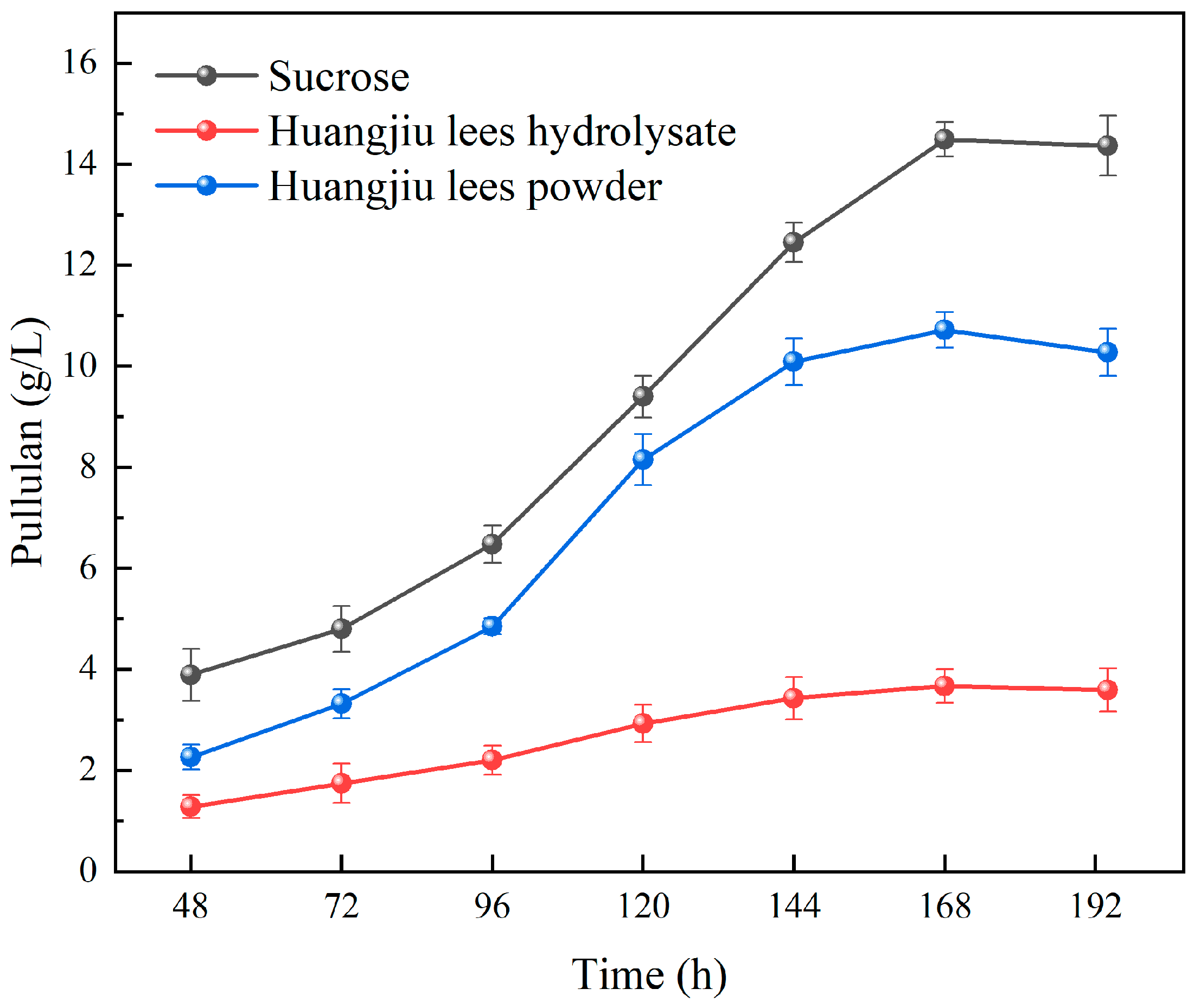
3.1. Effects of Different Pretreatment Methods of Huangjiu Lees on Pullulan Biosynthesis
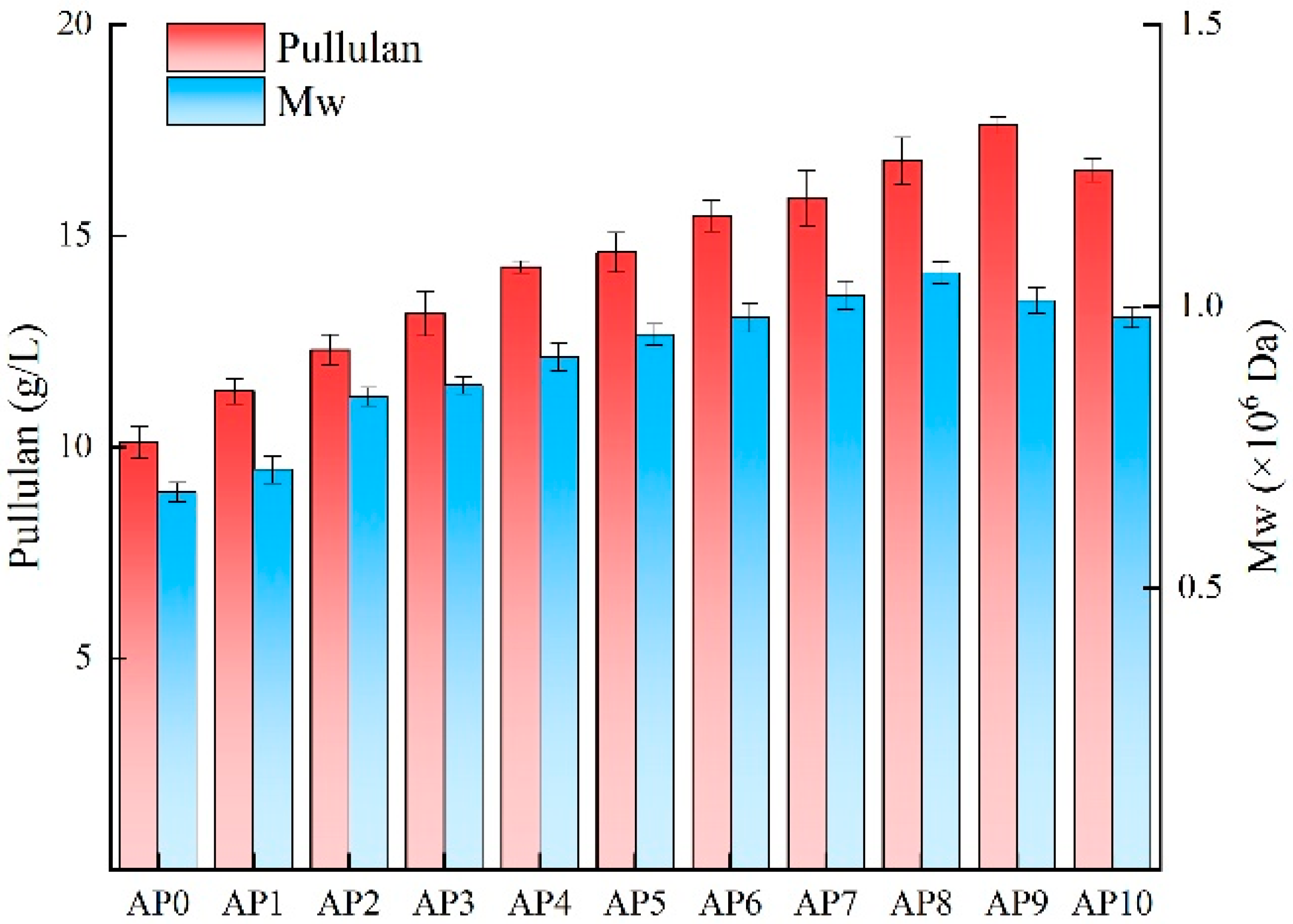
3.2. Adaptive Evolution of A. pullulans
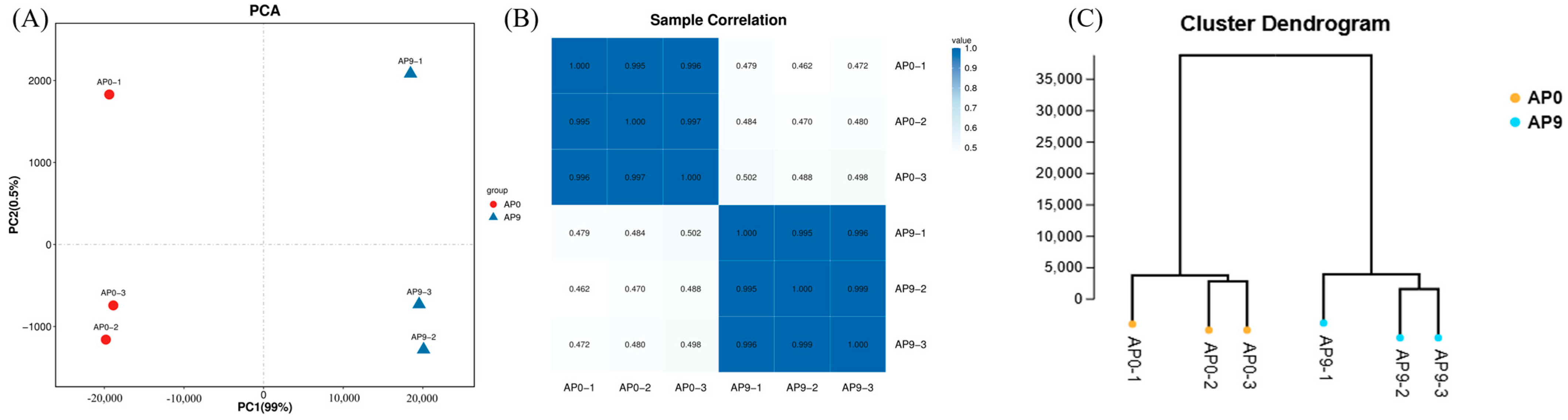
3.3. Transcriptome Sequencing Gene Statistics and Sample Relationships
3.4. Gene Expression Analysis
3.5. KEGG Enriched Pathway Analysis
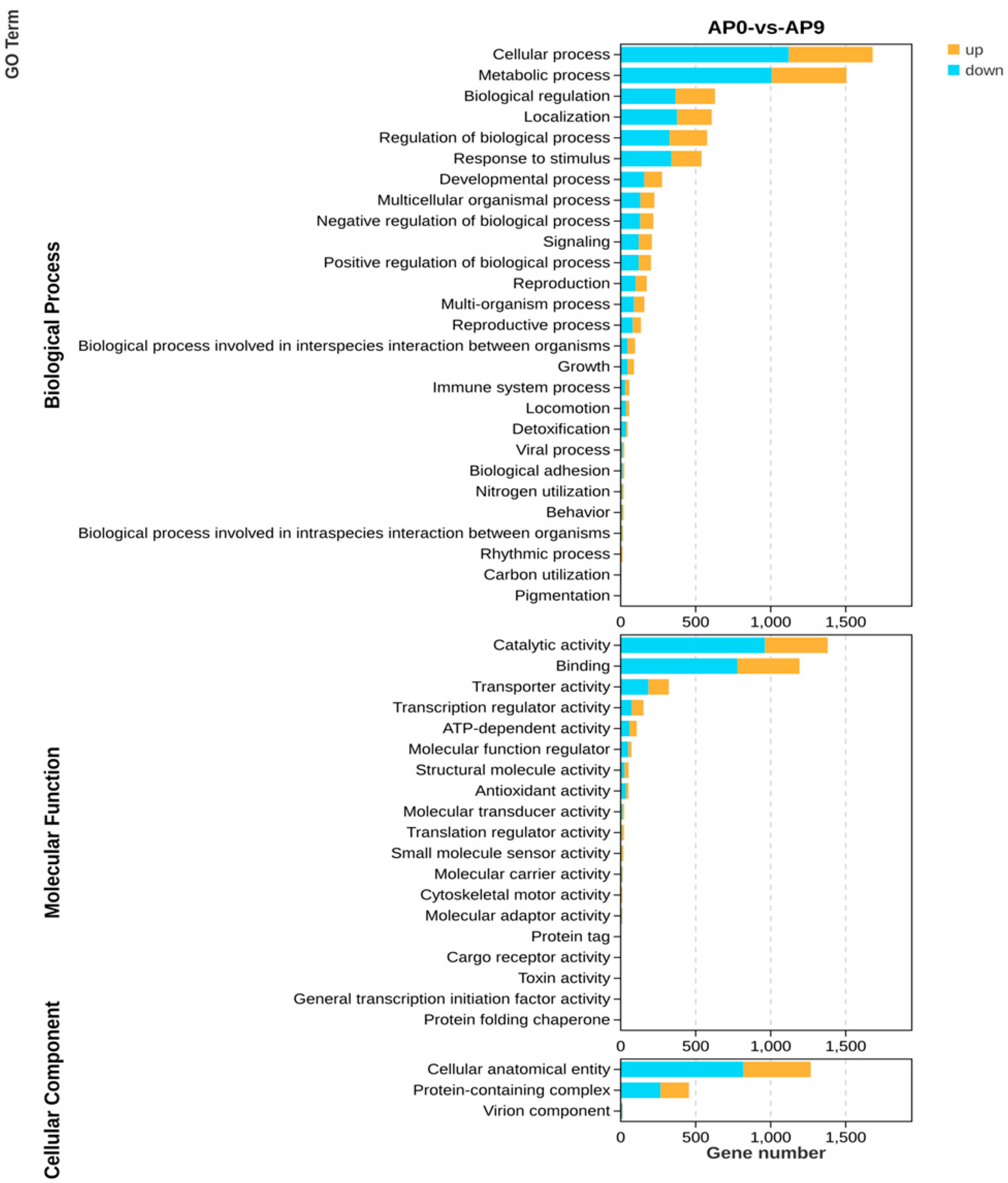
3.6. GO Enrichment Pathway Analysis
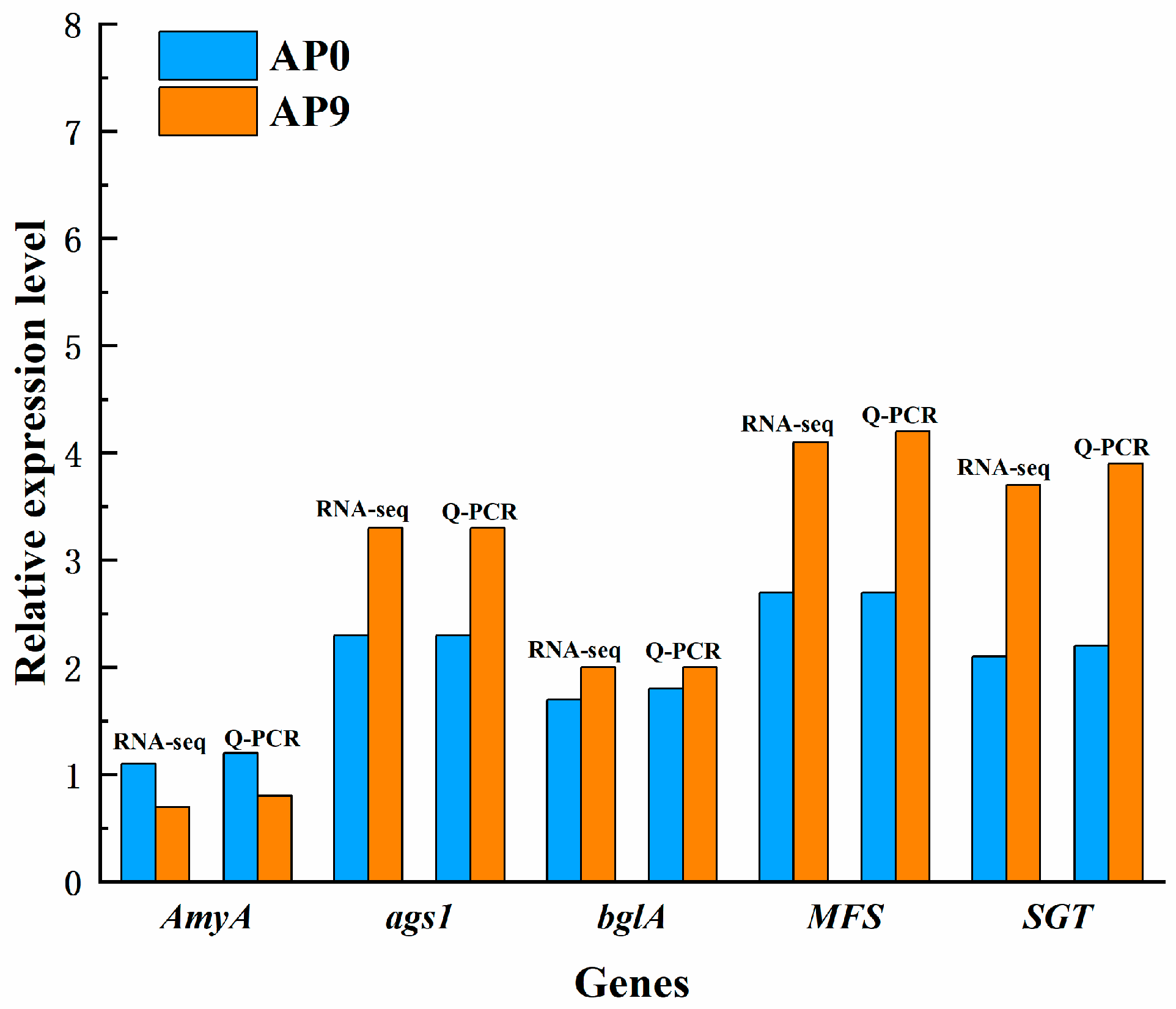
3.7. Validation of Key Expressed Genes
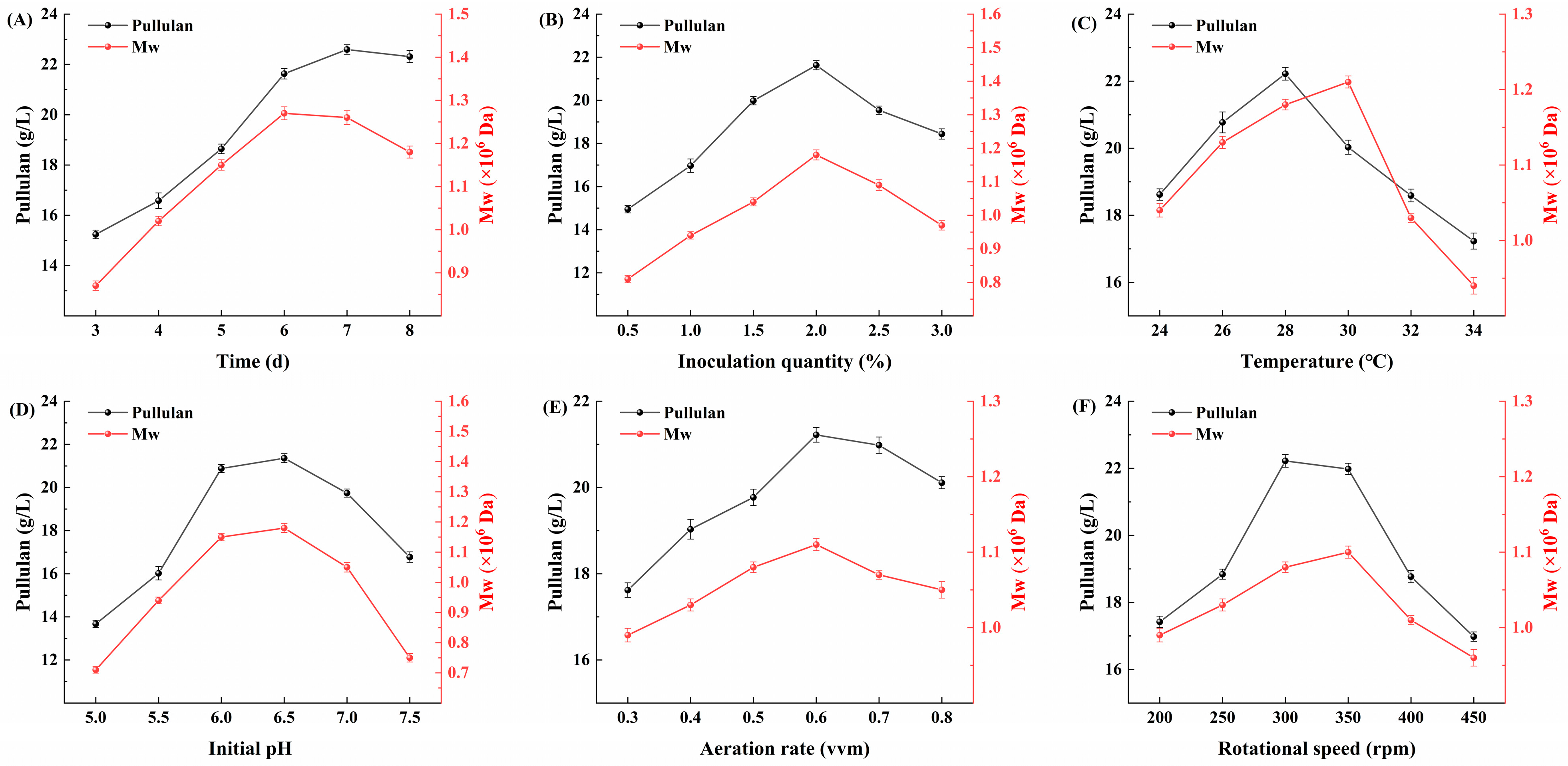
3.8. Optimization of Fermentation Process of Evolved Strains
3.9. Optimization of Staged Fermentation Strategy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hu, Y.; Pan, L.; Dun, Y.; Peng, N.; Liang, Y.; Zhao, S. Conversion of yellow wine lees into high-protein yeast culture by solid-state fermentation. Biotechnol. Biotechnol. Equip. 2014, 28, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.Y.; Wei, Z.H.; Xie, Y.Y.; Wang, D.M.; Liu, H.Y.; Fang, D.; Ma, M.R.; Liu, J.X. Lactation performance and nitrogen utilization of dairy cows on diets including unfermented or fermented yellow wine lees mix. Livest. Sci. 2020, 236, 104025. [Google Scholar] [CrossRef]
- Fabian, H.T.; Catalina, G.E. Characterization and chemical modification of pullulan produced from a submerged culture of Aureobasidium pullulans ATCC 15233. Polym. Test. 2022, 114, 107686. [Google Scholar]
- Guo, J.; Reis, J.; Salze, G.; Rhodes, M.; Tilton, S.; Davis, D.A. Using high protein distiller’s dried grain product to replace corn protein concentrate and fishmeal in practical diets for the Pacific white shrimp Litopenaeus vannamei. J. World Aquac. Soc. 2019, 50, 983–992. [Google Scholar] [CrossRef]
- Yao, K.; Xia, Q.; Cao, Y.; Chen, B.; Cai, J.; Liu, C. Feeding effect of yellow wine lees fermented with Candida utilis and Bacillus subtilis in the cow diet on milk composition. Food Biosci. 2023, 55, 103005. [Google Scholar] [CrossRef]
- Li, C.; Kong, D.; Yao, X.; Ma, X.; Wei, C.; Wang, H. Resource Recycling Utilization of Distillers Grains for Preparing Cationic Quaternary Ammonium—Ammonium Material and Adsorption of Acid Yellow 11. Sustainability 2022, 14, 2469. [Google Scholar] [CrossRef]
- Sharma, N.; Prasad, G.S.; Choudhury, A.R. Utilization of corn steep liquor for biosynthesis of pullulan, an important exopolysaccharide. Carbohydr. Polym. 2013, 93, 95–101. [Google Scholar] [CrossRef]
- Xia, Z.; Wu, S.; Pan, S. Effect of two-stage controlled pH and temperature on pullulan production by Auerobasidium pullulans. Carbohydr. Polym. 2011, 86, 1814–1816. [Google Scholar] [CrossRef]
- Duan, X.; Chi, Z.; Wang, L.; Wang, X. Influence of different sugars on pullulan production and activities of α-phosphoglucose mutase, UDPG-pyrophosphorylase and glucosyltransferase involved in pullulan synthesis in Aureobasidium pullulans Y68. Carbohydr. Polym. 2008, 73, 587–593. [Google Scholar] [CrossRef]
- Shingel, K.I. Current knowledge on biosynthesis, biological activity, and chemical modification of the exopolysaccharide, pullulan. Carbohydr. Res. 2004, 339, 447–460. [Google Scholar] [CrossRef]
- West, T.P. Impact of Aureobasidium Species Strain Improvement on the Production of the Polysaccharide Pullulan. Polysaccharides 2024, 5, 305–319. [Google Scholar] [CrossRef]
- Kang, C.; Hao, L.; Zhang, L.; Zheng, Z.; Yang, Y. Isolation, purification and antioxidant activity of polysaccharides from the leaves of maca (Lepidium meyenii). Int. J. Biol. Macromol. 2018, 107, 2611–2619. [Google Scholar]
- Wang, W.; Zhang, K.; Lin, C.; Zhao, S.; Guan, J.; Zhou, W.; Ru, X.; Cong, H.; Yang, Q. Influence of Cmr1 in the Regulation of Antioxidant Function Melanin Biosynthesis in Aureobasidium pullulans. Foods 2023, 12, 2135. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ji, F.; Bao, Y.; Xia, J.; Guo, L.; Wang, J.; Li, Y. Biocompatible cationic pullulan-g-desoxycholic acid-g-PEI micelles used to co-deliver drug and gene for cancer therapy. Mater. Sci. Eng. C 2017, 70, 418–429. [Google Scholar] [CrossRef]
- Mayra, S.; Pau, T.; Amparo, C. Improving Functional Properties of Cassava Starch-Based Films by Incorporating Xanthan, Gellan, or Pullulan Gums. Int. J. Polym. Sci. 2019, 2019, 5367164. [Google Scholar]
- María, C.M.; Fernandes, A.F.A.; Leda, d.A.G.; Pereira, S.V.; Aline, P.C.; Nayeli, O.; José, C.M.; Franco, M.P.R.; César, S.J. Production and applications of pullulan from lignocellulosic biomass: Challenges and perspectives. Bioresour. Technol. 2023, 385, 129460. [Google Scholar]
- Cheng, K.C.; Demirci, A.; Catchmark, J.M. Pullulan: Biosynthesis, production, and applications. Appl. Microbiol. Biotechnol. 2011, 92, 29–44. [Google Scholar] [CrossRef]
- Wang, D.; Ju, X.; Zhou, D.; Wei, G. Efficient production of pullulan using rice hull hydrolysate by adaptive laboratory evolution of Aureobasidium pullulans. Bioresour. Technol. 2014, 164, 12–19. [Google Scholar] [CrossRef]
- Herrera, A.; Téllez-Luis, S.J.; Ramıírez, J.A.; Vázquez, M. Production of Xylose from Sorghum Straw Using Hydrochloric Acid. J. Cereal Sci. 2003, 37, 267–274. [Google Scholar] [CrossRef]
- Lavarack, B.P.; Griffin, G.J.; Rodman, D. The acid hydrolysis of sugarcane bagasse hemicellulose to produce xylose, arabinose, glucose and other products. Biomass Bioenergy 2002, 23, 367–380. [Google Scholar] [CrossRef]
- Lazaridou, A.; Roukas, T.; Biliaderis, C.G.; Vaikousi, H. Characterization of pullulan produced from beet molasses by Aureobasidium pullulans in a stirred tank reactor under varying agitation. Enzym. Microb. Technol. 2002, 31, 122–132. [Google Scholar] [CrossRef]
- Tagne, R.F.T.; Santos, M.M.C.; Antunes, F.A.F.; Shibukawa, V.P.; Miano, S.B.; Kenfack, J.A.A.; Silva, S.S.d.; Ngomade, S.B.L.; Santos, J.C. Pullulan Production from Sugarcane Bagasse Hemicellulosic Hydrolysate by Aureobasidium pullulans ATCC 42023 inBubble Column Reactor. Fermentation 2024, 10, 322. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, W.; Yang, Q. Transcriptome Analysis Reveals the Regulation of Aureobasidium pullulans under Different pH Stress. Int. J. Mol. Sci. 2023, 24, 16103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Lin, C.; Zhao, S.; Wang, W.; Zhou, W.; Ru, X.; Cong, H.; Yang, Q. The role of pH transcription factor Appacc in upregulation of pullulan biosynthesis in Aureobasidium pullulans using potato waste as a substrate. Int. J. Biol. Macromol. 2023, 242, 124797. [Google Scholar] [CrossRef]
- Wang, Q.; Liang, L.; Xi, F.; Tian, G.; Mao, Q.; Meng, X. Adsorption of Azo Dye Acid Red 73 onto Rice Wine Lees: Adsorption Kinetics and Isotherms. Adv. Mater. Sci. Eng. 2020, 2020, 3469579. [Google Scholar] [CrossRef]
- He, C.; Zhang, X.; Zhang, Z.; Wang, C.; Wang, D.; Wei, G. Whole-crop biorefinery of corn biomass for pullulan production by Aureobasidium pullulans. Bioresour. Technol. 2022, 370, 128517. [Google Scholar] [CrossRef]
- Liu, N.-N.; Chi, Z.; Wang, Q.-Q.; Hong, J.; Liu, G.-L.; Hu, Z.; Chi, Z.-M. Simultaneous production of both high molecular weight pullulan and oligosaccharides by Aureobasdium melanogenum P16 isolated from a mangrove ecosystem. Int. J. Biol. Macromol. 2017, 102, 1016–1024. [Google Scholar] [CrossRef]
- Cheng, K.-C. Evaluation of Medium Composition and Fermentation Parameters on Pullulan Production by Aureobasidium pullulans. Food Sci. Technol. Int. 2011, 17, 99–109. [Google Scholar] [CrossRef]
- Jiang, B.; Liu, J.; Wang, J.; Zhao, G.; Zhao, Z. Adaptive Evolution for the Efficient Production of High-Quality d-Lactic Acid Using Engineered Klebsiella pneumoniae. Microorganisms 2024, 12, 1167. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, J.; Wang, P.; Xia, J.; Li, X.; Zou, X. CRISPR/Cas9-mediated efficient genome editing via protoplast-based transformation in yeast-like fungus Aureobasidium pullulans. Gene 2019, 709, 8–16. [Google Scholar] [CrossRef]
- Gostinčar, C.; Ohm, R.A.; Kogej, T.; Sonjak, S.; Turk, M.; Zajc, J.; Zalar, P.; Grube, M.; Sun, H.; Han, J.; et al. Genome sequencing of four Aureobasidium pullulans varieties: Biotechnological potential, stress tolerance, and description of new species. BMC Genom. 2014, 15, 549. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.N.; Chi, Z.; Liu, G.L.; Chen, T.J.; Jiang, H.; Hu, Z.; Chi, Z.M. α-Amylase, glucoamylase and isopullulanase determine molecular weight of pullulan produced by Aureobasidium melanogenum P16. Int. J. Biol. Macromol. 2018, 117, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Chen, L.; Zhang, L.; Zou, G.; Liu, R.; Jiang, Y.; Zhou, Z. RNA Sequencing Reveals Xyr1 as a Transcription Factor Regulating Gene Expression beyond Carbohydrate Metabolism. BioMed Res. Int. 2016, 2016, 4841756. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-J.; Liu, G.-L.; Chen, L.; Yang, G.; Hu, Z.; Chi, Z.-M.; Chi, Z. Alternative primers are required for pullulan biosynthesis in Aureobasidium melanogenum P16. Int. J. Biol. Macromol. 2020, 147, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Aquinas, N.; Chithra, C.H.; Bhat, M.R. Progress in bioproduction, characterization and applications of pullulan: A review. Polym. Bull. 2024, 81, 12347–12382. [Google Scholar] [CrossRef]
- Wani, S.M.; Mir, S.A.; Khanday, F.A.; Masoodi, F.A. Advances in pullulan production from agro-based wastes by Aureobasidium pullulans and its applications. Innov. Food Sci. Emerg. Technol. 2021, 74, 102846. [Google Scholar] [CrossRef]
- Xu, Y.; Ye, D.; Zhang, W.; Wang, Y.; Li, J.; Zhang, L.; Huang, J.; Zhu, X.; Liao, Q. Dual-scale pore network modeling of two-phase transport in anode porous transport layer and catalyst layer of proton exchange membrane electrolyzers. Energy Convers. Manag. 2024, 322, 119089. [Google Scholar] [CrossRef]
- Shu, L.; Yang, M.; Zhao, H.; Li, T.; Yang, L.; Zou, X.; Li, Y. Process optimization in a stirred tank bioreactor based on CFD-Taguchi method: A case study. J. Clean. Prod. 2019, 230, 1074–1084. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, K.; Zhao, S.; Wang, W.; Ru, X.; Song, J.; Cong, H.; Yang, Q. Screening and identification of a strain of Aureobasidium pullulans and its application in potato starch industrial waste. Environ. Res. 2022, 214, 113947. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Single Factor | Variable |
|---|---|
| Fermentation time (d) | 3, 4, 5, 6, 7, 8 |
| Initial pH | 5.0, 5.5, 6.0, 6.5, 7.0, 7.5 |
| Inoculation quantity (%) | 0.5, 1.0, 1.5, 2.0, 2.5, 3.0 |
| Aeration rate (vvm) | 0.3, 0.4, 0.5, 0.6, 0.7, 0.8 |
| Rotational speed (rpm) | 200, 250, 300, 350, 400, 450 |
| Temperature (°C) | 24, 26, 28, 30, 32, 34 |
| Influencing Factors | Variable Range | Other Parameters |
|---|---|---|
| Aspergillus niger inoculation quantity (%) | 1, 2, 3, 4, 5, 6 | pH 6.5, 5 d, 28 °C, 300 rpm, 0.6 vvm |
| Aspergillus niger inoculation time (d) | 0.5, 1.0, 1.5, 2.0, 2.5, 3.0 | pH 6.5, 1.0% inoculation quantity, 28 °C, 300 rpm, 0.6 vvm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, P.; Liu, T.; Ma, J.; Kan, T.; Han, X.; Ji, Z.; Mao, J. Investigation of Efficient Pullulan Synthesis Utilizing Huangjiu Lees as a Substrate. Foods 2024, 13, 3874. https://doi.org/10.3390/foods13233874
Lu P, Liu T, Ma J, Kan T, Han X, Ji Z, Mao J. Investigation of Efficient Pullulan Synthesis Utilizing Huangjiu Lees as a Substrate. Foods. 2024; 13(23):3874. https://doi.org/10.3390/foods13233874
Chicago/Turabian StyleLu, Peiqi, Tiantian Liu, Jingqiu Ma, Tao Kan, Xiao Han, Zhongwei Ji, and Jian Mao. 2024. "Investigation of Efficient Pullulan Synthesis Utilizing Huangjiu Lees as a Substrate" Foods 13, no. 23: 3874. https://doi.org/10.3390/foods13233874
APA StyleLu, P., Liu, T., Ma, J., Kan, T., Han, X., Ji, Z., & Mao, J. (2024). Investigation of Efficient Pullulan Synthesis Utilizing Huangjiu Lees as a Substrate. Foods, 13(23), 3874. https://doi.org/10.3390/foods13233874