

From Food Waste to Functional Biopolymers: Characterization of Chitin and Chitosan Produced from Prepupae of Black Soldier Fly Reared with Different Food Waste-Based Diets
 , , ,
, , ,  ,
, 
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Hermetia illucens Rearing
2.2. Hermetia illucens Prepupae Production on Different Diets
2.3. Evaluation of Hermetia illucens Larval Mortality
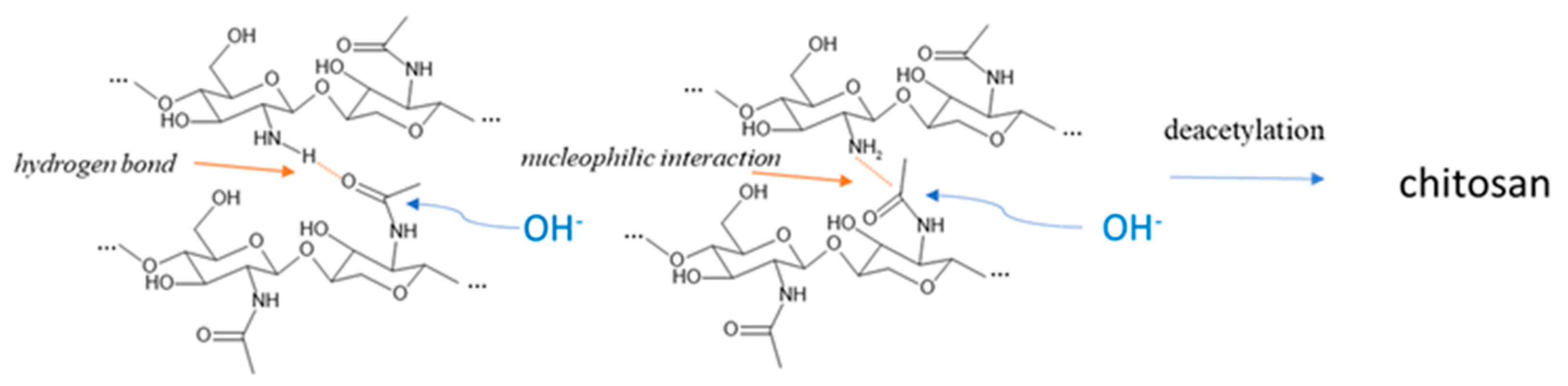
2.4. Chitin Extraction and Chitosan Production from Hermetia illucens Prepupae
2.5. Chitin and Chitosan Characterization
2.5.1. ATR-IR Characterization
2.5.2. SEM Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Effect of Diets on H. illucens Larval Mortality
3.2. Influence of the Diet on Biomass Recovery
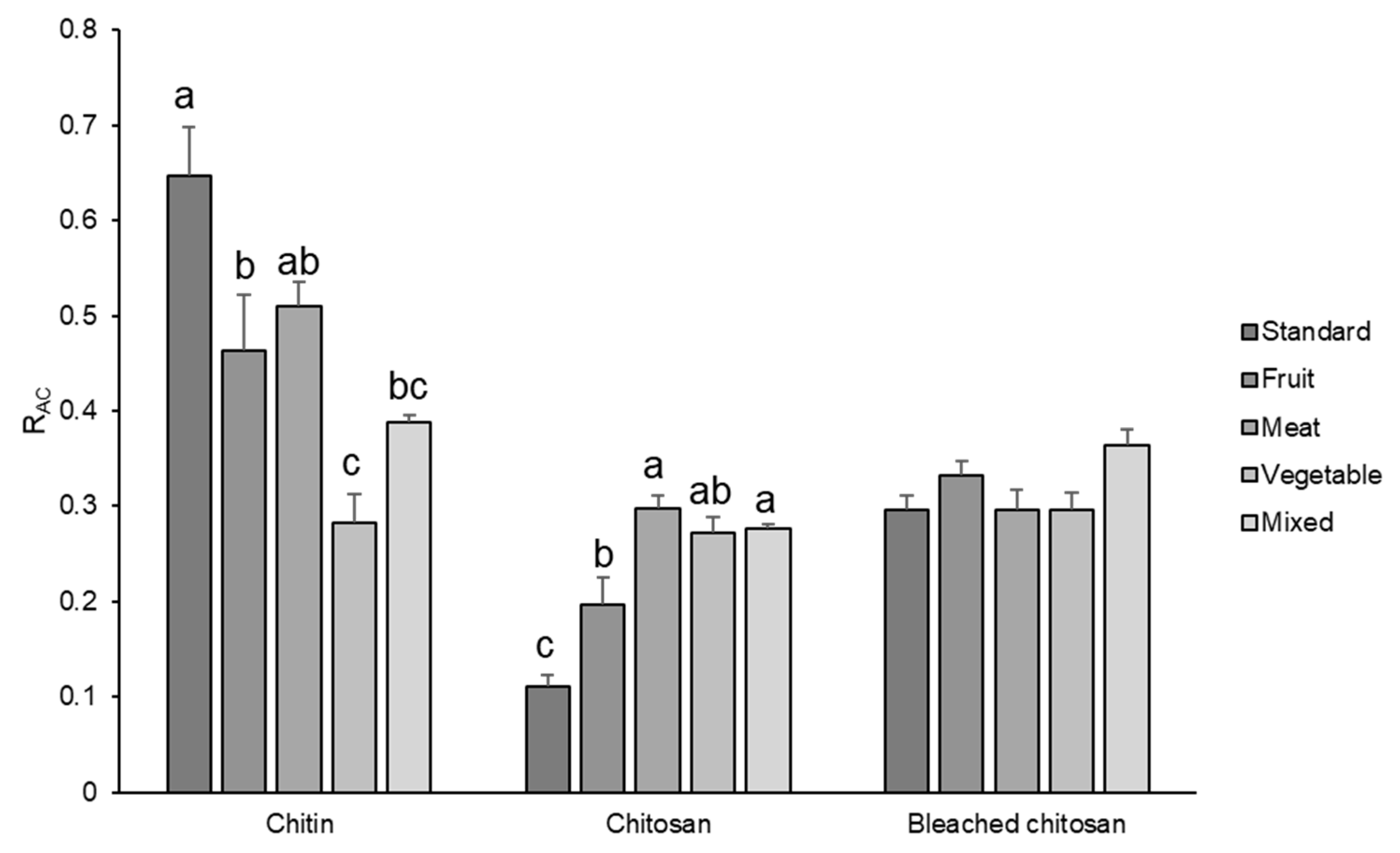
3.3. ATR-IR Analysis
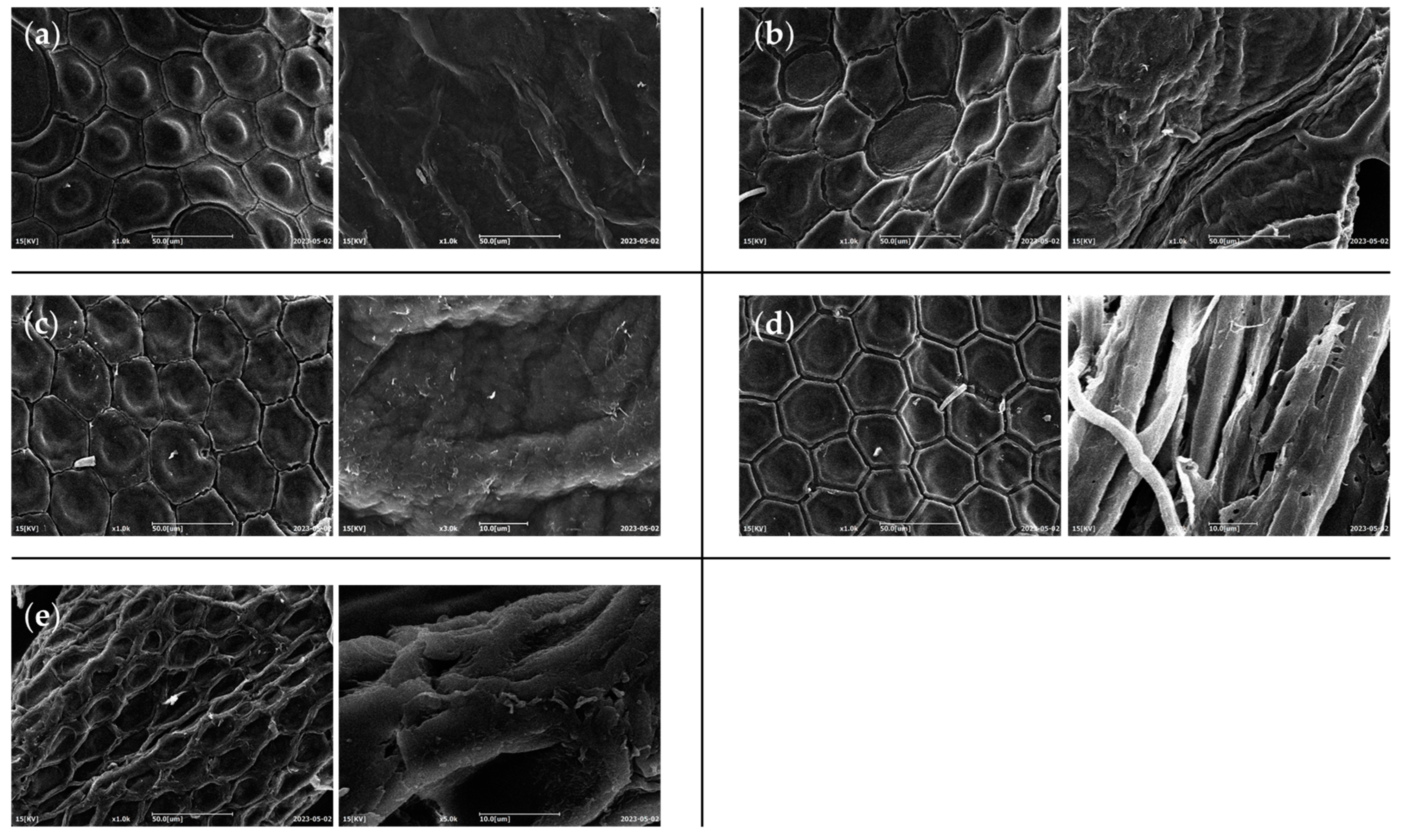
3.4. SEM Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fowles, T.M.; Nansen, C. Insect-Based Bioconversion: Value from Food Waste. In Food Waste Management: Solving the Wicked Problem; Springer International Publishing: Cham, Switzerland, 2019; pp. 321–346. ISBN 9783030205614. [Google Scholar]
- Finke, M.D. Complete Nutrient Content of Four Species of Feeder Insects. Zoo Biol. 2013, 32, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Triunfo, M.; Tafi, E.; Guarnieri, A.; Salvia, R.; Scieuzo, C.; Hahn, T.; Zibek, S.; Gagliardini, A.; Panariello, L.; Coltelli, M.B.; et al. Characterization of Chitin and Chitosan Derived from Hermetia Illucens, a Further Step in a Circular Economy Process. Sci. Rep. 2022, 12, 6613. [Google Scholar] [CrossRef] [PubMed]
- Hahn, T.; Tafi, E.; von Seggern, N.; Falabella, P.; Salvia, R.; Thomä, J.; Febel, E.; Fijalkowska, M.; Schmitt, E.; Stegbauer, L.; et al. Purification of Chitin from Pupal Exuviae of the Black Soldier Fly. Waste Biomass Valorization 2022, 13, 1993–2008. [Google Scholar] [CrossRef]
- Soetemans, L.; Uyttebroek, M.; Bastiaens, L. Characteristics of Chitin Extracted from Black Soldier Fly in Different Life Stages. Int. J. Biol. Macromol. 2020, 165, 3206–3214. [Google Scholar] [CrossRef]
- Mincea, M.; Negrulescu, A.; Ostafe, V. Preparation, Modification, and Applications Of chitin Nanowhiskers: A Review. Rev. Adv. Mater. Sci. 2012, 30, 225–242. [Google Scholar]
- Abidin, N.A.Z.; Kormin, F.; Abidin, N.A.Z.; Anuar, N.A.F.M.; Bakar, M.F.A. The Potential of Insects as Alternative Sources of Chitin: An Overview on the Chemical Method of Extraction from Various Sources. Int. J. Mol. Sci. 2020, 21, 4978. [Google Scholar] [CrossRef]
- Jantzen da Silva Lucas, A.; Quadro Oreste, E.; Leão Gouveia Costa, H.; Martín López, H.; Dias Medeiros Saad, C.; Prentice, C. Extraction, Physicochemical Characterization, and Morphological Properties of Chitin and Chitosan from Cuticles of Edible Insects. Food Chem. 2021, 343, 128550. [Google Scholar] [CrossRef]
- Marei, N.H.; El-Samie, E.A.; Salah, T.; Saad, G.R.; Elwahy, A.H.M. Isolation and Characterization of Chitosan from Different Local Insects in Egypt. Int. J. Biol. Macromol. 2016, 82, 871–877. [Google Scholar] [CrossRef]
- Poerio, A.; Petit, C.; Jehl, J.P.; Arab-Tehrany, E.; Mano, J.F.; Cleymand, F. Extraction and Physicochemical Characterization of Chitin from Cicada Orni Sloughs of the South-Eastern French Mediterranean Basin. Molecules 2020, 25, 2543. [Google Scholar] [CrossRef]
- Mohan, K.; Ganesan, A.R.; Muralisankar, T.; Jayakumar, R.; Sathishkumar, P.; Uthayakumar, V.; Chandirasekar, R.; Revathi, N. Recent Insights into the Extraction, Characterization, and Bioactivities of Chitin and Chitosan from Insects. Trends Food Sci. Technol. 2020, 105, 17–42. [Google Scholar] [CrossRef]
- Aranaz, I.; Alcántara, A.R.; Civera, M.C.; Arias, C.; Elorza, B.; Caballero, A.H.; Acosta, N. Chitosan: An Overview of Its Properties and Applications. Polymers 2021, 13, 3256. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wang, D.; Liu, D.; Su, J.; Jin, Y.; Wang, D.; Han, B.; Jiang, Z.; Liu, B. Applications of Chitosan and Its Derivatives in Skin and Soft Tissue Diseases. Front. Bioeng. Biotechnol. 2022, 10, 894667. [Google Scholar]
- Pichyangkura, R.; Chadchawan, S. Biostimulant Activity of Chitosan in Horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, F.; Li, C.; An, H.; Wan, T.; Zhang, P. Application of Chitosan and Its Derivative Polymers in Clinical Medicine and Agriculture. Polymers 2022, 14, 958. [Google Scholar] [CrossRef]
- Abenaim, L.; Conti, B. Chitosan as a Control Tool for Insect Pest Management: A Review. Insects 2023, 14, 949. [Google Scholar] [CrossRef] [PubMed]
- No, H.K.; Meyers, S.P.; Prinyawiwatkul, W.; Xu, Z. Applications of Chitosan for Improvement of Quality and Shelf Life of Foods: A Review. J. Food Sci. 2007, 72, R87–R100. [Google Scholar] [CrossRef]
- Mujtaba, M.; Morsi, R.E.; Kerch, G.; Elsabee, M.Z.; Kaya, M.; Labidi, J.; Khawar, K.M. Current Advancements in Chitosan-Based Film Production for Food Technology; A Review. Int. J. Biol. Macromol. 2019, 121, 889–904. [Google Scholar] [CrossRef] [PubMed]
- Farina, P.; Ascrizzi, R.; Bedini, S.; Castagna, A.; Flamini, G.; Macaluso, M.; Mannucci, A.; Pieracci, Y.; Ranieri, A.; Sciampagna, M.C.; et al. Chitosan and Essential Oils Combined for Beef Meat Protection against the Oviposition of Calliphora Vomitoria, Water Loss, Lipid Peroxidation, and Colour Changes. Foods 2022, 11, 3994. [Google Scholar] [CrossRef]
- Flórez, M.; Guerra-Rodríguez, E.; Cazón, P.; Vázquez, M. Chitosan for Food Packaging: Recent Advances in Active and Intelligent Films. Food Hydrocoll. 2022, 124, 107328. [Google Scholar] [CrossRef]
- Hahn, T.; Roth, A.; Ji, R.; Schmitt, E.; Zibek, S. Chitosan Production with Larval Exoskeletons Derived from the Insect Protein Production. J. Biotechnol. 2020, 310, 62–67. [Google Scholar] [CrossRef]
- Coltelli, M.B.; Cinelli, P.; Gigante, V.; Aliotta, L.; Morganti, P.; Panariello, L.; Lazzeri, A. Chitin Nanofibrils in Poly(Lactic Acid) (PLA) Nanocomposites: Dispersion and Thermo-Mechanical Properties. Int. J. Mol. Sci. 2019, 20, 504. [Google Scholar] [CrossRef] [PubMed]
- Panariello, L.; Coltelli, M.B.; Hadrich, A.; Braca, F.; Fiori, S.; Haviv, A.; Miketa, F.; Lazzeri, A.; Staebler, A.; Gigante, V.; et al. Antimicrobial and Gas Barrier Crustaceans and Fungal Chitin-Based Coatings on Biodegradable Bioplastic Films. Polymers 2022, 14, 5211. [Google Scholar] [CrossRef]
- Coltelli, M.B.; Aliotta, L.; Vannozzi, A.; Morganti, P.; Panariello, L.; Danti, S.; Neri, S.; Fernandez-Avila, C.; Fusco, A.; Donnarumma, G.; et al. Properties and Skin Compatibility of Films Based on Poly(Lactic Acid) (PLA) Bionanocomposites Incorporating Chitin Nanofibrils (CN). J. Funct. Biomater. 2020, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Coltelli, M.B.; Morganti, P.; Castelvetro, V.; Lazzeri, A.; Danti, S.; Benjelloun-Mlayah, B.; Gagliardini, A.; Fusco, A.; Donnarumma, G. Chitin Nanofibril-Nanolignin Complexes as Carriers of Functional Molecules for Skin Contact Applications. Nanomaterials 2022, 12, 1295. [Google Scholar] [CrossRef]
- Panariello, L.; Coltelli, M.B.; Giangrandi, S.; Garrigós, M.C.; Hadrich, A.; Lazzeri, A.; Cinelli, P. Influence of Functional Bio-Based Coatings Including Chitin Nanofibrils or Polyphenols on Mechanical Properties of Paper Tissues. Polymers 2022, 14, 2274. [Google Scholar] [CrossRef] [PubMed]
- Eggink, K.M.; Lund, I.; Pedersen, P.B.; Hansen, B.W.; Dalsgaard, J. Biowaste and By-Products as Rearing Substrates for Black Soldier Fly (Hermetia Illucens) Larvae: Effects on Larval Body Composition and Performance. PLoS ONE 2022, 17, e0275213. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C. Lekking behavior of the black soldier fly (Diptera: Stratiomyidae). Fla. Èntomol. 2001, 84, 729. [Google Scholar] [CrossRef]
- Bosch, G.; Van Zanten, H.H.E.; Zamprogna, A.; Veenenbos, M.; Meijer, N.P.; Van der Fels-Klerx, H.J.; Van Loon, J.J.A. Conversion of organic resources by black soldier fly larvae: Legislation, efficiency and environmental impact. J. Clean. Prod. 2019, 222, 355–363. [Google Scholar] [CrossRef]
- Jucker, C.; Erba, D.; Leonardi, M.G.; Lupi, D.; Savoldelli, S. Assessment of Vegetable and Fruit Substrates as Potential Rearing Media for Hermetia Illucens (Diptera: Stratiomyidae) Larvae. Environ. Èntomol. 2017, 46, 1415–1423. [Google Scholar] [CrossRef]
- Julita, U.; Suryani, Y.; Kinasih, I.; Yuliawati, A.; Cahyanto, T.; Maryeti, Y.; Permana, A.D.; Fitri, L.L. Growth Performance and Nutritional Composition of Black Soldier Fly, Hermetia Illucens (L), (Diptera: Stratiomyidae) Reared on Horse and Sheep Manure. In Proceedings of the OP Conference Series: Earth and Environmental Science; Institute of Physics Publishing: Bristol, UK, 2018; Volume 187. [Google Scholar]
- Bava, L.; Jucker, C.; Gislon, G.; Lupi, D.; Savoldelli, S.; Zucali, M.; Colombini, S. Rearing of Hermetia Illucens on Different Organic By-Products: Influence on Growth, Waste Reduction, and Environmental Impact. Animals 2019, 9, 289. [Google Scholar] [CrossRef]
- Nafisah, A.; Nahrowi; Mutia, R.; Jayanegara, A. Chemical Composition, Chitin and Cell Wall Nitrogen Content of Black Soldier Fly (Hermetia Illucens) Larvae after Physical and Biological Treatment. IOP Conf. Series Mater. Sci. Eng. 2019, 546, 042028. [Google Scholar] [CrossRef]
- Henriques, B.S.; Garcia, E.S.; Azambuja, P.; Genta, F.A. Determination of Chitin Content in Insects: An Alternate Method Based on Calcofluor Staining. Front. Physiol. 2020, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Muthukrishnan, S.; Mun, S.; Noh, M.Y.; Geisbrecht, E.R.; Arakane, Y. Insect Cuticular Chitin Contributes to Form and Function. Curr. Pharm. Des. 2020, 26, 3530–3545. [Google Scholar] [CrossRef] [PubMed]
- Smets, R.; Claes, J.; Van Der Borght, M. On the Nitrogen Content and a Robust Nitrogen-to-Protein Conversion Factor of Black Soldier Fly Larvae (Hermetia Illucens). Anal. Bioanal. Chem. 2021, 413, 6365–6377. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of Rearing Substrate on Growth Performance, Waste Reduction Efficiency and Chemical Composition of Black Soldier Fly (Hermetia Illucens) Larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Deringer, V.L.; Englert, U.; Dronskowski, R. Nature, Strength, and Cooperativity of the Hydrogen-Bonding Network in α-Chitin. Biomacromolecules 2016, 17, 996–1003. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | Composition |
|---|---|
| Standard | Poultry feed and water (40:60) |
| Fruit | 75% apple, orange, banana (1:1:1) + 25% standard diet |
| Vegetable | 75% celery, sweet pepper, potatoes (1:1:1) + 25% standard diet |
| Meat | 75% poultry meat + 25% standard diet |
| Mixed | 25% for each previous diet (fruit, vegetable, meat and standard) (1:1:1:1) |
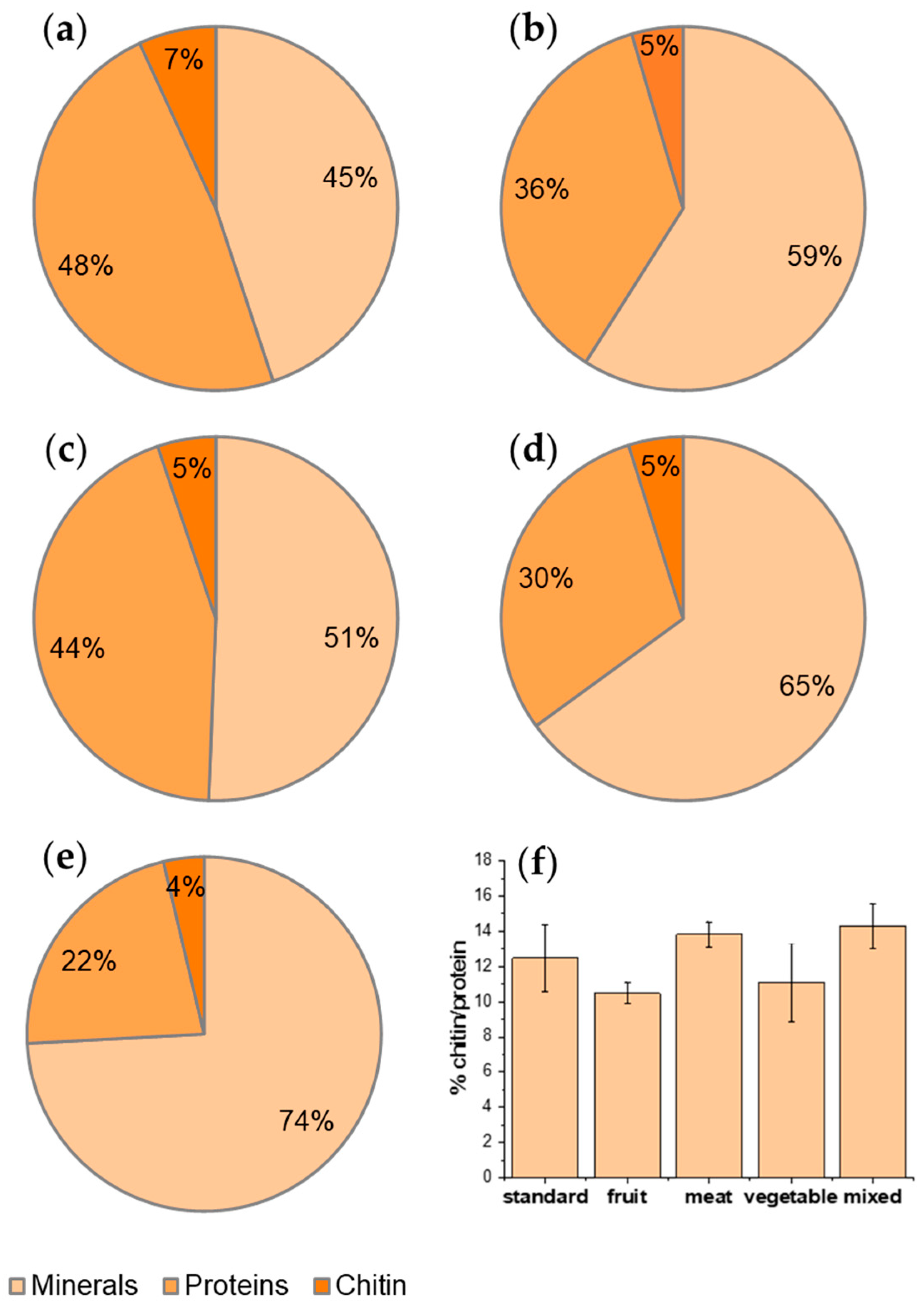
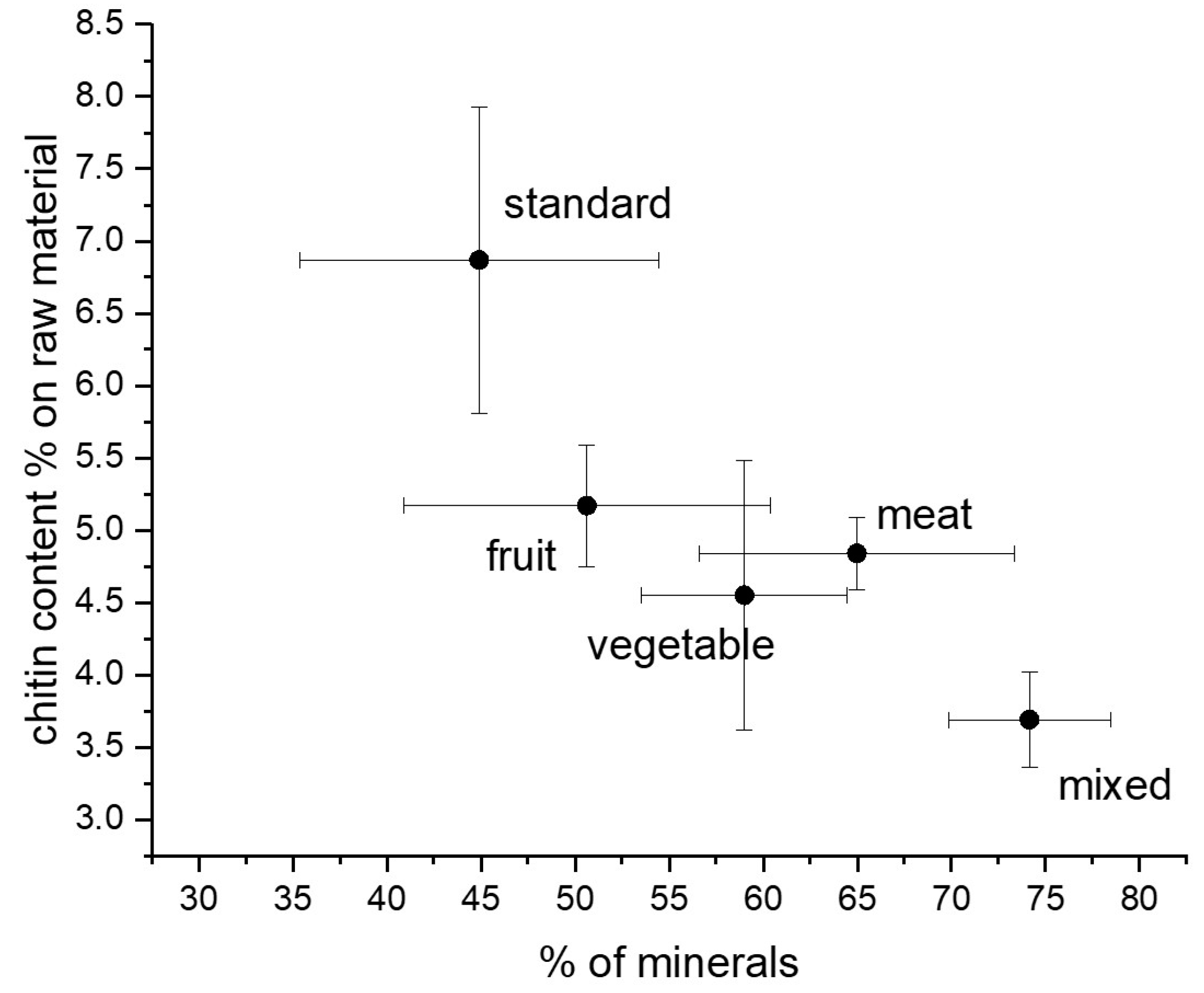
| DM/RAW (%) | DP/RAW (%) | CHT/RAW (%) | CHT/CHITIN (%) | Bleached CHT/CHT (%) | |
|---|---|---|---|---|---|
| Standard | 55.09 ± 11.53 | 6.87 ± 1.06 | 4.69 ± 0.92 | 67.22 ± 4.41 | 13.90 ± 3.19 |
| Fruit | 49.37 ± 11.73 | 5.17 ± 0.29 | 3.39 ± 0.42 | 65.24 ± 5.09 | 13.05 ± 0.02 |
| Meat | 35.02 ± 10.40 | 4.84 ± 0.25 | 3.30 ± 0.28 | 68.05 ± 2.19 | 12.64 ± 2.40 |
| Vegetable | 41.00 ± 5.47 | 4.55 ± 0.93 | 2.97 ± 0.67 | 64.64 ± 2.31 | 19.31 ± 8.20 |
| Mixed | 25.84 ± 4.28 | 3.69 ± 0.33 | 2.40 ± 0.32 | 64.91 ± 5.71 | 18.50 ± 6.81 |
| p value | 0.2577 | 0.0700 | 0.1476 | 0.9672 | 0.8124 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannucci, A.; Panariello, L.; Abenaim, L.; Coltelli, M.B.; Ranieri, A.; Conti, B.; Santin, M.; Castagna, A. From Food Waste to Functional Biopolymers: Characterization of Chitin and Chitosan Produced from Prepupae of Black Soldier Fly Reared with Different Food Waste-Based Diets. Foods 2024, 13, 278. https://doi.org/10.3390/foods13020278
Mannucci A, Panariello L, Abenaim L, Coltelli MB, Ranieri A, Conti B, Santin M, Castagna A. From Food Waste to Functional Biopolymers: Characterization of Chitin and Chitosan Produced from Prepupae of Black Soldier Fly Reared with Different Food Waste-Based Diets. Foods. 2024; 13(2):278. https://doi.org/10.3390/foods13020278
Chicago/Turabian StyleMannucci, Alessia, Luca Panariello, Linda Abenaim, Maria Beatrice Coltelli, Annamaria Ranieri, Barbara Conti, Marco Santin, and Antonella Castagna. 2024. "From Food Waste to Functional Biopolymers: Characterization of Chitin and Chitosan Produced from Prepupae of Black Soldier Fly Reared with Different Food Waste-Based Diets" Foods 13, no. 2: 278. https://doi.org/10.3390/foods13020278
APA StyleMannucci, A., Panariello, L., Abenaim, L., Coltelli, M. B., Ranieri, A., Conti, B., Santin, M., & Castagna, A. (2024). From Food Waste to Functional Biopolymers: Characterization of Chitin and Chitosan Produced from Prepupae of Black Soldier Fly Reared with Different Food Waste-Based Diets. Foods, 13(2), 278. https://doi.org/10.3390/foods13020278