

Identification of Antibacterial Components and Modes in the Methanol-Phase Extract from a Herbal Plant Potentilla kleiniana Wight et Arn

Abstract
1. Introduction
2. Results and Discussion
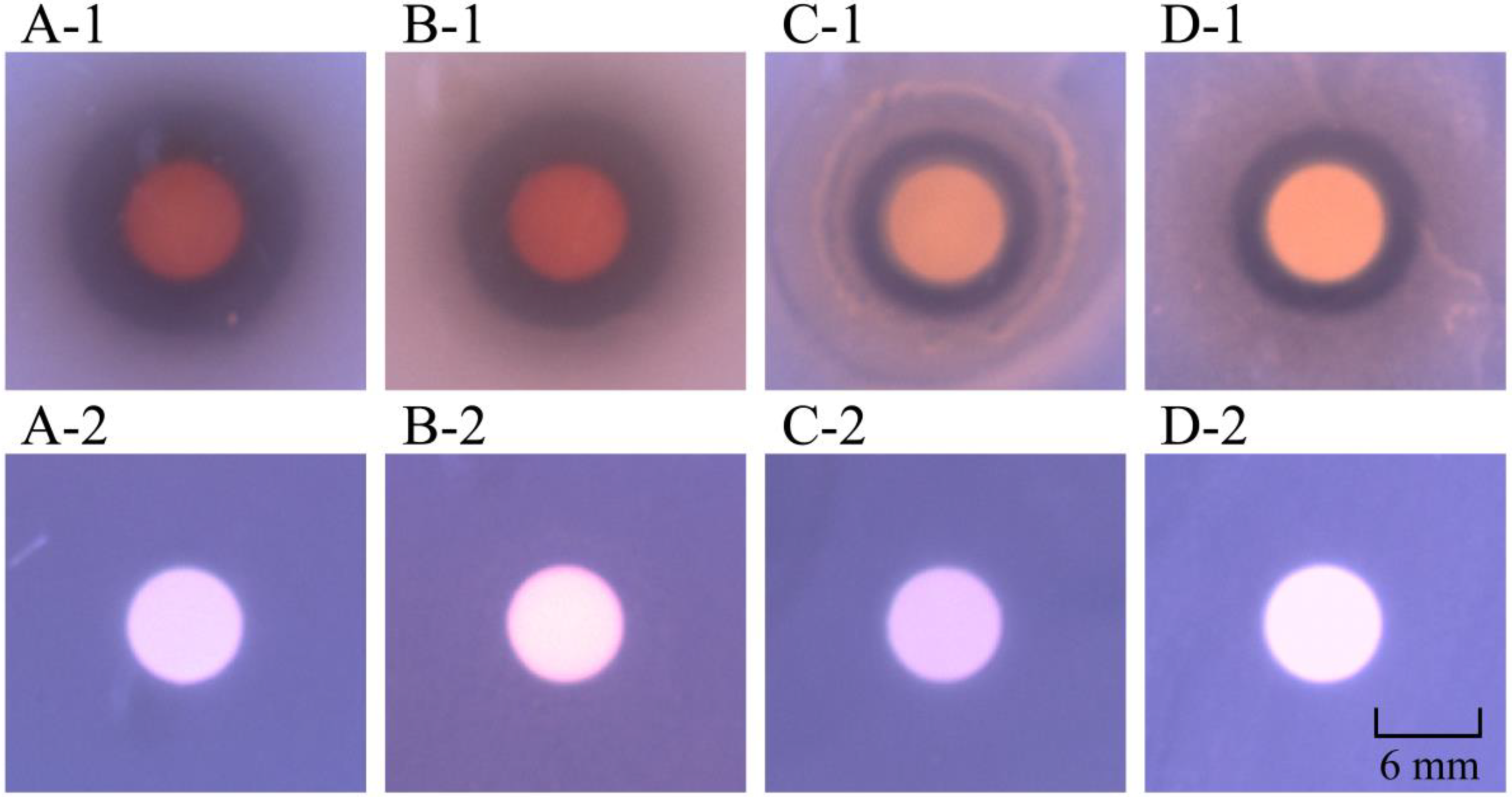
2.1. Antibacterial Activity of Crude Extracts from P. kleiniana Wight et Arn
2.2. Purification of the Methanol-Phase Crude Extract from P. kleiniana Wight et Arn
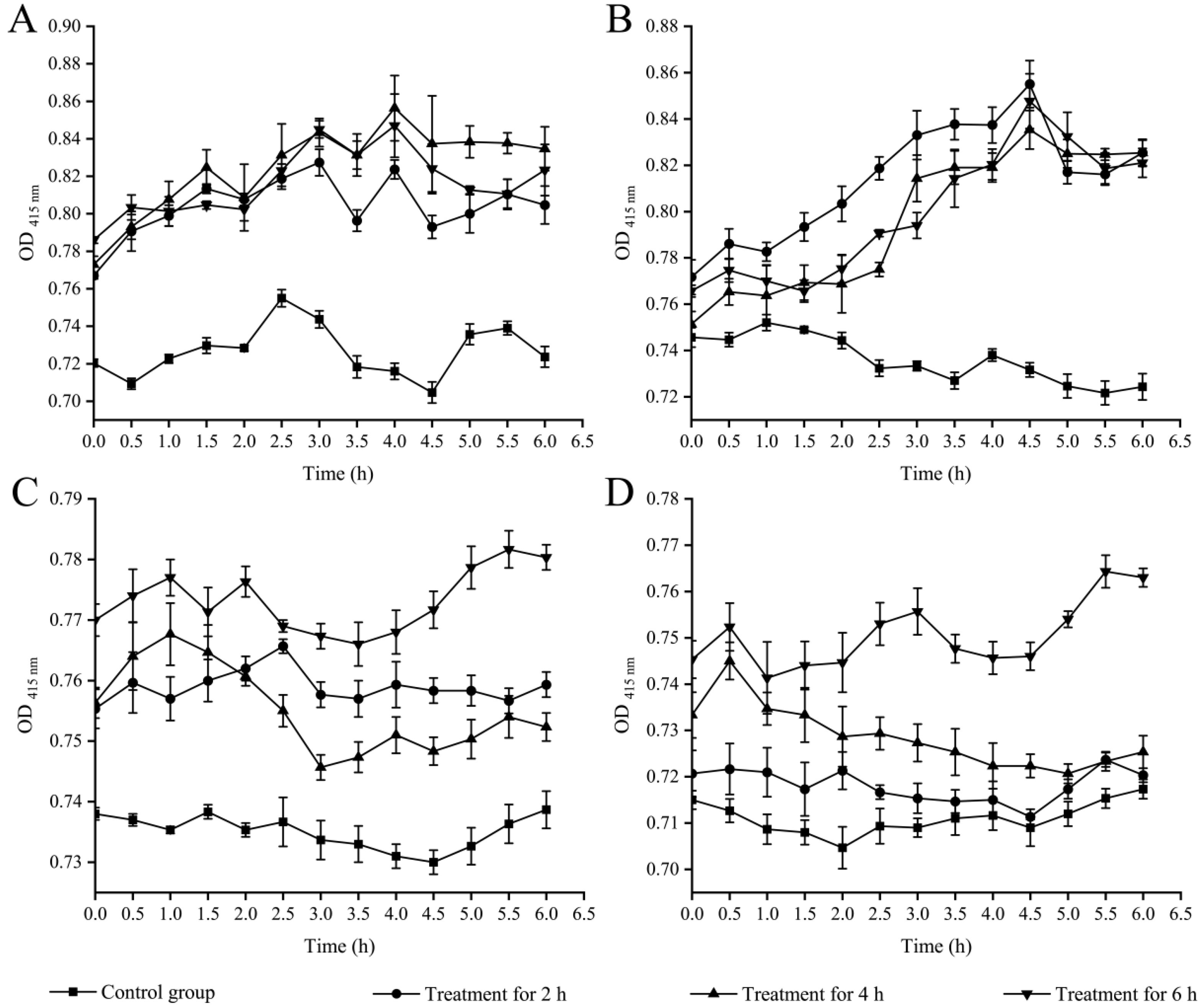
2.3. Bacterial Cell Surface Hydrophobicity, Membrane Fluidity and Permeability Changes Triggered by Fragment 1 from P. kleiniana Wight et Arn
2.3.1. Cell Surface Hydrophobicity
2.3.2. Cell Membrane Fluidity
2.3.3. Cell Membrane Permeability
2.4. Bacterial Cell Surface Structure Changes Triggered by Fragment 1 from P. kleiniana Wight et Arn
2.5. Identification of Potential Antibacterial Compounds in Fragment 1 from P. kleiniana Wight et Arn
2.6. Differential Transcriptomes Triggered by Fragment 1 from P. kleiniana Wight et Arn
2.6.1. The Major Changed Metabolic Pathways in V. parahaemolyticus ATCC17802
2.6.2. The Major Changed Metabolic Pathways in S. aureus ATCC25923
3. Materials and Methods
3.1. Bacterial Strains and Culture Conditions
3.2. Extraction of Bioactive Substances from P. kleiniana Wight et Arn
3.3. Antimicrobial Susceptibility Assay
3.4. Prep-HPLC Analysis
3.5. UHPLC–MS Analysis
3.6. Bacterial Cell Surface Hydrophobicity and Membrane Fluidity Assays
3.7. Cell Membrane Permeability Analysis
3.8. Scanning Electron Microscope (SEM) Assay
3.9. Illumina RNA Sequencing
3.10. RT-qPCR Assay
3.11. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bueno, E.; Pinedo, V.; Cava, F. Adaptation of Vibrio cholerae to hypoxic environments. Front. Microbiol. 2020, 11, 739. [Google Scholar] [CrossRef]
- Stocco, G.; Lucafò, M.; Decorti, G. Pharmacogenomics of antibiotics. Int. J. Mol. Sci. 2020, 21, 5975. [Google Scholar] [CrossRef]
- Bombaywala, S.; Mandpe, A.; Paliya, S.; Kumar, S. Antibiotic resistance in the environment: A critical insight on its occurrence, fate, and eco-toxicity. Environ. Sci. Pollut. Res. Int. 2021, 28, 24889–24916. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial antibiotic resistance: The most critical pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.; Rodrigues, M.E.; Martins, N.; Ferreira, I.C.F.R.; Henriques, M. Phenolic plant extracts versus penicillin G: In vitro susceptibility of Staphylococcus aureus isolated from bovine mastitis. Pharmaceuticals 2019, 12, 128. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef]
- Chandra, G.; Mukherjee, D.; Ray, A.S.; Chatterjee, S.; Bhattacharjee, I. Phytoextracts as antibacterials: A review. Curr. Drug Discov. Technol. 2020, 17, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Volis, S. Securing a future for China’s plant biodiversity through an integrated conservation approach. Plant Divers. 2018, 40, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Yan, S.; Zhou, C.; Liu, Q.; Zhu, H.; Wen, Z. Total flavonoids from Potentilla kleiniana Wight et Arn inhibits biofilm formation and virulence factors production in methicillin-resistant Staphylococcus aureus (MRSA). J. Ethnopharmacol. 2021, 279, 114383. [Google Scholar] [CrossRef]
- Zhou, Y.Q.; Li, S.M.; Wei, X.; Yang, X.; Xiao, J.W.; Pan, B.W.; Xie, S.X.; Zhou, Y.; Yang, J.; Wei, Y. Identification and quantitative analysis of bioactive components from Potentilla kleiniana Wight et Arn with anti HIV-1 proteases activity. Nat. Prod. Res. 2022, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huang, X.; Liu, Q.; Li, X.; Chen, M.; Zhu, Y.; Chen, X. Separation of α-glucosidase inhibitors from Potentilla kleiniana Wight et Arn using solvent and flow-rate gradient high-speed counter-current chromatography target-guided by ultrafiltration HPLC-MS screening. Phytochem. Anal. 2019, 30, 661–668. [Google Scholar] [CrossRef]
- Li, S.; Tan, J.; Zeng, J.; Wu, X.W.X.; Zhang, J. Antihyperglycemic and antioxidant effect of the total flavones of Potentilla kleiniana Wight et Arn. in streptozotocin induced diabetic rats. Pak. J. Pharm. Sci. 2017, 30, 171–178. [Google Scholar]
- Tao, J.; Yan, S.; Wang, H.; Zhao, L.; Zhu, H.; Wen, Z. Antimicrobial and antibiofilm effects of total flavonoids from Potentilla kleiniana Wight et Arn on Pseudomonas aeruginosa and its potential application to stainless steel surfaces. LWT-Food Sci. Technol. 2022, 154, 112631. [Google Scholar] [CrossRef]
- Xuan, S.H.; Hong, I.K.; Lee, Y.J.; Kim, J.W.; Park, S.N. Biological activities and chemical components of Potentilla kleiniana Wight & Arn. Nat. Prod. Res. 2020, 34, 3262–3266. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, L.; Liu, P.; Jin, Y.; Qin, S.; Chen, L. Identification of antibacterial components in the methanol-phase extract from edible herbaceous plant Rumex madaio Makino and their antibacterial action modes. Molecules 2022, 27, 660. [Google Scholar] [CrossRef]
- Fu, J.; Wang, Y.; Sun, M.; Xu, Y.; Chen, L. Antibacterial activity and components of the methanol-phase extract from rhizomes of pharmacophagous plant Alpinia officinarum Hance. Molecules 2022, 27, 4308. [Google Scholar] [CrossRef]
- Wang, J.; Ding, Q.; Yang, Q.; Fan, H.; Yu, G.; Liu, F.; Bello, B.K.; Zhang, X.; Zhang, T.; Dong, J.; et al. Vibrio alginolyticus triggers inflammatory response in mouse peritoneal macrophages via activation of NLRP3 inflammasome. Front. Cell. Infect. Microbiol. 2021, 11, 769777. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Robles, M.F.; Natividad-Bonifacio, I.; Álvarez-Contreras, A.K.; Tercero-Alburo, J.J.; Quiñones-Ramírez, E.I.; Vázquez-Salinas, C. Characterization of potential virulence factors of Vibrio mimicus isolated from fishery products and water. Int. J. Microbiol. 2021, 2021, 8397930. [Google Scholar] [CrossRef]
- Dey, R.; Rieger, A.M.; Stephens, C.; Ashbolt, N.J. Interactions of Pseudomonas aeruginosa with Acanthamoeba polyphaga observed by imaging flow cytometry. Cytom. Part A 2019, 95, 555–564. [Google Scholar] [CrossRef]
- Liu, J.; Qin, K.; Wu, C.; Fu, K.; Yu, X.; Zhou, L. De Novo sequencing provides insights into the pathogenicity of foodborne Vibrio parahaemolyticus. Front. Cell. Infect. Microbiol. 2021, 11, 652957. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Flint, S.H.; Palmer, J.S. Bacillus cereus spores and toxins—The potential role of biofilms. Food Microbiol. 2020, 90, 103493. [Google Scholar] [CrossRef]
- Ojha, R.; Dittmar, A.A.; Severin, G.B.; Koestler, B.J. Shigella flexneri diguanylate cyclases regulate virulence. J. Bacteriol. 2021, 203, e0024221. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Tang, T.; Stegger, M.; Dalsgaard, A.; Liu, T.; Leisner, J.J. Characterization of antimicrobial-resistant Staphylococcus aureus from retail foods in Beijing, China. Food Microbiol. 2021, 93, 103603. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Song, G.; Sun, M.; Wang, J.; Wang, Y. Prevalence and therapies of antibiotic-resistance in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2020, 10, 107. [Google Scholar] [CrossRef]
- Danchik, C.; Casadevall, A. Role of cell surface hydrophobicity in the pathogenesis of medically-significant fungi. Front. Cell. Infect. Microbiol. 2020, 10, 594973. [Google Scholar] [CrossRef] [PubMed]
- Soundharrajan, I.; Kim, D.; Kuppusamy, P.; Muthusamy, K.; Lee, H.J.; Choi, K.C. Probiotic and triticale silage fermentation potential of Pediococcus pentosaceus and Lactobacillus brevis and their impacts on pathogenic bacteria. Microorganisms 2019, 7, 318. [Google Scholar] [CrossRef]
- Yu, X.; Sha, L.; Liu, Q.; Zhao, Y.; Fang, H.; Cao, Y.; Zhao, J. Recent advances in cell membrane camouflage-based biosensing application. Biosens. Bioelectron. 2021, 194, 113623. [Google Scholar] [CrossRef]
- Zhang, M.; Yu, Y.; Lian, L.; Li, W.; Ren, J.; Liang, Y.; Xue, F.; Tang, F.; Zhu, X.; Ling, J.; et al. Functional mechanism of antimicrobial peptide bomidin and its safety for Macrobrachium rosenbergii. Probiotics Antimicrob. Proteins 2022, 14, 169–179. [Google Scholar] [CrossRef]
- Bojkovic, J.; Richie, D.L.; Six, D.A.; Rath, C.M.; Sawyer, W.S.; Hu, Q.; Dean, C.R. Characterization of an acinetobacter baumannii lptD deletion strain: Permeability defects and response to inhibition of lipopolysaccharide and fatty acid biosynthesis. J. Bacteriol. 2015, 198, 731–741. [Google Scholar] [CrossRef]
- Veiko, A.G.; Olchowik-Grabarek, E.; Sekowski, S.; Roszkowska, A.; Lapshina, E.A.; Dobrzynska, I.; Zamaraeva, M.; Zavodnik, I.B. Antimicrobial activity of quercetin, naringenin and catechin: Flavonoids inhibit Staphylococcus aureus-induced hemolysis and modify membranes of bacteria and erythrocytes. Molecules 2023, 28, 1252. [Google Scholar] [CrossRef]
- Zdybicka-Barabas, A.; Stączek, S.; Pawlikowska-Pawlęga, B.; Mak, P.; Luchowski, R.; Skrzypiec, K.; Mendyk, E.; Wydrych, J.; Gruszecki, W.I.; Cytryńska, M. Studies on the interactions of neutral Galleria mellonella cecropin D with living bacterial cells. Amino Acids 2019, 51, 175–191. [Google Scholar] [CrossRef]
- Mizzi, L.; Maniscalco, D.; Gaspari, S.; Chatzitzika, C.; Gatt, R.; Valdramidis, V.P. Assessing the individual microbial inhibitory capacity of different sugars against pathogens commonly found in food systems. Lett. Appl. Microbiol. 2020, 71, 251–258. [Google Scholar] [CrossRef]
- Fan, X.; Kong, D.; He, S.; Chen, J.; Jiang, Y.; Ma, Z.; Feng, J.; Yan, H. Phenanthrene derivatives from asarum heterotropoides showed excellent antibacterial activity against phytopathogenic bacteria. J. Agric. Food Chem. 2021, 69, 14520–14529. [Google Scholar] [CrossRef]
- Tan, Z.; Deng, J.; Ye, Q.; Zhang, Z. The antibacterial activity of natural-derived flavonoids. Curr. Top. Med. Chem. 2022, 22, 1009–1019. [Google Scholar] [CrossRef]
- Kachur, K.; Suntres, Z. The antibacterial properties of phenolic isomers, carvacrol and thymol. Crit. Rev. Food Sci. Nutr. 2020, 60, 3042–3053. [Google Scholar] [CrossRef]
- Wibowo, J.T.; Ahmadi, P.; Rahmawati, S.I.; Bayu, A.; Putra, M.Y.; Kijjoa, A. marine-derived indole alkaloids and their biological and pharmacological activities. Mar. Drugs 2021, 20, 3. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Wang, Y.; Liu, L.; Dong, H.; Satoh, T. Membrane-active amino acid-coupled polyetheramine derivatives with high selectivity and broad-spectrum antibacterial activity. Acta Biomater. 2022, 142, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Maklashina, E.; Cecchini, G.; Iverson, T.M. The roles of SDHAF2 and dicarboxylate in covalent flavinylation of SDHA, the human complex II flavoprotein. Proc. Natl. Acad. Sci. USA 2020, 117, 23548–23556. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.K.; Wijaya, A.; Kohlstedt, M.; Gläser, L.; Brear, P.; Silva-Rocha, R.; Wittmann, C.; Welch, M. Systems-wide dissection of organic acid assimilation in Pseudomonas aeruginosa Reveals a Novel Path to Underground Metabolism. Mbio 2022, 13, e0254122. [Google Scholar] [CrossRef]
- Park, C.; Shin, B.; Park, W. Alternative fate of glyoxylate during acetate and hexadecane metabolism in Acinetobacter oleivorans DR1. Sci. Rep. 2019, 9, 14402. [Google Scholar] [CrossRef] [PubMed]
- Durall, C.; Kukil, K.; Hawkes, J.A.; Albergati, A.; Lindblad, P.; Lindberg, P. Production of succinate by engineered strains of Synechocystis PCC 6803 overexpressing phosphoenolpyruvate carboxylase and a glyoxylate shunt. Microb. Cell Factories 2021, 20, 39. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, Z.; Zhao, M.; Ma, Y.; Luo, Z.; Li, S.; Xu, H. Metabolic engineering of Escherichia coli for ectoine production with a fermentation strategy of supplementing the amino donor. Front. Bioeng. Biotechnol. 2022, 10, 824859. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.K.; Nourse, A.; White, S.W.; Rock, C.O.; Heath, R.J. Crystal structure and allosteric regulation of the cytoplasmic Escherichia coli L-asparaginase I. J. Mol. Biol. 2007, 369, 794–811. [Google Scholar] [CrossRef]
- Wang, Z.; Bie, P.; Cheng, J.; Lu, L.; Cui, B.; Wu, Q. The ABC transporter YejABEF is required for resistance to antimicrobial peptides and the virulence of Brucella melitensis. Sci. Rep. 2016, 6, 31876. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Huang, L.; Zhao, L.; Qin, Y.; Su, Y.; Yan, Q. The regulation of oxidative phosphorylation pathway on Vibrio alginolyticus adhesion under adversities. Microbiologyopen 2019, 8, e00805. [Google Scholar] [CrossRef]
- Santucci, R.; Sinibaldi, F.; Cozza, P.; Polticelli, F.; Fiorucci, L. Cytochrome c: An extreme multifunctional protein with a key role in cell fate. Int. J. Biol. Macromol. 2019, 136, 1237–1246. [Google Scholar] [CrossRef]
- Maza-Márquez, P.; Lee, M.D.; Detweiler, A.M.; Bebout, B.M. Millimeter-scale vertical partitioning of nitrogen cycling in hypersaline mats reveals prominence of genes encoding multi-heme and prismane proteins. ISME J. 2022, 16, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef]
- Liu, F.; Yao, Q.; Huang, J.; Wan, J.; Xie, T.; Gao, X.; Sun, D.; Zhang, F.; Bei, W.; Lei, L. The two-component system CpxA/CpxR is critical for full virulence in Actinobacillus pleuropneumoniae. Front. Microbiol. 2022, 13, 1029426. [Google Scholar] [CrossRef]
- Hu, M.; Huang, X.; Xu, X.; Zhang, Z.; He, S.; Zhu, J.; Liu, H.; Shi, X. Characterization of the role of two-component systems in antibiotic resistance formation in Salmonella enterica Serovar Enteritidis. mSphere 2022, 7, e0038322. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Siasat, P.; McNeil, H.E.; Colclough, A.; Ricci, V.; Lawler, A.J.; Abdalaal, H.; Buckner, M.M.C.; Baylay, A.; Busby, S.J.; et al. EnvR is a potent repressor of acrAB transcription in Salmonella. J. Antimicrob. Chemother. 2022, 78, 133–140. [Google Scholar] [CrossRef]
- Oberlies, J.; Watzl, C.; Giese, T.; Luckner, C.; Kropf, P.; Müller, I. Regulation of NK cell function by human granulocyte arginase. J. Immunol. 2009, 182, 5259–5267. [Google Scholar] [CrossRef]
- Kim, S.H.; Sierra, R.A.; McGee, D.J.; Zabaleta, J. Transcriptional profiling of gastric epithelial cells infected with wild type or arginase-deficient Helicobacter pylori. BMC Microbiol. 2012, 12, 175. [Google Scholar] [CrossRef]
- Yurgel, S.N.; Johnson, S.A.; Rice, J.; Sa, N.; Bailes, C.; Baumgartner, J.; Pitzer, J.E.; Roop, R.M., II; Roje, S. A novel formamidase is required for riboflavin biosynthesis in invasive bacteria. J. Biol. Chem. 2022, 298, 102377. [Google Scholar] [CrossRef]
- Lai, R.Z.; Parkinson, J.S. Monitoring two-component sensor kinases with a chemotaxis signal readout. Methods Mol. Biol. 2018, 1729, 127–135. [Google Scholar] [CrossRef]
- Tan, L.; Huang, Y.; Shang, W.; Yang, Y.; Peng, H.; Hu, Z.; Wang, Y.; Rao, Y.; Hu, Q.; Rao, X.; et al. Accessory gene regulator (agr) allelic variants in cognate Staphylococcus aureus strain display similar phenotypes. Front. Microbiol. 2022, 13, 700894. [Google Scholar] [CrossRef]
- Párraga Solórzano, P.K.; Shupe, A.C.; Kehl-Fie, T.E. The sensor histidine kinase ArlS is necessary for Staphylococcus aureus to activate ArlR in response to nutrient availability. J. Bacteriol. 2021, 203, e0042221. [Google Scholar] [CrossRef]
- Loiseau, L.; Vergnes, A.; Ezraty, B. Methionine oxidation under anaerobic conditions in Escherichia coli. Mol. Microbiol. 2022, 118, 387–402. [Google Scholar] [CrossRef]
- Fang, J.; Yan, L.; Tan, M.; Li, G.; Liang, Y.; Li, K. Nitrogen removal characteristics of a marine denitrifying Pseudomonas stutzeri BBW831 and a simplified strategy for improving the denitrification performance under stressful conditions. Mar. Biotechnol. 2023, 25, 109–122. [Google Scholar] [CrossRef]
- Alvarez, L.; Sanchez-Hevia, D.; Sánchez, M.; Berenguer, J. A new family of nitrate/nitrite transporters involved in denitrification. Int. Microbiol. 2019, 22, 19–28. [Google Scholar] [CrossRef]
- Harborne, N.R.; Griffiths, L.; Busby, S.J.; Cole, J.A. Transcriptional control, translation and function of the products of the five open reading frames of the Escherichia coli nir operon. Mol. Microbiol. 1992, 6, 2805–2813. [Google Scholar] [CrossRef]
- Léger, L.; Byrne, D.; Guiraud, P.; Germain, E.; Maisonneuve, E. NirD curtails the stringent response by inhibiting RelA activity in Escherichia coli. Elife 2021, 10, e64092. [Google Scholar] [CrossRef] [PubMed]
- Kanagarajan, S.; Dhamodharan, P.; Mutharasappan, N.; Choubey, S.K.; Jayaprakash, P.; Biswal, J.; Jeyaraman, J. Structural insights on binding mechanism of CAD complexes (CPSase, ATCase and DHOase). J. Biomol. Struct. Dyn. 2021, 39, 3144–3157. [Google Scholar] [CrossRef] [PubMed]
- Garavito, M.F.; Narváez-Ortiz, H.Y.; Zimmermann, B.H. Pyrimidine metabolism: Dynamic and versatile pathways in pathogens and cellular development. J. Genet. Genom. 2015, 42, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Fu, H.; Chen, D.; Shao, Z.; Zhu, J.; Alali, W.Q.; Chen, L. Simple visualized detection method of virulence-associated genes of Vibrio cholerae by loop-mediated isothermal amplification. Front. Microbiol. 2019, 10, 2899. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, L.; Pandak, W.M.; Heuman, D.; Hylemon, P.B.; Ren, S. High Glucose Induces Lipid Accumulation via 25-Hydroxycholesterol DNA-CpG Methylation. iScience 2020, 23, 101102. [Google Scholar] [CrossRef]
- Shan, X.; Fu, J.; Li, X.; Peng, X.; Chen, L. Comparative proteomics and secretomics revealed virulence, and coresistance-related factors in non O1/O139 Vibrio cholerae recovered from 16 species of consumable aquatic animals. J. Proteom. 2022, 251, 104408. [Google Scholar] [CrossRef]
- Cui, J.; Hölzl, G.; Karmainski, T.; Tiso, T.; Kubicki, S.; Thies, S.; Blank, L.M.; Jaeger, K.E.; Dörmann, P. The glycine-glucolipid of Alcanivorax borkumensis is resident to the bacterial cell wall. Appl. Environ. Microbiol. 2022, 88, e0112622. [Google Scholar] [CrossRef]
- Kuhry, J.G.; Duportail, G.; Bronner, C.; Laustriat, G. Plasma membrane fluidity measurements on whole living cells by fluorescence anisotropy of trimethylammoniumdiphenylhexatriene. Biochim. Biophys. Acta. 1985, 845, 60–67. [Google Scholar] [CrossRef]
- Wang, Z.; Qin, Q.; Zheng, Y.; Li, F.; Zhao, Y.; Chen, G.Q. Engineering the permeability of Halomonas bluephagenesis enhanced its chassis properties. Metab. Eng. 2021, 67, 53–66. [Google Scholar] [CrossRef]
- Huang, B.; Liu, X.; Li, Z.; Zheng, Y.; Wai Kwok Yeung, K.; Cui, Z.; Liang, Y.; Zhu, S.; Wu, S. Rapid bacteria capturing and killing by AgNPs/N-CD@ZnO hybrids strengthened photo-responsive xerogel for rapid healing of bacteria-infected wounds. Chem. Eng. J. 2021, 414, 128805. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Yu, P.; Ren, S.; Zhu, Z.; Jin, Y.; Yan, J.; Peng, X.; Chen, L. Prophage-related gene VpaChn25_0724 contributes to cell membrane integrity and growth of Vibrio parahaemolyticus CHN25. Front. Cell. Infect. Microbiol. 2020, 10, 595709. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Inhibition Zone (Diameter, mm) | MIC (mg/mL) | ||
|---|---|---|---|---|
| CPE | MPE | CPE | MPE | |
| Aeromonas hydrophila ATCC35654 | - | - | - | - |
| Bacillus cereus A1-1 | 7.03 ± 0.01 | 10.54 ± 0.48 | 50 | 6.25 |
| Bacillus cereus A2-2 | 7.11 ± 0.02 | 10.54 ± 0.75 | 50 | 1.56 |
| Enterobacter cloacae ATCC13047 | 7.00 ± 0.11 | 7.11 ± 0.26 | 50 | 50 |
| Enterobacter cloacae C1-1 | - | - | - | - |
| Escherichia coli ATCC8739 | - | 7.62 ± 0.37 | - | 25 |
| Escherichia coli ATCC25922 | - | - | - | - |
| Escherichia coli K12 | - | 7.51 ± 0.29 | - | 25 |
| Enterobacter sakazakii CMCC45401 | - | - | - | - |
| Klebsiella pneumoniae 8-2-10-8 | - | - | - | - |
| Klebsiella pneumoniae 8-2-1-14 | - | - | - | - |
| Pseudomonas aeruginosa ATCC9027 | - | 10.51 ± 0.41 | - | 6.25 |
| Pseudomonas aeruginosa ATCC27853 | - | 8.14 ± 0.32 | - | 25 |
| Salmonella enterica subsp. enterica (ex Kauffmann and Edwards) Le Minor and Popoff serovar Choleraesuis ATCC13312 | - | - | - | - |
| Salmonella paratyphi-A CMCC50093 | - | - | - | - |
| Salmonellaenterica subsp. enterica (ex Kauffmann and Edwards) Le Minor and Popoff serovar Vellore ATCC15611 | 7.09 ± 0.09 | 10.11 ± 0.61 | 50 | 6.25 |
| Salmonella E1-1 | - | - | - | - |
| Shigella dysenteriae CMCC51252 | 7.02 ± 0.11 | 9.29 ± 0.51 | 50 | 12.5 |
| Shigella flexneri CMCC51572 | 7.82 ± 0.20 | 10.17 ± 0.20 | 25 | 6.25 |
| Shigella flexneri ATCC12022 | - | - | - | - |
| Shigella flexneri CMCC51574 | - | 9.17 ± 0.21 | - | 12.5 |
| Shigella sonnei ATCC25931 | 7.00 ± 0.11 | 8.19 ± 0.51 | 50 | 25 |
| Shigella sonnet CMCC51592 | - | - | - | - |
| Staphylococcus aureus ATCC25923 | 7.03 ± 0.14 | 9.41 ± 0.27 | 50 | 12.5 |
| Staphylococcus aureus ATCC8095 | 7.07 ± 0.15 | 10.15 ± 0.24 | 50 | 6.25 |
| Staphylococcus aureus ATCC29213 | 7.78 ± 0.10 | 9.21 ± 0.01 | 25 | 12.5 |
| Staphylococcus aureus ATCC6538 | 7.62 ± 0.61 | 9.55 ± 0.37 | 25 | 12.5 |
| Staphylococcus aureus D1-1 | 7.11 ± 0.25 | 7.00 ± 0.51 | 50 | 50 |
| Vibrio alginolyticus ATCC17749 | - | 10.11 ± 0.24 | - | 3.13 |
| Vibrio alginolyticus ATCC33787 | - | - | - | - |
| Vibrio cholerae GIM1.449 | - | 7.00 ± 0.14 | - | 50 |
| Vibrio cholerae Q10-54 | 7.22 ± 0.10 | 7.02 ± 0.21 | 50 | 50 |
| Vibrio fluvialis ATCC33809 | - | 7.12 ± 0.03 | - | 50 |
| Vibrio harvey ATCC BAA-1117 | - | - | - | - |
| Vibrio harveyi ATCC33842 | - | - | - | - |
| Vibrio mimicus bio-56759 | 7.21 ± 0.41 | 11.00 ± 0.32 | 25 | 3.13 |
| Vibrio parahemolyticus ATCC17802 | - | 10.67 ± 1.21 | - | 1.56 |
| Vibrio parahemolyticus ATCC33847 | 8.63 ± 0.24 | 7.14 ± 0.12 | 12.5 | 50 |
| Vibrio parahemolyticus B3-13 | 7.17 ± 0.29 | 12.33 ± 0.65 | 50 | 3.13 |
| Vibrio parahemolyticus B4-10 | - | 11.26 ± 0.34 | - | 6.25 |
| Vibrio parahemolyticus B5-29 | 7.17 ± 0.04 | 13.77 ± 0.85 | 50 | 3.13 |
| Vibrio parahemolyticus B9-35 | 7.20 ± 0.09 | 13.15 ± 0.44 | 25 | 3.13 |
| Vibrio parahemolyticus A1-1 | 7.13 ± 0.15 | 10.35 ± 0.58 | 50 | 3.13 |
| Vibrio vulnificus ATCC27562 | 7.65 ± 0.44 | 7.01 ± 0.23 | 25 | 50 |
| Strain | Inhibition Zone (Diameter, mm) | MIC (mg/mL) |
|---|---|---|
| B. cereus A2-2 | 8.03 ± 0.45 | 6.25 |
| S. flexneri CMCC51572 | 7.50 ± 0.50 | 6.25 |
| S. aureus ATCC25923 | 8.03 ± 0.40 | 12.5 |
| S. aureus ATCC8095 | 9.53 ± 0.35 | 6.25 |
| S. aureus ATCC6538 | 7.10 ± 0.36 | 50.0 |
| V. parahemolyticus ATCC17802 | 10.31 ± 0.62 | 6.25 |
| V. parahemolyticus A1-1 | 8.57 ± 0.60 | 25.0 |
| V. parahemolyticus B3-13 | 10.37 ± 0.32 | 6.25 |
| V. parahemolyticus B4-10 | 10.30 ± 0.50 | 12.5 |
| V. parahemolyticus B5-29 | 11.30 ± 0.26 | 6.25 |
| V. parahemolyticus B9-35 | 11.27 ± 0.40 | 12.5 |
| Peak No. | Identified Compound | Compound Nature | Rt (min) | Formula | Exact Mass | Peak Area (%) |
|---|---|---|---|---|---|---|
| 1 | D-Maltose | Carbohydrates | 0.76 | C12H22O11 | 342.1162 | 6.77% |
| 2 | Oxymorphone | Phenanthrenes and derivatives | 11.18 | C17H19NO4 | 301.1314 | 6.29% |
| 3 | Rutin | Flavonoids | 12.99 | C27H30O16 | 281.0899 | 6.29% |
| 4 | D-Proline | Amino acid and derivatives | 0.76 | C5H9NO2 | 115.0633 | 5.41% |
| 5 | L-Proline | Amino acid and derivatives | 0.73 | C5H9NO2 | 115.0633 | 5.41% |
| 6 | L-Glutamic acid | Amino acid and derivatives | 0.66 | C5H9NO4 | 147.0532 | 5.20% |
| 7 | Sucrose | Carbohydrates | 0.89 | C12H22O11 | 342.1162 | 3.62% |
| 8 | Cynaroside | Flavonoids | 12.98 | C21H20O11 | 282.162 | 3.37% |
| 9 | Piperlonguminine | Alkaloids | 10.57 | C16H19NO3 | 273.1365 | 3.21% |
| 10 | 5-Aminovaleric acid | Amino acid and derivatives | 1.11 | C5H11NO2 | 117.079 | 3.12% |
| 11 | D-Glutamine | Carboxylic acids and derivatives | 0.66 | C5H10N2O3 | 146.0691 | 2.99% |
| 12 | L-Lysine | Amino acid and derivatives | 0.64 | C6H14N2O2 | 146.1055 | 2.99% |
| 13 | p-Octopamine | Phenols | 3.84 | C8H11NO2 | 153.079 | 2.96% |
| 14 | Oleic acid | Fatty acyls | 13.03 | C18H34O2 | 282.2559 | 2.91% |
| 15 | Isoquercitrin | Flavonoids | 10.58 | C21H20O12 | 274.1933 | 2.44% |
| 16 | L-Pipecolic acid | Amino acid and derivatives | 0.69 | C6H11NO2 | 129.079 | 2.31% |
| 17 | Moracin C | Phenols | 0.67 | C19H18O4 | 129.0426 | 2.31% |
| 18 | Kojibiose | Fatty acyls | 0.72 | C12H22O11 | 342.1162 | 2.22% |
| 19 | Gluconic acid | Carbohydrates | 0.69 | C6H12O7 | 196.0583 | 1.97% |
| 20 | Betaine | Alkaloids | 1.06 | C5H11NO2 | 117.079 | 1.51% |
| 21 | L-Valine | Amino acid and derivatives | 0.93 | C5H11NO2 | 117.079 | 1.49% |
| 22 | D-alpha-Aminobutyric acid | Carboxylic acids and derivatives | 0.65 | C4H9NO2 | 103.0633 | 1.46% |
| 23 | cis-Aconitic acid | Organic acids and derivatives | 1.46 | C6H6O6 | 174.0164 | 1.34% |
| 24 | Lactulose | Organooxygen compounds | 0.77 | C12H22O11 | 342.1162 | 1.33% |
| 25 | Turanose | Fatty acyls | 0.79 | C12H22O11 | 342.1162 | 1.33% |
| 26 | L-Pipecolic acid | Amino acid and derivatives | 1.47 | C6H11NO2 | 129.079 | 1.15% |
| 27 | DL-Norvaline | Amino acid and derivatives | 1.05 | C5H11NO2 | 117.079 | 1.11% |
| 28 | L-Asparagine | Amino acid and derivatives | 0.64 | C4H8N2O3 | 132.0535 | 1.11% |
| 29 | Malic acid | Hydroxy acids and derivatives | 0.8 | C4H6O5 | 134.0215 | 0.90% |
| 30 | Trigonelline | Alkaloids | 0.82 | C7H7NO2 | 137.0477 | 0.90% |
| 31 | Acetamide | Alkaloids | 13.95 | C2H5NO | 59.03711 | 0.88% |
| 32 | Beta-D-fructose 2-phosphate | Organooxygen compounds | 0.75 | C6H13O9P | 260.0297 | 0.77% |
| 33 | 22-Dehydroclerosterol | Steroids | 12.59 | C29H46O | 410.3549 | 0.76% |
| 34 | Artemisinin | Sesquiterpenoids | 13.02 | C15H22O5 | 282.1467 | 0.72% |
| 35 | Kaempferol-3-O-rutinoside | flavonoids | 6.29 | C27H30O15 | 594.1585 | 0.54% |
| 36 | L-Homoserine | Amino acid and derivatives | 0.67 | C4H9NO3 | 119.0582 | 0.52% |
| 37 | L-Threonine | Amino acid and derivatives | 0.64 | C4H9NO3 | 119.0582 | 0.50% |
| 38 | Palmitic acid | Lipids | 12.92 | C16H32O2 | 256.2402 | 0.49% |
| 39 | O-Acetylethanolamine | Alkaloids | 0.67 | C4H9NO2 | 103.0633 | 0.46% |
| 40 | Galactose 1-phosphate | Organooxygen compounds | 0.65 | C6H13O9P | 260.0297 | 0.46% |
| 41 | Glucose 1-phosphate | Organooxygen compounds | 13 | C6H13O9P | 260.0297 | 0.45% |
| 42 | Adenosine 5′-monophosphate | Nucleotide and its derivates | 1.38 | C10H14N5O7P | 347.0631 | 0.43% |
| 43 | L-Arginine | Amino acid and derivatives | 0.6 | C6H14N4O2 | 174.1117 | 0.43% |
| 44 | Maltotriose | Organooxygen compounds | 1.23 | C18H32O16 | 504.169 | 0.40% |
| 45 | Indole | Alkaloids | 3.82 | C8H7N | 117.0578 | 0.38% |
| 46 | D-Glucose 6-phosphate | Carbohydrates | 0.65 | C6H13O9P | 260.0297 | 0.37% |
| 47 | D-Aspartic acid | Alkaloids | 0.76 | C4H7NO4 | 133.0375 | 0.36% |
| 48 | Vitexin rhamnoside | Flavonoids | 6.78 | C27H30O14 | 578.1636 | 0.35% |
| 49 | L-Aspartic acid | Amino acid and derivatives | 0.63 | C4H7NO4 | 133.0375 | 0.33% |
| 50 | Maltol | Flavonoids | 0.9 | C6H6O3 | 126.0317 | 0.33% |
| 51 | Astragalin | Flavonoids | 6.52 | C21H20O11 | 448.1006 | 0.32% |
| 52 | 3-Hydroxy-3-methylpentane-1,5-dioic acid | Amino acid and derivatives | 2.32 | C6H10O5 | 162.0528 | 0.31% |
| 53 | Campesterol | Steroids and steroid derivatives | 12.18 | C28H48O | 400.3705 | 0.30% |
| 54 | L-Ornithine | Amino acid and derivatives | 0.55 | C5H12N2O2 | 132.0899 | 0.30% |
| 55 | Adenosine | Nucleotide and its derivates | 2.58 | C10H13N5O4 | 267.0968 | 0.29% |
| 56 | Vidarabine | Purine nucleosides | 2.28 | C10H13N5O4 | 267.0968 | 0.27% |
| 57 | Nicotinic acid | Nicotinic acid derivatives | 0.73 | C6H5NO2 | 123.032 | 0.27% |
| 58 | Pelargonidin-3-O-glucoside | Flavonoids | 1.11 | C21H20O10 | 100.0524 | 0.26% |
| 59 | L-Citruline | Amino acid and derivatives | 0.66 | C6H13N3O3 | 175.0957 | 0.26% |
| 60 | Diallyl disulfide | Miscellaneous | 0.68 | C6H10S2 | 146.0224 | 0.26% |
| 61 | Sarracine | Alkaloids | 13.14 | C18H27NO5 | 337.1889 | 0.22% |
| 62 | N-Acetylputrescine | Phenolamides | 1.79 | C6H14N2O | 130.1106 | 0.22% |
| 63 | Salicylic acid | Organic acid | 7.06 | C7H6O3 | 138.0317 | 0.22% |
| 64 | 5-Methylcytosine | Nucleotide and its derivates | 2.26 | C5H7N3O | 125.0589 | 0.21% |
| 65 | Ellagic acid | Phenols | 6.12 | C14H6O8 | 302.0063 | 0.21% |
| 66 | Isodiospyrin | Quinones | 11.28 | C22H14O6 | 374.079 | 0.21% |
| Metabolic Pathway | Gene ID | Gene Name | Fold Change | Gene Description |
|---|---|---|---|---|
| Citrate cycle | WU75_19785 | sucA | 0.146 | 2-oxoglutarate dehydrogenase |
| WU75_07425 | pckA | 0.465 | Phosphoenolpyruvate carboxykinase | |
| WU75_19790 | sucB | 0.133 | Dihydrolipoamide succinyltransferase | |
| WU75_11550 | acnB | 0.143 | Bifunctional aconitate hydratase 2/2-methylisocitrate dehydratase | |
| WU75_19795 | sucC | 0.134 | Succinyl-CoA synthetase subunit beta | |
| WU75_19800 | sucD | 0.16 | Succinyl-CoA synthetase subunit alpha | |
| WU75_19770 | sdhD | 0.199 | Succinate dehydrogenase | |
| WU75_19780 | sdhB | 0.157 | Succinate dehydrogenase | |
| WU75_19765 | sdhC | 0.182 | Succinate dehydrogenase | |
| WU75_13785 | fumA | 0.497 | Fumarate hydratase | |
| WU75_09605 | icd | 0.179 | Isocitrate dehydrogenase | |
| WU75_19775 | sdhA | 0.144 | Succinate dehydrogenase | |
| WU75_06430 | mdh | 0.177 | Malate dehydrogenase | |
| WU75_16530 | lpd | 0.35 | Dihydrolipoamide dehydrogenase | |
| Glyoxylate and dicarboxylate metabolism | WU75_19760 | gltA | 0.129 | Type II citrate synthase |
| WU75_19150 | aceA | 0.37 | Isocitrate lyase | |
| WU75_19145 | aceB | 0.352 | Malate synthase | |
| WU75_00290 | aceB | 0.315 | Malate synthase | |
| WU75_10840 | phbB | 0.277 | 3-ketoacyl-ACP reductase | |
| WU75_03265 | katE | 2.389 | Catalase | |
| Fatty acid degradation | WU75_22235 | fadB | 0.151 | Multifunctional fatty acid oxidation complex subunit alpha |
| WU75_08655 | fadE | 0.184 | Acyl-CoA dehydrogenase | |
| WU75_20175 | fadJ | 0.204 | Multifunctional fatty acid oxidation complex subunit alpha | |
| WU75_22230 | fadA | 0.208 | 3-ketoacyl-CoA thiolase | |
| WU75_20180 | fadA | 0.305 | 3-ketoacyl-CoA thiolase | |
| WU75_10835 | atoB | 0.433 | Acetyl-CoA acetyltransferase | |
| WU75_10445 | atoB | 0.445 | Acetyl-CoA acetyltransferase | |
| WU75_12560 | fadE | 0.452 | Acyl-CoA dehydrogenase | |
| WU75_19885 | fadD | 0.493 | Long-chain fatty acid—CoA ligase | |
| Glycine, serine and threonine metabolism | WU75_14910 | gcvP | 0.113 | Glycine dehydrogenase |
| WU75_14915 | gcvH | 0.127 | Glycine cleavage system protein H | |
| WU75_10395 | betA | 0.162 | Choline dehydrogenase | |
| WU75_14930 | gcvT | 0.184 | Glycine cleavage system protein T | |
| WU75_16130 | lysC | 0.187 | Aspartate kinase | |
| WU75_14920 | glyA | 0.203 | Serine hydroxymethyltransferase | |
| WU75_16140 | ectB | 0.222 | Diaminobutyrate-2-oxoglutarate aminotransferase | |
| WU75_16145 | ectA | 0.246 | L-2,4-diaminobutyric acid acetyltransferase | |
| WU75_10400 | betB | 0.259 | Betaine-aldehyde dehydrogenase | |
| WU75_00565 | sdaA | 0.264 | Serine dehydratase | |
| WU75_16135 | ectC | 0.27 | Ectoine synthase | |
| WU75_02030 | trpB | 0.397 | Tryptophan synthase subunit beta | |
| WU75_05755 | thrC | 0.429 | Threonine synthase | |
| WU75_05760 | thrB | 0.47 | Serine kinase | |
| WU75_05330 | glxK | 0.495 | Glycerate kinase | |
| Oxidative phosphorylation | WU75_06010 | petC | 0.195 | Cytochrome C |
| WU75_06015 | petB | 0.209 | Cytochrome B | |
| WU75_14570 | ccoO | 0.228 | Peptidase S41 | |
| WU75_14575 | ccoN | 0.272 | Cbb3-type cytochrome c oxidase subunit I | |
| WU75_14560 | ccoP | 0.301 | Cytochrome Cbb3 | |
| WU75_06485 | ppa | 0.339 | Inorganic pyrophosphatase | |
| WU75_06020 | petA | 0.442 | Ubiquinol-cytochrome C reductase | |
| WU75_14565 | ccoQ | 0.475 | Cytochrome C oxidase | |
| WU75_02240 | cyoC | 0.478 | Cytochrome o ubiquinol oxidase subunit III | |
| WU75_19125 | ppk2 | 2.159 | Polyphosphate kinase | |
| WU75_09420 | cydA | 3.637 | Cytochrome d terminal oxidase subunit 1 | |
| WU75_09415 | cydB | 4.11 | Cytochrome d ubiquinol oxidase subunit 2 | |
| WU75_09410 | cydX | 5.362 | Membrane protein | |
| Pyruvate metabolism | WU75_01940 | yiaY | 0.171 | Alcohol dehydrogenase |
| WU75_03655 | lldD | 0.276 | Lactate dehydrogenase | |
| WU75_22155 | dld | 0.322 | Lactate dehydrogenase | |
| WU75_16665 | oadA | 0.324 | Oxaloacetate decarboxylase | |
| WU75_16060 | aldB | 0.397 | Aldehyde dehydrogenase | |
| WU75_20855 | gloA | 2.451 | Lactoylglutathione lyase | |
| WU75_12805 | pta | 8.464 | Phosphate acetyltransferase | |
| WU75_02150 | ackA | 8.851 | Acetate kinase | |
| WU75_12810 | ackA | 10.365 | Acetate kinase | |
| WU75_09685 | pflD | 12.853 | Pyruvate formate-lyase | |
| WU75_00810 | gloA | 13.536 | Glyoxalase | |
| Propanoate metabolism | WU75_15760 | prpF | 0.402 | 3-methylitaconate isomerase |
| WU75_15770 | prpC | 0.435 | Methylcitrate synthase | |
| beta-Lactam resistance | WU75_09315 | acrA | 6.699 | Hemolysin D |
| WU75_09310 | acrB | 8.911 | Multidrug transporter | |
| WU75_09925 | acrA | 40.366 | Hemolysin D | |
| ABC transporters | WU75_10385 | proW | 0.106 | ABC transporter permease |
| WU75_16175 | proX | 0.116 | Glycine/betaine ABC transporter substrate-binding protein | |
| WU75_10390 | proX | 0.122 | Glycine/betaine ABC transporter substrate-binding protein | |
| WU75_12775 | oppC | 0.133 | Peptide ABC transporter permease | |
| WU75_10380 | proV | 0.138 | ABC transporter ATP-binding protein | |
| WU75_09655 | aotM | 0.143 | Amino acid ABC transporter permease | |
| WU75_09665 | aotJ | 0.144 | Nickel transporter | |
| WU75_13090 | yejA | 0.151 | Diguanylate cyclase | |
| WU75_12770 | oppB | 0.164 | Oligopeptide transporter permease | |
| WU75_12780 | oppD | 0.172 | Oligopeptide transporter ATP-binding component | |
| WU75_09660 | aotQ | 0.176 | ABC transporter | |
| WU75_16170 | proW | 0.199 | Glycine/betaine ABC transporter permease | |
| WU75_08085 | oppA | 0.201 | Peptide ABC transporter substrate-binding protein | |
| WU75_07210 | yejA | 0.204 | Diguanylate cyclase | |
| WU75_12765 | oppA | 0.214 | Peptide ABC transporter substrate-binding protein | |
| WU75_07220 | yejB | 0.22 | Hypothetical protein | |
| WU75_07215 | yejE | 0.221 | Peptide ABC transporter permease | |
| WU75_09670 | aotP | 0.228 | Amino acid transporter | |
| WU75_12785 | oppF | 0.228 | Peptide ABC transporter ATP-binding protein | |
| WU75_04720 | oppA | 0.341 | Peptide ABC transporter substrate-binding protein | |
| WU75_16165 | proV | 0.343 | Glycine/betaine ABC transporter ATP-binding protein | |
| WU75_14765 | aapQ | 0.377 | Amino acid ABC transporter permease | |
| WU75_03180 | malE | 0.4 | Sugar ABC transporter substrate-binding protein | |
| WU75_14775 | aapP | 0.405 | ABC transporter ATP-binding protein | |
| WU75_04605 | vcaM | 0.406 | Multidrug ABC transporter ATP-binding protein | |
| WU75_14055 | mdlB | 0.411 | Multidrug ABC transporter ATP-binding protein | |
| WU75_10275 | rbsD | 0.438 | D-ribose pyranase | |
| WU75_05845 | btuF | 0.487 | Vitamin B12-binding protein | |
| WU75_14760 | aapJ | 0.491 | Amino acid ABC transporter substrate-binding protein | |
| WU75_03185 | malK | 2.175 | Maltose/maltodextrin transporter ATP-binding protein | |
| WU75_19815 | znuA | 2.204 | Zinc ABC transporter substrate-binding protein | |
| WU75_19810 | znuC | 2.491 | Zinc ABC transporter ATPase | |
| WU75_02265 | artP | 2.617 | Arginine ABC transporter ATP-binding protein | |
| WU75_19805 | znuB | 2.666 | Membrane protein | |
| WU75_00425 | macB | 14.353 | Macrolide transporter | |
| Two-component system | WU75_07480 | glnG | 0.186 | Nitrogen regulation protein NR(I) |
| WU75_13735 | mcp | 0.218 | Chemotaxis protein | |
| WU75_15795 | tctB | 0.237 | TctB | |
| WU75_21750 | dctD | 0.288 | C4-dicarboxylate ABC transporter | |
| WU75_13155 | ttrB | 0.31 | 4Fe-4S ferredoxin | |
| WU75_21770 | dctP | 0.31 | C4-dicarboxylate ABC transporter | |
| WU75_01920 | mcp | 0.32 | Chemotaxis protein | |
| WU75_21745 | dctB | 0.352 | ATPase | |
| WU75_10200 | phoA | 0.353 | Alkaline phosphatase | |
| WU75_21765 | dctQ | 0.368 | C4-dicarboxylate ABC transporter permease | |
| WU75_00210 | dctD | 0.406 | C4-dicarboxylate ABC transporter | |
| WU75_16210 | qseC | 0.423 | Histidine kinase | |
| WU75_23015 | fliC | 0.435 | Flagellin | |
| WU75_07100 | mcp | 0.453 | Chemotaxis protein | |
| WU75_13380 | crp | 0.457 | Transcriptional regulator | |
| WU75_09825 | mcp | 0.471 | Chemotaxis protein | |
| WU75_16525 | hapR | 0.477 | LuxR family transcriptional regulator | |
| WU75_15800 | tctA | 0.485 | Tripartite tricarboxylate transporter TctA | |
| WU75_14800 | mcp | 0.491 | Chemotaxis protein | |
| WU75_06085 | tolC | 2.068 | Outer membrane channel protein | |
| WU75_15630 | dcuB | 2.125 | C4-dicarboxylate transporter | |
| WU75_06045 | degP | 2.148 | Serine endoprotease DegQ | |
| WU75_04355 | mcp | 2.163 | Chemotaxis protein | |
| WU75_10915 | luxQ | 3.377 | ATPase | |
| WU75_22175 | mcp | 4.001 | Chemotaxis protein | |
| WU75_02450 | pfeR | 4.828 | Transcriptional regulator | |
| WU75_18570 | cpxA | 10.981 | Two-component sensor protein | |
| WU75_18575 | cpxR | 26.5 | Transcriptional regulator | |
| Alanine, aspartate and glutamate metabolism | WU75_06265 | glmS | 0.037 | Glucosamine-fructose-6-phosphate Aminotransferase |
| WU75_07465 | glnA | 0.123 | Glutamine synthetase | |
| WU75_04655 | putA | 0.145 | Pyrroline-5-carboxylate dehydrogenase | |
| WU75_14680 | - | 0.286 | NAD-glutamate dehydrogenase | |
| WU75_05875 | carB | 0.343 | Carbamoyl phosphate synthase large subunit | |
| WU75_05820 | gltB | 0.414 | Glutamate synthase | |
| WU75_05825 | gltD | 0.44 | Glutamate synthase | |
| WU75_05880 | carA | 0.46 | Carbamoyl phosphate synthase small subunit | |
| WU75_18095 | pyrI | 0.462 | Aspartate carbamoyltransferase regulatory subunit | |
| WU75_18090 | pyrB | 0.466 | Aspartate carbamoyltransferase catalytic subunit | |
| WU75_20915 | ansA | 2.141 | Cytoplasmic asparaginase I | |
| WU75_01110 | ansB | 2.718 | L-asparaginase II | |
| WU75_18550 | aspA | 7.015 | Aspartate ammonia-lyase | |
| PTS | WU75_03285 | ptsN | 0.462 | PTS fructose transporter subunit IIA |
| WU75_12990 | ptsG | 0.5 | PTS glucose transporter subunit IIBC | |
| WU75_17910 | celC | 2.36 | Molecular chaperone TorD | |
| WU75_14970 | fruB | 2.451 | Bifunctional PTS system fructose-Specific transporter subunit IIA/HPr protein | |
| WU75_19555 | ptsH | 3.973 | PTS sugar transporter | |
| WU75_00455 | ulaB | 3.977 | PTS ascorbate transporter subunit IIB | |
| WU75_19550 | ptsI | 4.075 | Phosphoenolpyruvate-protein Phosphotransferase | |
| WU75_00460 | cmtB | 4.118 | PTS system mannitol-specific Transporter subunit IIA | |
| WU75_01640 | cmtB | 4.539 | PTS mannitol transporter subunit IIA | |
| WU75_14960 | fruA | 5.096 | PTS fructose transporter subunit IIBC | |
| WU75_00450 | ulaA | 6.946 | PTS beta-glucoside transporter subunit IIBC | |
| Butanoate metabolism | WU75_01985 | acsA | 0.334 | Acetoacetyl-CoA synthetase |
| WU75_10825 | phaC | 0.336 | Poly(3-hydroxyalkanoate) synthetase | |
| Lysine degradation | WU75_21960 | ldcC | 7.207 | Lysine decarboxylase LdcC |
| QS | WU75_07805 | - | 0.109 | Cytochrome C |
| WU75_07800 | - | 0.181 | ABC transporter permease | |
| WU75_07795 | - | 0.202 | ABC transporter permease | |
| WU75_07810 | ddpD | 0.216 | ABC transporter ATP-binding protein | |
| WU75_11620 | - | 0.218 | Peptide ABC transporter permease | |
| WU75_11630 | - | 0.233 | Peptide ABC transporter substrate-binding protein | |
| WU75_11625 | - | 0.261 | Peptide ABC transporter permease | |
| WU75_11610 | ddpF | 0.358 | Chemotaxis protein | |
| WU75_11615 | ddpD | 0.484 | Sugar ABC transporter ATP-binding protein | |
| WU75_21410 | aphA | 2.288 | Transcriptional regulator | |
| Nitrogen metabolism | WU75_00760 | ncd2 | 0.276 | 2-nitropropane dioxygenase |
| WU75_10810 | napA | 2.286 | Nitrate reductase | |
| WU75_15655 | nirD | 3.934 | Nitrite reductase | |
| WU75_10815 | napB | 6.27 | Nitrate reductase | |
| WU75_08850 | hcp | 63.107 | Hydroxylamine reductase |
| Metabolic Pathway | Gene ID | Gene Name | Fold Change | Gene Description |
|---|---|---|---|---|
| Two-component system | KQ76_00500 | - | 0.373 | Capsular biosynthesis protein |
| KQ76_00560 | wecC | 0.490 | UDP-N-acetyl-D-mannosamine dehydrogenase | |
| KQ76_12475 | nreC | 2.117 | Nitrate respiration regulation response regulator NreC | |
| KQ76_12480 | nreB | 2.276 | Nitrate respiration regulation sensor histidine kinase NreB | |
| KQ76_12485 | nreA | 2.433 | Nitrate respiration regulation accessory nitrate sensor NreA | |
| KQ76_10520 | agrB | 2.565 | Histidine kinase | |
| KQ76_03245 | graS | 2.989 | Histidine kinase | |
| KQ76_10785 | kdpF | 5.371 | ATPase | |
| KQ76_04230 | dltC | 28.924 | Alanine-phosphoribitol ligase | |
| Nitrogen metabolism | KQ76_12490 | narI | 3.529 | Nitrate reductase |
| KQ76_12515 | nirD | 4.199 | Nitrite reductase | |
| KQ76_12520 | nirB | 5.060 | Nitrite reductase | |
| KQ76_12460 | narT | 6.376 | Nitrate transporter NarT | |
| KQ76_12500 | narH | 5.799 | Nitrate reductase | |
| KQ76_12505 | narZ | 8.442 | Nitrate reductase | |
| KQ76_12495 | narJ | 10.404 | Nitrate reductase | |
| Riboflavin metabolism | KQ76_09200 | ribE | 0.373 | Riboflavin synthase subunit alpha |
| KQ76_09195 | ribBA | 0.413 | GTP cyclohydrolase | |
| KQ76_09205 | ribD | 0.430 | Diaminohydroxyphosphoribosylaminopyrimidine deaminase | |
| KQ76_09190 | ribH | 0.480 | 6,7-dimethyl-8-ribityllumazine synthase | |
| Arginine and proline metabolism | KQ76_09185 | fadM | 0.109 | Proline dehydrogenase |
| KQ76_00580 | - | 0.218 | Aldehyde dehydrogenase | |
| KQ76_13360 | - | 0.303 | 1-pyrroline-5-carboxylate dehydrogenase | |
| KQ76_11235 | rocF | 0.461 | Arginase | |
| Atrazine degradation | KQ76_11915 | ureC | 0.406 | Urease subunit alpha |
| KQ76_11910 | ureB | 0.412 | Urease subunit beta | |
| Alanine, aspartate and glutamate metabolism | KQ76_13360 | - | 0.303 | 1-pyrroline-5-carboxylate dehydrogenase |
| KQ76_05770 | carB | 2.158 | Carbamoyl phosphate synthase large subunit | |
| KQ76_05765 | carA | 3.084 | Carbamoyl phosphate synthase small subunit | |
| Pyrimidine metabolism | KQ76_05745 | pyrR | 2.968 | Phosphoribosyl transferase |
| KQ76_05760 | pyrC | 3.115 | Dihydroorotase | |
| KQ76_05755 | pyrB | 3.213 | Aspartate carbamoyltransferase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Yu, P.; Chen, L. Identification of Antibacterial Components and Modes in the Methanol-Phase Extract from a Herbal Plant Potentilla kleiniana Wight et Arn. Foods 2023, 12, 1640. https://doi.org/10.3390/foods12081640
Tang Y, Yu P, Chen L. Identification of Antibacterial Components and Modes in the Methanol-Phase Extract from a Herbal Plant Potentilla kleiniana Wight et Arn. Foods. 2023; 12(8):1640. https://doi.org/10.3390/foods12081640
Chicago/Turabian StyleTang, Yingping, Pan Yu, and Lanming Chen. 2023. "Identification of Antibacterial Components and Modes in the Methanol-Phase Extract from a Herbal Plant Potentilla kleiniana Wight et Arn" Foods 12, no. 8: 1640. https://doi.org/10.3390/foods12081640
APA StyleTang, Y., Yu, P., & Chen, L. (2023). Identification of Antibacterial Components and Modes in the Methanol-Phase Extract from a Herbal Plant Potentilla kleiniana Wight et Arn. Foods, 12(8), 1640. https://doi.org/10.3390/foods12081640