
Lactobacillus gasseri LG-G12 Restores Gut Microbiota and Intestinal Health in Obesity Mice on Ceftriaxone Therapy

 ,
,  , ,
, ,  ,
,
Abstract
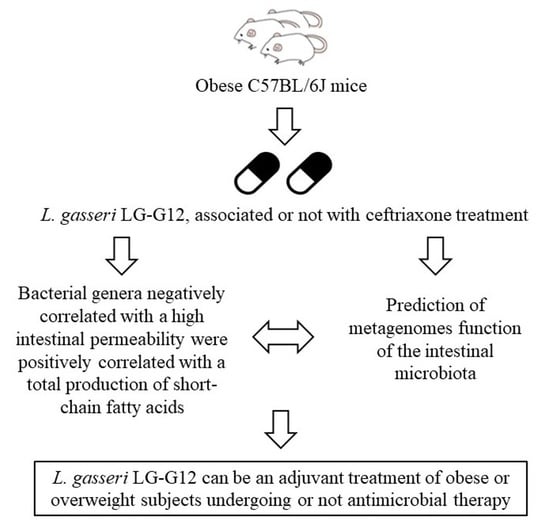
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design and Diet
2.3. Intestinal Permeability
2.4. Quantification of Short-Chain Fatty Acids
2.5. Composition and Functional Prediction of the Intestinal Microbiota
2.6. Statistical Analysis
3. Results and Discussion
3.1. Multidimensional Scaling Analysis (MDS)
3.2. Alpha and Beta Diversity
3.3. Linear Discriminant Analysis Effect Size (LEfSe)
3.3.1. Phylum Level
3.3.2. Genus Level
3.4. Correlation Analyses
3.4.1. Groups Treated with LG-G12
3.4.2. Groups Treated with Ceftriaxone
3.4.3. Groups Treated with Ceftriaxone Followed by LG-G12
3.5. Functional Predictions of the Gut Microbiota
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbial ecology linked to obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H. Definition and roles of the gut microbiota. Rev. Prat. 2019, 69, 776–782. [Google Scholar] [PubMed]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [PubMed]
- Maruvada, P.; Leone, V.; Kaplan, L.M.; Chang, E.B. The Human Microbiome and Obesity: Moving beyond Associations. Cell Host Microbe 2017, 22, 589–599. [Google Scholar] [CrossRef]
- Hunt, R.; Armstrong, D.; Katelaris, P.; Afihene, M.; Bane, A.; Bhatia, S.; Chen, M.H.; Choi, M.G.; Melo, A.C.; Fock, K.M.; et al. World Gastroenterology Organisation Global Guidelines. J. Clin. Gastroenterol. 2017, 51, 467–478. [Google Scholar] [CrossRef]
- Crovesy, L.; Ostrowski, M.; Ferreira, D.M.T.P.; Rosado, E.L.; Soares-Mota, M. Effect of Lactobacillus on body weight and body fat in overweight subjects: A systematic review of randomized controlled clinical trials. Int. J. Obes. 2017, 41, 1607–1614. [Google Scholar] [CrossRef]
- Dias, M.M.; Louzano, S.A.R.; Conceição, L.L.; Conceição, R.F.; Mendes, T.A.O.; Pereira, S.S.; Oliveira, L.L.; Peluzio, M.C.G. Antibiotic Followed by a Potential Probiotic Increases Brown Adipose Tissue, Reduces Biometric Measurements, and Changes Intestinal Microbiota Phyla in Obesity. Probiotics Antimicrob. Proteins 2021, 13, 1621–1631. [Google Scholar] [CrossRef]
- Louzano, S.A.R.; Dias, M.M.; Conceição, L.L.; Mendes, T.A.O.; Peluzio, M.C.G. Ceftriaxone causes dysbiosis and changes intestinal structure in adjuvant obesity treatment. Pharmacol. Rep. 2022, 74, 111–123. [Google Scholar] [CrossRef]
- Lange, K.; Buerger, M.; Stallmach, A.; Bruns, T. Effects of antibiotics on gut microbiota. Dig. Dis. 2016, 34, 260–268. [Google Scholar] [CrossRef]
- Kim, S.; Covington, A.; Pamer, E.G. The intestinal microbiota: Antibiotics, colonization resistance, and enteric pathogens. Immunol. Rev. 2017, 279, 90–105. [Google Scholar] [CrossRef]
- Richards, D.M.; Heel, R.C.; Brogden, R.N.; Speight, T.M.; Avery, G.S. Ceftriaxone. Drugs 1984, 27, 469–527. [Google Scholar] [CrossRef]
- Dabke, K.; Hendrick, G.; Devkota, S. The gut microbiome and metabolic syndrome. J. Clin. Investig. 2019, 129, 4050–4057. [Google Scholar] [CrossRef]
- Khalili, H.; Chan, S.S.M.; Lochhead, P.; Ananthakrishnan, A.N.; Hart, A.R.; Chan, A.T. The role of diet in the aetiopathogenesis of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 525–535. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de Los Reyes-Gavilán, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Khoshbin, K.; Camilleri, M. Effects of dietary components on intestinal permeability in health and disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G589–G608. [Google Scholar] [CrossRef]
- Alpino, G.C.Á.; Pereira-Sol, G.A.; Dias, M.M.E.; Aguiar, A.S.; Peluzio, M.D.C.G. Beneficial effects of butyrate on brain functions: A view of epigenetic. Crit. Rev. Food Sci. Nutr. 2022, 1–10. [Google Scholar] [CrossRef]
- Brasil. Conselho Nacional de Controle de Experimentação Animal (CONSEA) (2008) Lei no 11794, de 8 de outubro de 2008. Available online: http://www.planalto.gov.br/ccivil_03/_ato2007-2010/2008/lei/l11794.htm (accessed on 3 November 2022).
- Membrez, M.; Blancher, F.; Jaquet, M.; Bibiloni, R.; Cani, P.D.; Burcelin, R.G.; Corthesy, I.; Macé, K.; Chou, C.J. Gut microbiota modulation with norfloxacin and ampicillin enhances glucose tolerance in mice. FASEB J. 2008, 22, 2416–2426. [Google Scholar] [CrossRef]
- Della Vedova, M.C.; Muñoz, M.D.; Santillan, L.D.; Plateo-Pignatari, M.G.; Germanó, M.J.; Rinaldi Tosi, M.E.; Garcia, S.; Gomez, N.N.; Fornes, M.W.; Gomez Mejiba, S.E.; et al. A Mouse Model of Diet-Induced Obesity Resembling Most Features of Human Metabolic Syndrome. Nutr. Metab. Insights 2016, 9, NMI-S32907. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Rajpal, D.K.; Klein, J.L.; Mayhew, D.; Boucheron, J.; Spivak, A.T.; Kumar, V.; Ingraham, K.; Paulik, M.; Chen, L.; Van Horn, S.; et al. Selective Spectrum Antibiotic Modulation of the Gut Microbiome in Obesity and Diabetes Rodent Models. PLoS ONE 2015, 10, e0145499. [Google Scholar] [CrossRef]
- Siegfried, V.R.; Ruckermann, H.; Stumpf, G.; Siegfried, B.D.; Ruckemann, H.; Siegfried, R.; Siegfried, M.R. Method for the determination of organic acids in silage by high performance liquid chromatography. Landwirtsch. Forsch. 1984, 37, 298–304. [Google Scholar]
- Zhang, B.W.; Li, M.; Ma, L.C.; Wei, F.W. A widely applicable protocol for DNA isolation from fecal samples. Biochem. Genet. 2006, 44, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2-approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Barbera, P.; Kozlov, A.M.; Czech, L.; Morel, B.; Darriba, D.; Flouri, T.; Stamatakis, A. EPA-ng: Massively Parallel Evolutionary Placement of Genetic Sequences. Syst. Biol. 2019, 68, 365–369. [Google Scholar] [CrossRef]
- Czech, L.; Stamatakis, A. Scalable methods for analyzing and visualizing phylogenetic placement of metagenomic samples. PLoS ONE 2019, 14, e0217050. [Google Scholar] [CrossRef]
- Louca, S.; Doebeli, M. Efficient comparative phylogenetics on large trees. Bioinformatics 2018, 34, 1053–1055. [Google Scholar] [CrossRef]
- Ye, Y.; Doak, T.G. A parsimony approach to biological pathway reconstruction/inference for genomes and metagenomes. PLoS Comput. Biol. 2009, 5, e1000465. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes—A 2019 update. Nucleic Acids Res. 2020, 48, D445–D453. [Google Scholar] [CrossRef]
- Cheng, R.; Liang, H.; Zhang, Y.; Guo, J.; Miao, Z.; Shen, X.; Chen, G.; Cheng, G.; Li, M.; He, F. Contributions of Lactobacillus plantarum PC170 administration on the recovery of gut microbiota after short-term ceftriaxone exposure in mice. Benef. Microbes 2020, 11, 489–509. [Google Scholar] [CrossRef]
- Ianiro, G.; Tilg, H.; Gasbarrini, A. Antibiotics as deep modulators of gut microbiota: Between good and evil. Gut 2016, 11, 1906–1915. [Google Scholar] [CrossRef]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; Qin, H. Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 2019, 60, 175–184. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, K.S.; Dolan, B.; Eklund, L.; Wising, C.; Ermund, A.; Johansson, Å.; Törnblom, H.; Simren, M.; Hansson, G.C. Association between Brachyspira and irritable bowel syndrome with diarrhoea. Gut 2021, 70, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, B.; Mu, L.; Wang, H.; Luo, J.; Yang, Y.; Yang, H.; Li, M.; Zhou, L.; Tao, C. Long-Term Exposure to Ceftriaxone Sodium Induces Alteration of Gut Microbiota Accompanied by Abnormal Behaviors in Mice. Front. Cell. Infect. Microbiol. 2020, 10, 258. [Google Scholar] [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A central, enigmatic component of the human gut microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef]
- Guo, S.; Zhao, H.; Ma, Z.; Zhang, S.; Li, M.; Zheng, Z.; Ren, X.; Ho, C.T.; Bai, N. Anti-Obesity and Gut Microbiota Modulation Effect of Secoiridoid-Enriched Extract from Fraxinus mandshurica Seeds on High-Fat Diet-Fed Mice. Molecules 2020, 25, 4001. [Google Scholar] [CrossRef]
- Wang, J.; Wang, P.; Li, D.; Hu, X.; Chen, F. Beneficial effects of ginger on prevention of obesity through modulation of gut microbiota in mice. Eur. J. Nutr. 2020, 59, 699–718. [Google Scholar] [CrossRef]
- Zhou, L.; Xiao, X.; Zhang, Q.; Zheng, J.; Li, M.; Wang, X.; Deng, M.; Zhai, X.; Liu, J. Gut microbiota might be a crucial factor in deciphering the metabolic benefits of perinatal genistein consumption in dams and adult female offspring. Food Funct. 2019, 10, 4505–4521. [Google Scholar] [CrossRef]
- Machate, D.J.; Figueiredo, P.S.; Marcelino, G.; Guimarães, R.C.A.; Hiane, P.A.; Bogo, D.; Pinheiro, V.A.Z.; Oliveira, L.C.S.; Pott, A. Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis. Int. J. Mol. Sci. 2020, 21, 4093. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Deng, R.; Zhang, Y.; Zheng, X. Dietary anthocyanin-rich extract of açai protects from diet-induced obesity, liver steatosis, and insulin resistance with modulation of gut microbiota in mice. Nutrition 2021, 86, 111176. [Google Scholar] [CrossRef]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring bacterial community of human gut microbiota reveals an increase in Lactobacillus in obese patients and Methanogens in anorexic patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef]
- Million, M.; Maraninchi, M.; Henry, M.; Armougom, F.; Richet, H.; Carrieri, P.; Valero, R.; Raccah, D.; Vialettes, B.; Raoult, D. Obesity-associated gut microbiota is enriched in Lactobacillus reuteri and depleted in Bifidobacterium animalis and Methanobrevibacter smithii. Int. J. Obes. 2012, 36, 817–825. [Google Scholar] [CrossRef]
- Michalovich, D.; Rodriguez-Perez, N.; Smolinska, S.; Pirozynski, M.; Mayhew, D.; Uddin, S.; Van Horn, S.; Sokolowska, M.; Altunbulakli, C.; Eljaszewicz, A.; et al. Obesity and disease severity magnify disturbed microbiome-immune interactions in asthma patients. Nat. Commun. 2019, 10, 5711. [Google Scholar] [CrossRef]
- Langsetmo, L.; Johnson, A.; Demmer, R.T.; Fino, N.; Orwoll, E.S.; Ensrud, K.E.; Hoffman, A.R.; Cauley, J.A.; Shmagel, A.; Meyer, K.; et al. The Association between Objectively Measured Physical Activity and the Gut Microbiome among Older Community Dwelling Men. J. Nutr. Health Aging 2019, 23, 538–546. [Google Scholar] [CrossRef]
- Bailén, M.; Bressa, C.; Martínez-López, S.; González-Soltero, R.; Montalvo Lominchar, M.G.; San Juan, C.; Larrosa, M. Microbiota Features Associated with a High-Fat/Low-Fiber Diet in Healthy Adults. Front. Nutr. 2020, 7, 583608. [Google Scholar] [CrossRef]
- Castaner, O.; Goday, A.; Park, Y.M.; Lee, S.H.; Magkos, F.; Shiow, S.T.E.; Schröder, H. The Gut Microbiome Profile in Obesity: A Systematic Review. Int. J. Endocrinol. 2018, 2018, 4095789. [Google Scholar] [CrossRef]
- Palmas, V.; Pisanu, S.; Madau, V.; Casula, E.; Deledda, A.; Cusano, R.; Uva, P.; Vascellari, S.; Loviselli, A.; Manzin, A.; et al. Gut microbiota markers associated with obesity and overweight in Italian adults. Sci. Rep. 2021, 11, 5532. [Google Scholar] [CrossRef]
- Naito, Y.; Uchiyama, K.; Takagi, T. A next-generation beneficial microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr. 2018, 63, 33–35. [Google Scholar] [CrossRef]
- Vesić, D.; Kristich, C.J. MurAA is required for intrinsic cephalosporin resistance of Enterococcus faecalis. Antimicrob. Agents Chemother. 2012, 56, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Park, D.Y.; Ahn, Y.T.; Park, S.H.; Huh, C.S.; Yoo, S.R.; Yu, R.; Sung, M.K.; McGregor, R.A.; Choi, M.S. Supplementation of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 in diet-induced obese mice is associated with gut microbial changes and reduction in obesity. PLoS ONE 2013, 8, e59470. [Google Scholar] [CrossRef] [PubMed]
- Fine, R.L.; Manfredo Vieira, S.; Gilmore, M.S.; Kriegel, M.A. Mechanisms and consequences of gut commensal translocation in chronic diseases. Gut Microbes 2020, 11, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Effects of dietary Enterococcus faecium NCIMB 11181 supplementation on growth performance and cellular and humoral immune responses in broiler chickens. Poult. Sci. 2019, 98, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Jiang, X.Y.; Zhou, L.S.; Song, J.H.; Zhang, X. Effects of Probiotics on Intestinal Mucosa Barrier in Patients With Colorectal Cancer after Operation: Meta-Analysis of Randomized Controlled Trials. Medicine 2016, 95, e3342. [Google Scholar] [CrossRef]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.R.; Delzenne, N.M. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 2007, 50, 2374–2383. [Google Scholar] [CrossRef]
- De Vuyst, L.; Moens, F.; Selak, M.; Rivière, A.; Leroy, F. Summer Meeting 2013: Growth and physiology of bifidobacteria. J. Appl. Microbiol. 2014, 116, 477–4791. [Google Scholar] [CrossRef]
- Huang, C.C.; Shen, M.H.; Chen, S.K.; Yang, S.H.; Liu, C.Y.; Guo, J.W.; Chang, K.W.; Huang, C.J. Gut butyrate-producing organisms correlate to Placenta Specific 8 protein: Importance to colorectal cancer progression. J. Adv. Res. 2019, 22, 7–20. [Google Scholar] [CrossRef]
- Zeng, J.; Hu, Y.; Jia, T.; Zhang, R.; Su, T.; Sun, J.; Gao, H.; Li, G.; Cao, M.; Song, M. Chemoenzymatic synthesis of sialylated lactuloses and their inhibitory effects on Staphylococcus aureus. PLoS ONE 2018, 13, e0199334. [Google Scholar] [CrossRef]
- Buccigrossi, V.; Lo Vecchio, A.; Marano, A.; Guarino, A. Differential effects of Clostridium difficile toxins on ion secretion and cell integrity in human intestinal cells. Pediatr. Res. 2019, 85, 1048–1054. [Google Scholar] [CrossRef]
- Looney, W.J.; Narita, M.; Mühlemann, K. Stenotrophomonas maltophilia: An emerging opportunist human pathogen. Lancet Infect. Dis. 2009, 9, 312–323. [Google Scholar] [CrossRef]
- Kalidasan, V.; Joseph, N.; Kumar, S.; Awang Hamat, R.; Neela, V.K. Iron and Virulence in Stenotrophomonas Maltophilia: All We Know So Far. Front. Cell. Infect. Microbiol. 2018, 8, 401. [Google Scholar] [CrossRef]
- Chen, M.; Hui, S.; Lang, H.; Zhou, M.; Zhang, Y.; Kang, C.; Zeng, X.; Zhang, Q.; Yi, L.; Mi, M. SIRT3 Deficiency Promotes High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease in Correlation with Impaired Intestinal Permeability through Gut Microbial Dysbiosis. Mol. Nutr. Food Res. 2019, 63, e1800612. [Google Scholar] [CrossRef]
- Manome, A.; Abiko, Y.; Kawashima, J.; Washio, J.; Fukumoto, S.; Takahashi, N. Acidogenic Potential of Oral Bifidobacterium and Its High Fluoride Tolerance. Front. Microbiol. 2019, 10, 1099. [Google Scholar] [CrossRef]
- Aghaie, A.; Lechaplais, C.; Sirven, P.; Tricot, S.; Besnard-Gonnet, M.; Muselet, D.; de Berardinis, V.; Kreimeyer, A.; Gyapay, G.; Salanoubat, M.; et al. New insights into the alternative D-glucarate degradation pathway. J. Biol. Chem. 2008, 283, 15638–15646. [Google Scholar] [CrossRef]
- Verce, M.; De Vuyst, L.; Weckx, S. Comparative genomics of Lactobacillus fermentum suggests a free-living lifestyle of this lactic acid bacterial species. Food Microbiol. 2020, 89, 103448. [Google Scholar] [CrossRef]
- Aussel, L.; Pierrel, F.; Loiseau, L.; Lombard, M.; Fontecave, M.; Barras, F. Biosynthesis and physiology of coenzyme Q in bacteria. Biochim. Biophys. Acta 2014, 1837, 1004–1011. [Google Scholar] [CrossRef]
- Fenn, K.; Strandwitz, P.; Stewart, E.J.; Dimise, E.; Rubin, S.; Gurubacharya, S.; Clardy, J.; Lewis, K. Quinones are growth factors for the human gut microbiota. Microbiome 2017, 5, 161. [Google Scholar] [CrossRef]
- Lee, J.S.; Wang, R.X.; Goldberg, M.S.; Clifford, G.P.; Kao, D.J.; Colgan, S.P. Microbiota-Sourced Purines Support Wound Healing and Mucous Barrier Function. iScience 2020, 23, 101226. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Shannon | Chao1 | Faith_pd | Observed Features |
|---|---|---|---|---|
| G1 | 6.4 ± 0.1 A | 394 ± 7.0 A | 26.4 ± 2.8 | 393.3 ± 6.3 A |
| G2 | 6.1 ± 0.3 A | 363.3 ± 11.2 A | 26.5 ± 1.9 | 363.3 ± 11.2 A |
| G3 | 6.2 ± 0.1 A | 352.7 ± 16.9 A | 24.8 ± 1.7 | 351.0 ± 18.0 A |
| G4 | 2.1 ± 0.8 B | 124.7 ± 27.6 B | 11.7 ± 1.8 | 124.0 ± 27.8 B |
| G5 | 4.7 ± 0.4 AB | 192 ± 50.1 AB | 22.6 ± 3.2 | 228.5 ± 59.5 AB |
| G6 | 3.7 ± 0.5 AB | 217 ± 20.2 AB | 27.6 ± 1.6 | 233.0 ± 19.0 AB |
| G7 | 5.9 ± 0.2 A | 338.3 ± 15.9 A | 25.3 ± 3.7 | 337.6 ± 15.7 AB |
| G8 | 3.5 ± 1.5 AB | 250.5 ± 79.9 AB | 22.3 ± 6.5 | 249.5 ± 138.5 AB |
| G9 | 5.2 ± 0.6 AB | 270 ± 69.64 AB | 24.8 ± 5.7 | 270.0 ± 69.6 AB |
| Group | F/B Ratio | Gram-Negative | Gram-Positive | G+/G− Ratio | Mycoplasma | LAG |
|---|---|---|---|---|---|---|
| G1 | 3.40 B | 62.03% A | 37.80% A | 0.61 | 0.01% | 0.16% A |
| G2 | 1.72 B | 74.51% A | 25.37% A | 0.34 | 0.00% | 0.12% A |
| G3 | 2.41 B | 77.21% A | 22.35% A | 0.29 | 0.00% | 0.44% A |
| G4 | 16.81 A | 23.50% AB | 76.23% AB | 3.24 | 0.07% | 0.21% AB |
| G5 | 0.96 B | 76.56% A | 23.25% A | 0.30 | 0.00% | 0.19% A |
| G6 | 1.46 B | 81.64% AC | 18.29% AC | 0.22 | 0.01% | 0.07% AC |
| G7 | 2.06 B | 49.92% A | 49.96% A | 1.00 | 0.02% | 0.11% A |
| G8 | 1.18 B | 42.74% A | 39.36% A | 0.92 | 14.95% | 2.94% A |
| G9 | 2.07 B | 65.15% A | 34.63% A | 0.53 | 0.06% | 0.16% A |
| Group | Acetate | Propionate | Butyrate | Total SCFA | L/M |
|---|---|---|---|---|---|
| G1 | 571.80 ± 37.40 A | 150.70 ± 17.40 A | 31.35 ± 8.90 A | 753.80 ± 61.00 A | 1.41 ± 0.22 C |
| G2 | 0.19 ± 0.02 D | 0.04 ± 0.01 B | 0.05 ± 0.04 B | 0.28 ± 0.04 C | 55.10 ± 2.25 A |
| G3 | 0.16 ± 0.03 D | 0.04 ± 0.01 B | 0.01 ± 0.01 B | 0.19 ± 0.03 C | 49.30 ± 3.20 A |
| G4 | 0.06 ± 0.02 D | 0.03 ± 0.01 B | - | 0.10 ± 0.03 C | 59.60 ± 3.02 A |
| G5 | 0.20 ± 0.08 D | 0.07 ± 0.03 B | 0.02 ± 0.01 B | 0.27 ± 0.07 C | 27.50 ± 6.70 B |
| G6 | 382.80 ± 44.30 B | 142.20 ± 28.20 A | 7.20 ± 3.60 B | 530.70 ± 68.24 AB | 2.10 ± 0.80 C |
| G7 | 374.70 ± 61.90 B | 140.70 ± 17.70 A | 40.10 ± 4.00 A | 535.40 ± 80.13 AB | 0.98 ± 0.17 C |
| G8 | 165.30 ± 23.40 C | 185.70 ± 17.30 A | - | 351.00 ± 31.00 B | 0.58 ± 0.21 C |
| G9 | 396.00 ± 45.20 B | 211.20 ± 27.90 A | 36.70 ± 6.90 A | 644.00 ± 68.70 A | 2.00 ± 0.23 C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Moura e Dias, M.; da Silva Duarte, V.; Mota, L.F.M.; de Cássia Ávila Alpino, G.; dos Reis Louzano, S.A.; da Conceição, L.L.; Mantovanie, H.C.; Pereira, S.S.; Oliveira, L.L.; de Oliveira Mendes, T.A.; et al. Lactobacillus gasseri LG-G12 Restores Gut Microbiota and Intestinal Health in Obesity Mice on Ceftriaxone Therapy. Foods 2023, 12, 1092. https://doi.org/10.3390/foods12051092
de Moura e Dias M, da Silva Duarte V, Mota LFM, de Cássia Ávila Alpino G, dos Reis Louzano SA, da Conceição LL, Mantovanie HC, Pereira SS, Oliveira LL, de Oliveira Mendes TA, et al. Lactobacillus gasseri LG-G12 Restores Gut Microbiota and Intestinal Health in Obesity Mice on Ceftriaxone Therapy. Foods. 2023; 12(5):1092. https://doi.org/10.3390/foods12051092
Chicago/Turabian Stylede Moura e Dias, Mariana, Vinícius da Silva Duarte, Lúcio Flávio Macedo Mota, Gabriela de Cássia Ávila Alpino, Sandra Aparecida dos Reis Louzano, Lisiane Lopes da Conceição, Hilário Cuquetto Mantovanie, Solange Silveira Pereira, Leandro Licursi Oliveira, Tiago Antônio de Oliveira Mendes, and et al. 2023. "Lactobacillus gasseri LG-G12 Restores Gut Microbiota and Intestinal Health in Obesity Mice on Ceftriaxone Therapy" Foods 12, no. 5: 1092. https://doi.org/10.3390/foods12051092
APA Stylede Moura e Dias, M., da Silva Duarte, V., Mota, L. F. M., de Cássia Ávila Alpino, G., dos Reis Louzano, S. A., da Conceição, L. L., Mantovanie, H. C., Pereira, S. S., Oliveira, L. L., de Oliveira Mendes, T. A., Porcellato, D., & do Carmo Gouveia Peluzio, M. (2023). Lactobacillus gasseri LG-G12 Restores Gut Microbiota and Intestinal Health in Obesity Mice on Ceftriaxone Therapy. Foods, 12(5), 1092. https://doi.org/10.3390/foods12051092