


Biological Control of Aspergillus parasiticus and Aspergillus ochraceus and Reductions in the Amount of Ochratoxin A and Aflatoxins in Bread by Selected Non-Conventional Yeast




Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
Yeast Isolation and Identification
2.2. Confrontation Assay
2.3. Effect on Pathogen Spore Germination In Vitro
2.4. Hydrolytic Enzymes Production
2.4.1. Determination of Enzymes by the Qualitative Method
2.4.2. Determination of Enzymes by the Quantitative Method
2.5. Ability to Survive in Different Environmental Conditions
2.6. Hydrogen Sulfide (H2S) Production
2.7. Biogenic Amines Determination
2.8. Determination of Mycotoxin Content in Model Bread
2.9. Statistical Analysis
3. Results
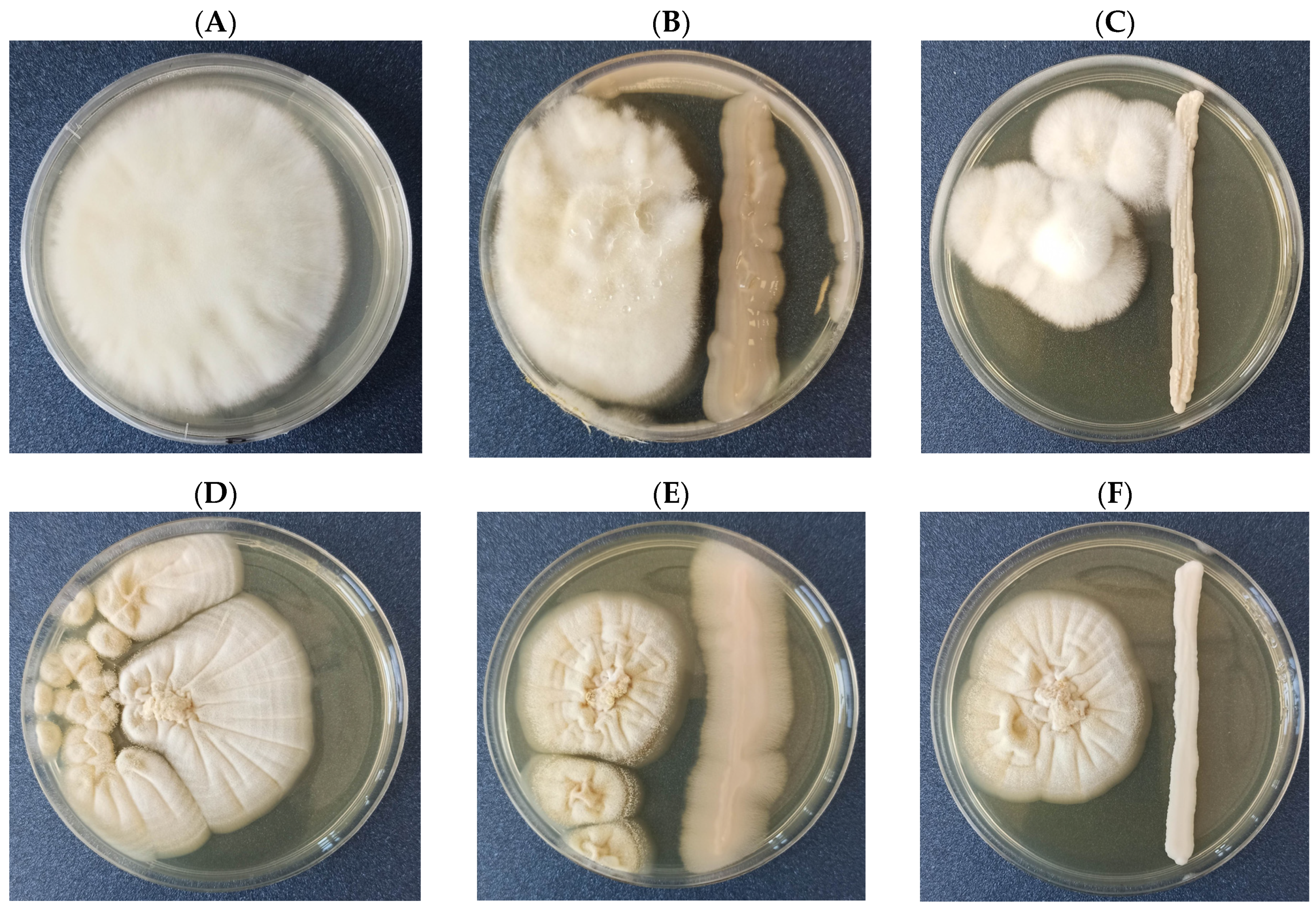
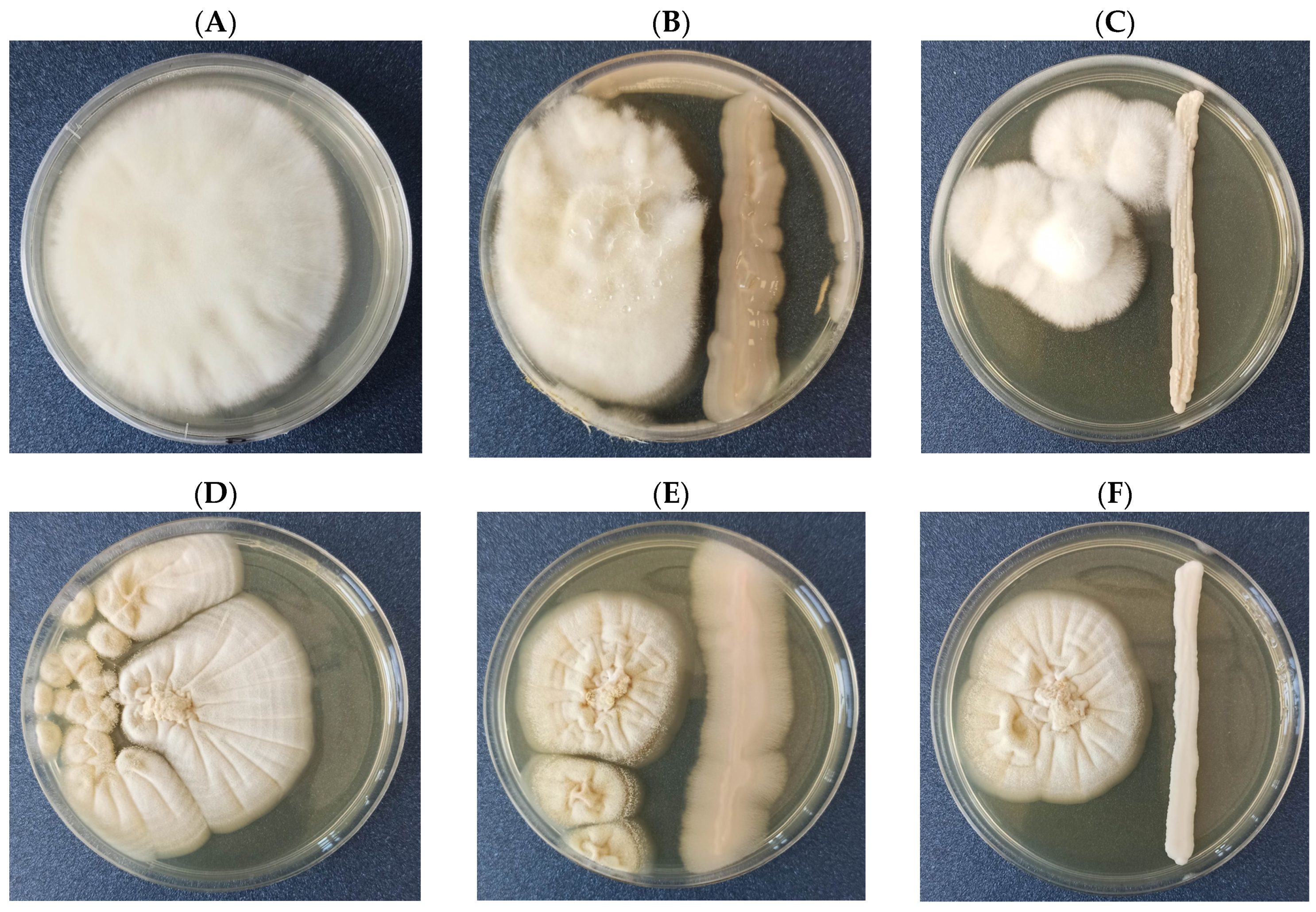
3.1. Confrontation Assay
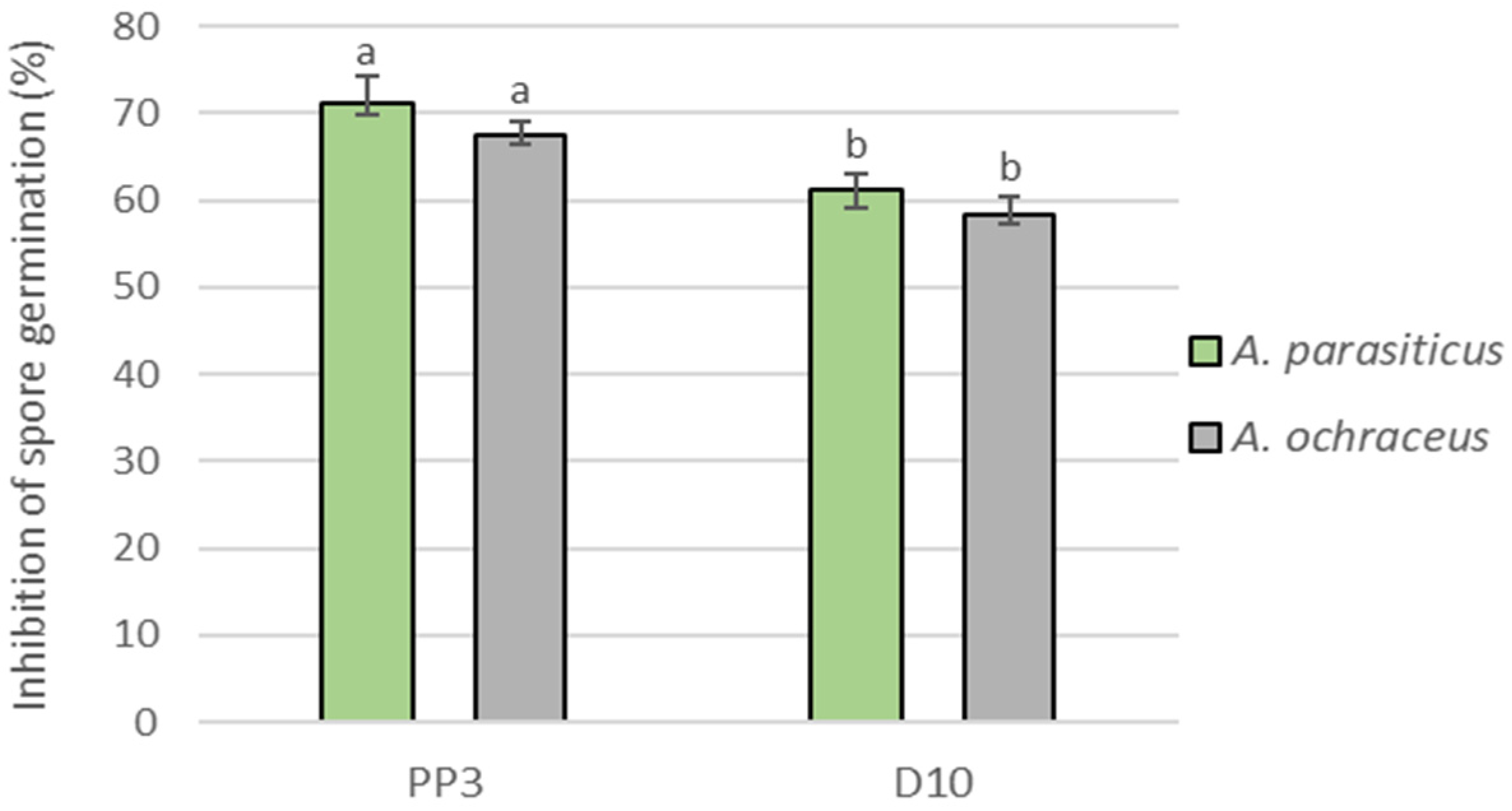
3.2. Effect on Pathogen Spore Germination In Vitro
3.3. Hydrolytic Enzymes Production
3.4. Ability to Survive in Different Environmental Conditions
3.5. Hydrogen Sulfide (H2S) Production
3.6. Biogenic Amines Determination
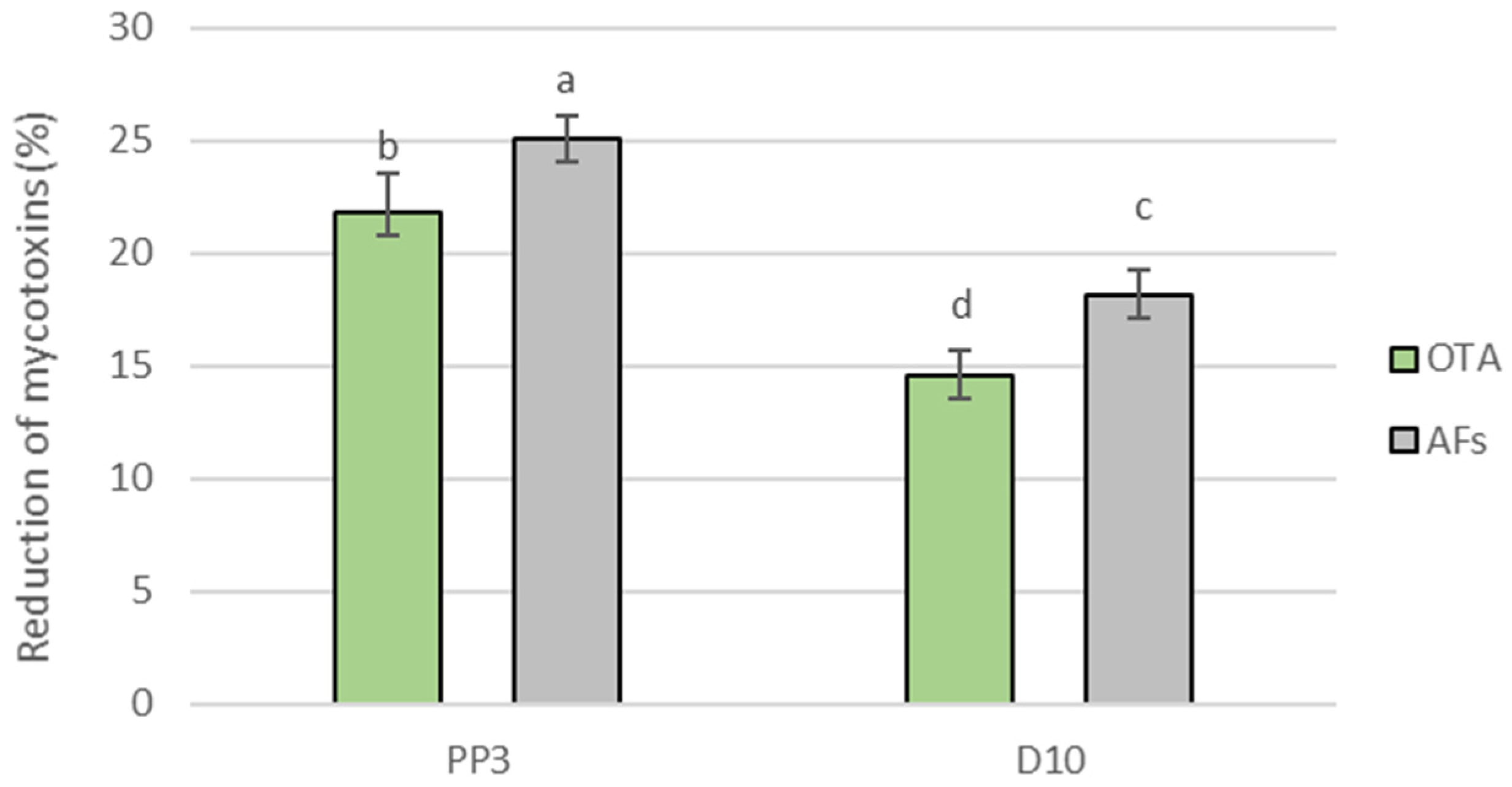
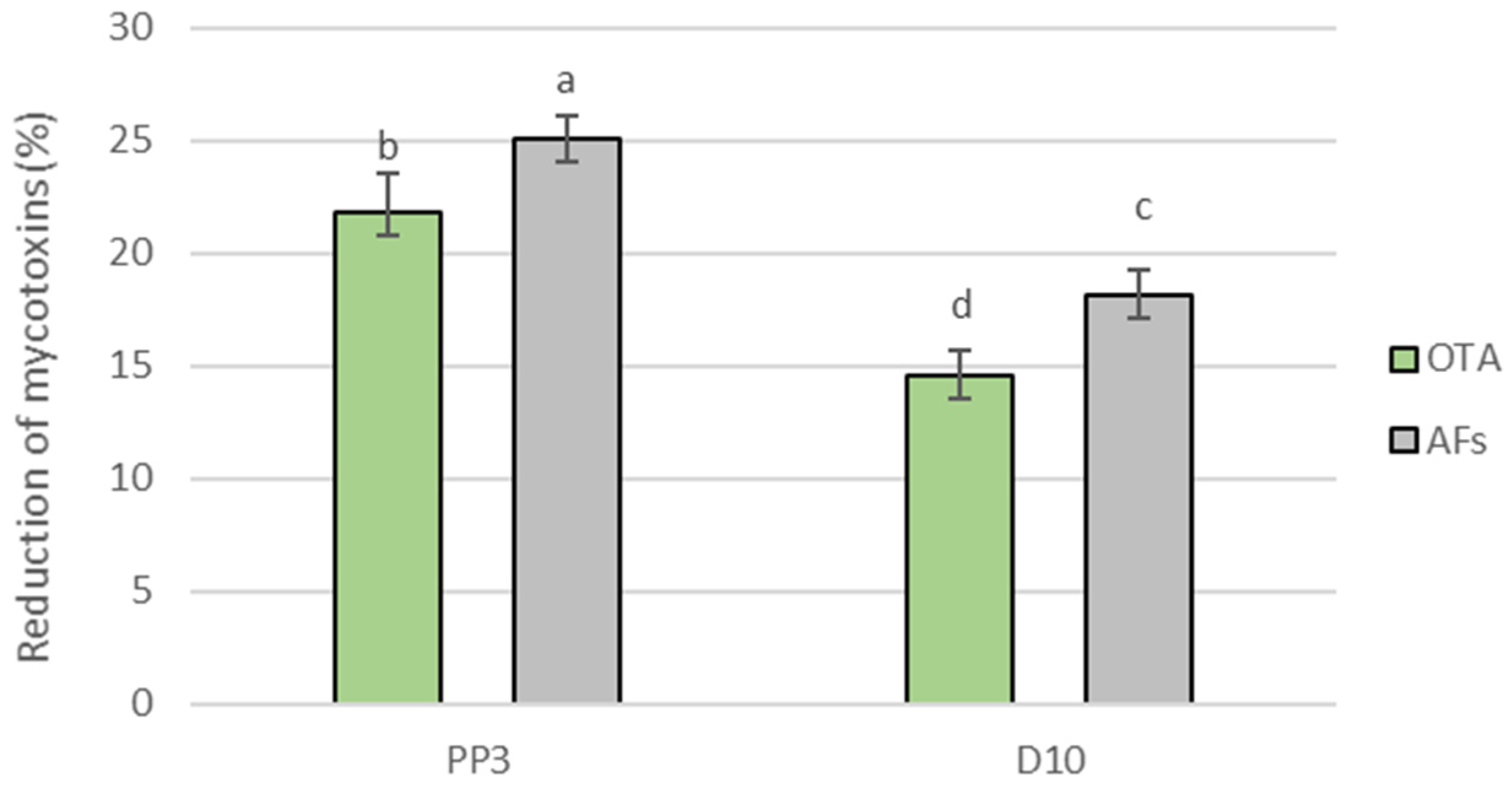
3.7. Determination of Mycotoxin Content in Model Bread
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kępińska-Pacelik, J.; Biel, W. Mycotoxins—Prevention, Detection, Impact on Animal Health. Processes 2021, 9, 2035. [Google Scholar] [CrossRef]
- Piotrowska, M. Wykorzystanie Mikroorganizmów Do Usuwania Mikotoksyn z Żywności i Pasz. Postępy Mikrobiol. 2012, 51, 109–119. [Google Scholar]
- Podgórska-Kryszczuk, I. Biological Control of Aspergillus Flavus by the Yeast Aureobasidium Pullulans In Vitro and on Tomato Fruit. Plants 2023, 12, 236. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef]
- Zahija, I.; Jeršek, B.; Demšar, L.; Polak, M.L.; Polak, T. Production of Aflatoxin B1 by Aspergillus parasiticus Grown on a Novel Meat-Based Media. Toxins 2023, 15, 25. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, G.; Yang, Q.; Yang, X.; Zheng, Y.; Liu, Y.; Xing, F. Effects of Light on the Ochratoxigenic Fungi Aspergillus ochraceus and A. carbonarius. Toxins 2021, 13, 251. [Google Scholar] [CrossRef]
- Escrivá, L.; Agahi, F.; Vila-Donat, P.; Mañes, J.; Meca, G.; Manyes, L. Bioaccessibility Study of Aflatoxin B1 and Ochratoxin A in Bread Enriched with Fermented Milk Whey and/or Pumpkin. Toxins 2022, 14, 6. [Google Scholar] [CrossRef]
- Nikolić, M.; Savić, I.; Nikolić, A.; Jauković, M.; Kandić, V.; Stevanović, M.; Stanković, S. Toxigenic Species Aspergillus parasiticus Originating from Maize Kernels Grown in Serbia. Toxins 2021, 13, 847. [Google Scholar] [CrossRef]
- Li, X.; Ma, W.; Ma, Z.; Zhang, Q.; Li, H. The Occurrence and Contamination Level of Ochratoxin a in Plant and Animal-Derived Food Commodities. Molecules 2021, 26, 6928. [Google Scholar] [CrossRef]
- Krska, R.; De Nijs, M.; McNerney, O.; Pichler, M.; Gilbert, J.; Edwards, S.; Suman, M.; Magan, N.; Rossi, V.; Van Der Fels-Klerx, H.J.; et al. Safe Food and Feed through an Integrated Toolbox for Mycotoxin Management: The MyToolBox Approach. World Mycotoxin J. 2016, 9, 487–495. [Google Scholar] [CrossRef]
- Helepciuc, F.E.; Todor, A. Improving the Authorization of Microbial Biological Control Products (MBCP) in the European Union within the EU Green Deal Framework. Agronomy 2022, 12, 1218. [Google Scholar] [CrossRef]
- Kowalska, G.; Kowalski, R. Pestycydy—Zakres i Ryzyko Stosowania, Korzyści i Zagrożenia. Praca Przeglądowa. Ann. Hortic. 2020, 29, 5–25. [Google Scholar] [CrossRef]
- Grzegorczyk, M.; Szalewicz, A.; Żarowska, B.; Połomska, X.; Wątorek, W.; Wojtatowicz, M. Drobnoustroje w Biologicznej Ochronie Roślin Przed Chorobami Grzybowymi. Acta Sci. Pol. Biotechnol. 2015, 14, 19–42. [Google Scholar]
- Muccilli, S.; Restuccia, C. Bioprotective Role of Yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef] [PubMed]
- Podgórska-Kryszczuk, I.; Solarska, E.; Kordowska-Wiater, M. Biological Control of Fusarium culmorum, Fusarium graminearum and Fusarium poae by Antagonistic Yeasts. Pathogens 2022, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Podgórska-Kryszczuk, I.; Solarska, E.; Kordowska-Wiater, M. Reduction of the Fusarium Mycotoxins: Deoxynivalenol, Nivalenol and Zearalenone by Selected Non-Conventional Yeast Strains in Wheat Grains and Bread. Molecules 2022, 27, 1578. [Google Scholar] [CrossRef]
- Repečkiene, J.; Levinskaite, L.; Paškevičius, A.; Raudoniene, V. Toxin-Producing Fungi on Feed Grains and Application of Yeasts for Their Detoxification. Pol. J. Vet. Sci. 2013, 16, 391–393. [Google Scholar] [CrossRef]
- Taheur, F.B.; Fedhila, K.; Chaieb, K.; Kouidhi, B.; Bakhrouf, A.; Abrunhosa, L. Adsorption of Aflatoxin B1, Zearalenone and Ochratoxin A by Microorganisms Isolated from Kefir Grains. Int. J. Food Microbiol. 2017, 251, 1–7. [Google Scholar] [CrossRef]
- Adebo, O.A.; Kayitesi, E.; Njobeh, P.B. Reduction of Mycotoxins during Fermentation of Whole Grain Sorghum to Whole Grain Ting (a Southern African Food). Toxins 2019, 11, 180. [Google Scholar] [CrossRef]
- Mozaffary, P.; Milani, J.M.; Heshmati, A. The Influence of Yeast Level and Fermentation Temperature on Ochratoxin A Decrement during Bread Making. Food Sci. Nutr. 2019, 7, 2144–2150. [Google Scholar] [CrossRef]
- Ramos, J.; Melero, Y.; Ramos-Moreno, L.; Michán, C.; Cabezas, L. Debaryomyces Hansenii Strains from Valle de Los Pedroches Iberian Dry Meat Products: Isolation, Identification, Characterization, and Selection for Starter Cultures. J. Microbiol. Biotechnol. 2017, 27, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Chan, Z.; Tian, S. Interaction of Antagonistic Yeasts against Postharvest Pathogens of Apple Fruit and Possible Mode of Action. Postharvest Biol. Technol. 2005, 36, 215–223. [Google Scholar] [CrossRef]
- Mamun-Or-Rashid, A.N.M.; Lucy, T.T.; Pramanik, M.K. Isolation, Identification, Optimization of Baker’s Yeast from Natural Sources, Scale-up Production Using Molasses as a Cheap Carbohydrate Source, and Evaluation for Bread Production. Appl. Microbiol. 2022, 2, 516–533. [Google Scholar] [CrossRef]
- Aslankoohi, E.; Herrera-Malaver, B.; Rezaei, M.N.; Steensels, J.; Courtin, C.M.; Verstrepen, K.J. Non-Conventional Yeast Strains Increase the Aroma Complexity of Bread. PLoS ONE 2016, 11, e0165126. [Google Scholar] [CrossRef]
- Escrivá, L.; Calpe, J.; Lafuente, C.; Moreno, A.; Musto, L.; Meca, G.; Luz, C. Aflatoxin B1 and Ochratoxin A Reduction by Lactobacillus spp. during Bread Making. J. Sci. Food Agric. 2023, 103, 7095–7103. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Zhang, Z.; Chen, Y.; Tian, S. Antagonistic Yeasts: A Promising Alternative to Chemical Fungicides for Controlling Postharvest Decay of Fruit. J. Fungi 2020, 6, 158. [Google Scholar] [CrossRef]
- Oztekin, S.; Dikmetas, D.N.; Devecioglu, D.; Acar, E.G.; Karbancioglu-Guler, F. Recent Insights into the Use of Antagonistic Yeasts for Sustainable Biomanagement of Postharvest Pathogenic and Mycotoxigenic Fungi in Fruits with Their Prevention Strategies against Mycotoxins. J. Agric. Food Chem. 2023, 71, 9923–9950. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Y.; Wang, Y.; Li, B.; Gu, X.; Zhang, X.; Boateng, N.A.S.; Zhang, H. Effect of β-Glucan on the Biocontrol Efficacy of Cryptococcus podzolicus against Postharvest Decay of Pears and the Possible Mechanisms Involved. Postharvest Biol. Technol. 2020, 160, 111057. [Google Scholar] [CrossRef]
- Alimadadi, N.; Pourvali, Z.; Nasr, S.; Fazeli, S.A.S. Screening of Antagonistic Yeast Strains for Postharvest Control of Penicillium expansum Causing Blue Mold Decay in Table Grape. Fungal Biol. 2023, 127, 901–908. [Google Scholar] [CrossRef]
- Zajc, J.; Černoša, A.; Di Francesco, A.; Castoria, R.; De Curtis, F.; Badri, H.; Jijakli, H.; Ippolito, A.; GostinČar, C.; Zalar, P.; et al. Characterization of Aureobasidium pullulans Isolates Selected as Biocontrol Agents against Fruit Decay Pathogens. Fungal Genom. Biol. 2020, 10, 163. [Google Scholar] [CrossRef]
- Galli, V.; Romboli, Y.; Barbato, D.; Mari, E.; Venturi, M.; Guerrini, S.; Granchi, L. Indigenous Aureobasidium pullulans Strains as Biocontrol Agents of Botrytis cinerea on Grape Berries. Sustainability 2021, 13, 9389. [Google Scholar] [CrossRef]
- Di Francesco, A.; Calassanzio, M.; Ratti, C.; Mari, M.; Folchi, A.; Baraldi, E. Molecular Characterization of the Two Postharvest Biological Control Agents Aureobasidium pullulans L1 and L8. Biol. Control 2018, 123, 53–59. [Google Scholar] [CrossRef]
- Zhang, D.; Spadaro, D.; Valente, S.; Garibaldi, A.; Gullino, M.L. Cloning, Characterization, Expression and Antifungal Activity of an Alkaline Serine Protease of Aureobasidium pullulans PL5 Involved in the Biological Control of Postharvest Pathogens. Int. J. Food Microbiol. 2012, 153, 453–464. [Google Scholar] [CrossRef]
- Banani, H.; Spadaro, D.; Zhang, D.; Matic, S.; Garibaldi, A.; Gullino, M.L. Biocontrol Activity of an Alkaline Serine Protease from Aureobasidium pullulans Expressed in Pichia pastoris against Four Postharvest Pathogens on Apple. Int. J. Food Microbiol. 2014, 182–183, 1–8. [Google Scholar] [CrossRef]
- Agirman, B.; Erten, H. Biocontrol Ability and Action Mechanisms of Aureobasidium pullulans GE17 and Meyerozyma guilliermondii KL3 against Penicillium digitatum DSM2750 and Penicillium expansum DSM62841 Causing Postharvest Diseases. Yeast 2020, 37, 437–448. [Google Scholar] [CrossRef]
- Ando, H.; Hatanaka, K.; Ohata, I.; Yamashita-Kitaguchi, Y.; Kurata, A.; Kishimoto, N. Antifungal Activities of Volatile Substances Generated by Yeast Isolated from Iranian Commercial Cheese. Food Control 2012, 26, 472–478. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiol. Spectr. 2017, 5, 267–292. [Google Scholar] [CrossRef]
- Pinto, C.; Custódio, V.; Nunes, M.; Songy, A.; Rabenoelina, F.; Courteaux, B.; Clément, C.; Gomes, A.C.; Fontaine, F. Understand the Potential Role of Aureobasidium pullulans, a Resident Microorganism from Grapevine, to Prevent the Infection Caused by Diplodia Seriata. Front. Microbiol. 2018, 9, 3047. [Google Scholar] [CrossRef]
- Gomes, F.C.O.; Safar, S.V.B.; Marques, A.R.; Medeiros, A.O.; Santos, A.R.O.; Carvalho, C.; Lachance, M.A.; Sampaio, J.P.; Rosa, C.A. The Diversity and Extracellular Enzymatic Activities of Yeasts Isolated from Water Tanks of Vriesea Minarum, an Endangered Bromeliad Species in Brazil, and the Description of Occultifur Brasiliensis f.a., Sp. Nov. Antonie Van Leeuwenhoek 2015, 107, 597–611. [Google Scholar] [CrossRef]
- Zajc, J.; Gostinčar, C.; Černoša, A.; Gunde-Cimerman, N. Stress-Tolerant Yeasts: Opportunistic Pathogenicity versus Biocontrol Potential. Genes 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Wisniewski, M.; Droby, S.; Liu, J. Responses of Yeast Biocontrol Agents to Environmental Stress. Appl. Environ. Microbiol. 2015, 81, 2968–2975. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, D.; Singh, R. Biological Control of Postharvest Diseases of Fruits and Vegetables by Microbial Antagonists: A Review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Moawad, A.S.; Bahr, H.S.; Abdelmohsen, U.R.; Mohammed, R. Natural Product Diversity from the Endophytic Fungi of the Genus Aspergillus. RSC Adv. 2020, 10, 22058–22079. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, S. Control of High Osmolarity Signalling in the Yeast Saccharomyces cerevisiae. FEBS Lett. 2009, 583, 4025–4029. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Semumu, T.; Gamero, A. Non-Conventional Yeasts as Alternatives in Modern Baking for Improved Performance and Aroma Enhancement. Fermentation 2021, 7, 102. [Google Scholar] [CrossRef]
- Musa Maryam, B.; Sambo Datsugwai Mohammed, S.; Abimbola Ayodeji, O.; Abimbola Ayodeji Screening, O. Screening of Fermentative Potency of Yeast Isolates from Iindigenous Sources for Dough Leavening. Int. J. Microbiol. Biotechnol. 2017, 2, 12. [Google Scholar] [CrossRef]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic Amines in Fermented Foods. Eur. J. Clin. Nutr. 2010, 64, 95–100. [Google Scholar] [CrossRef]
- Armando, M.R.; Dogi, C.A.; Poloni, V.; Rosa, C.A.R.; Dalcero, A.M.; Cavaglieri, L.R. In Vitro Study on the Effect of Saccharomyces cerevisiae Strains on Growth and Mycotoxin Production by Aspergillus carbonarius and Fusarium graminearum. Int. J. Food Microbiol. 2013, 161, 182–188. [Google Scholar] [CrossRef]
- Chlebicz, A.; Śliżewska, K. In Vitro Detoxification of Aflatoxin B1, Deoxynivalenol, Fumonisins, T-2 Toxin and Zearalenone by Probiotic Bacteria from Genus Lactobacillus and Saccharomyces cerevisiae Yeast. Probiotics Antimicrob. Proteins 2020, 12, 289. [Google Scholar] [CrossRef]
- Wei, M.; Dhanasekaran, S.; Legrand Ngolong Ngea, G.; Abiso Godana, E.; Zhang, X.; Yang, Q.; Zheng, X.; Zhang, H. Cryptococcus podzolicus Y3 Degrades Ochratoxin A by Intracellular Enzymes and Simultaneously Eliminates Citrinin. Biol. Control 2022, 168, 104857. [Google Scholar] [CrossRef]
- De Felice, D.V.; Solfrizzo, M.; De Curtis, F.; Lima, G.; Visconti, A.; Castoria, R. Strains of Aureobasidium pullulans Can Lower Ochratoxin A Contamination in Wine Grapes. Phytopathology 2008, 98, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Dimakopoulou, M.; Antoniou, P.P.; Tjamos, E.C. Biological Control Strategies of Mycotoxigenic Fungi and Associated Mycotoxins in Mediterranean Basin Crops. Phytopathol. Mediterr. 2012, 51, 158–174. [Google Scholar]
- Ndiaye, S.; Zhang, M.; Fall, M.; Ayessou, N.M.; Zhang, Q.; Li, P. Current Review of Mycotoxin Biodegradation and Bioadsorption: Microorganisms, Mechanisms, and Main Important Applications. Toxins 2022, 14, 729. [Google Scholar] [CrossRef] [PubMed]
- Bejaoul, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A Removal in Synthetic and Natural Grape Juices by Selected Oenological Saccharomyces Strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Yeast | Qualitative Determination (±) * | Quantitative Determination (U/mg Protein) | |||
|---|---|---|---|---|---|
| Protease | Cellulase | Amylase | Chitinase | β-1,3-Glucanase | |
| Aureobasidium pullulans PP3 | + | + | + | 1.79 ± 0.09 | 7.26 ± 0.12 |
| Saitozyma podzolicus D10 | − | − | + | 0.66 ± 0.1 | 5.04 ± 0.09 |
| Yeast Strain | ||||
|---|---|---|---|---|
| A. pullulans PP3 | S. podzolicus D10 | |||
| Growth conditions | Temperature (°C) | 4 | + | − |
| 14 | + | − | ||
| 18 | + | + | ||
| 22 | + | + | ||
| 30 | + | + | ||
| 32 | − | − | ||
| 37 | − | − | ||
| pH | 4 | + | + | |
| 5 | + | + | ||
| 6 | + | + | ||
| 7 | + | + | ||
| 9 | + | + | ||
| 11 | + | − | ||
| NaCl (%) | 0 | + | + | |
| 2 | + | + | ||
| 4 | + | + | ||
| 6 | + | − | ||
| 8 | + | − | ||
| 10 | + | − | ||
| 12 | + | − | ||
| 14 | − | − | ||
| H2S production | − | − | ||
| Biogenic amine production | − | − | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podgórska-Kryszczuk, I.; Pankiewicz, U.; Sas-Paszt, L. Biological Control of Aspergillus parasiticus and Aspergillus ochraceus and Reductions in the Amount of Ochratoxin A and Aflatoxins in Bread by Selected Non-Conventional Yeast. Foods 2023, 12, 3871. https://doi.org/10.3390/foods12203871
Podgórska-Kryszczuk I, Pankiewicz U, Sas-Paszt L. Biological Control of Aspergillus parasiticus and Aspergillus ochraceus and Reductions in the Amount of Ochratoxin A and Aflatoxins in Bread by Selected Non-Conventional Yeast. Foods. 2023; 12(20):3871. https://doi.org/10.3390/foods12203871
Chicago/Turabian StylePodgórska-Kryszczuk, Izabela, Urszula Pankiewicz, and Lidia Sas-Paszt. 2023. "Biological Control of Aspergillus parasiticus and Aspergillus ochraceus and Reductions in the Amount of Ochratoxin A and Aflatoxins in Bread by Selected Non-Conventional Yeast" Foods 12, no. 20: 3871. https://doi.org/10.3390/foods12203871
APA StylePodgórska-Kryszczuk, I., Pankiewicz, U., & Sas-Paszt, L. (2023). Biological Control of Aspergillus parasiticus and Aspergillus ochraceus and Reductions in the Amount of Ochratoxin A and Aflatoxins in Bread by Selected Non-Conventional Yeast. Foods, 12(20), 3871. https://doi.org/10.3390/foods12203871