
Influence of Benzothiadiazole on the Accumulation and Metabolism of C6 Compounds in Cabernet Gernischt Grapes (Vitis vinifera L.)

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Study Site
2.2. Reagents and Chemicals
2.3. Treatment and Sampling
2.4. Determination of Physicochemical Properties
2.5. Enzyme Activity
2.6. Sample Preparation for Aroma Compounds Analysis
2.7. Analysis of Grape Aroma Compounds
2.8. Analysis of Linolenic and Linoleic Acids
2.9. Statistical Analysis
3. Results and Discussion
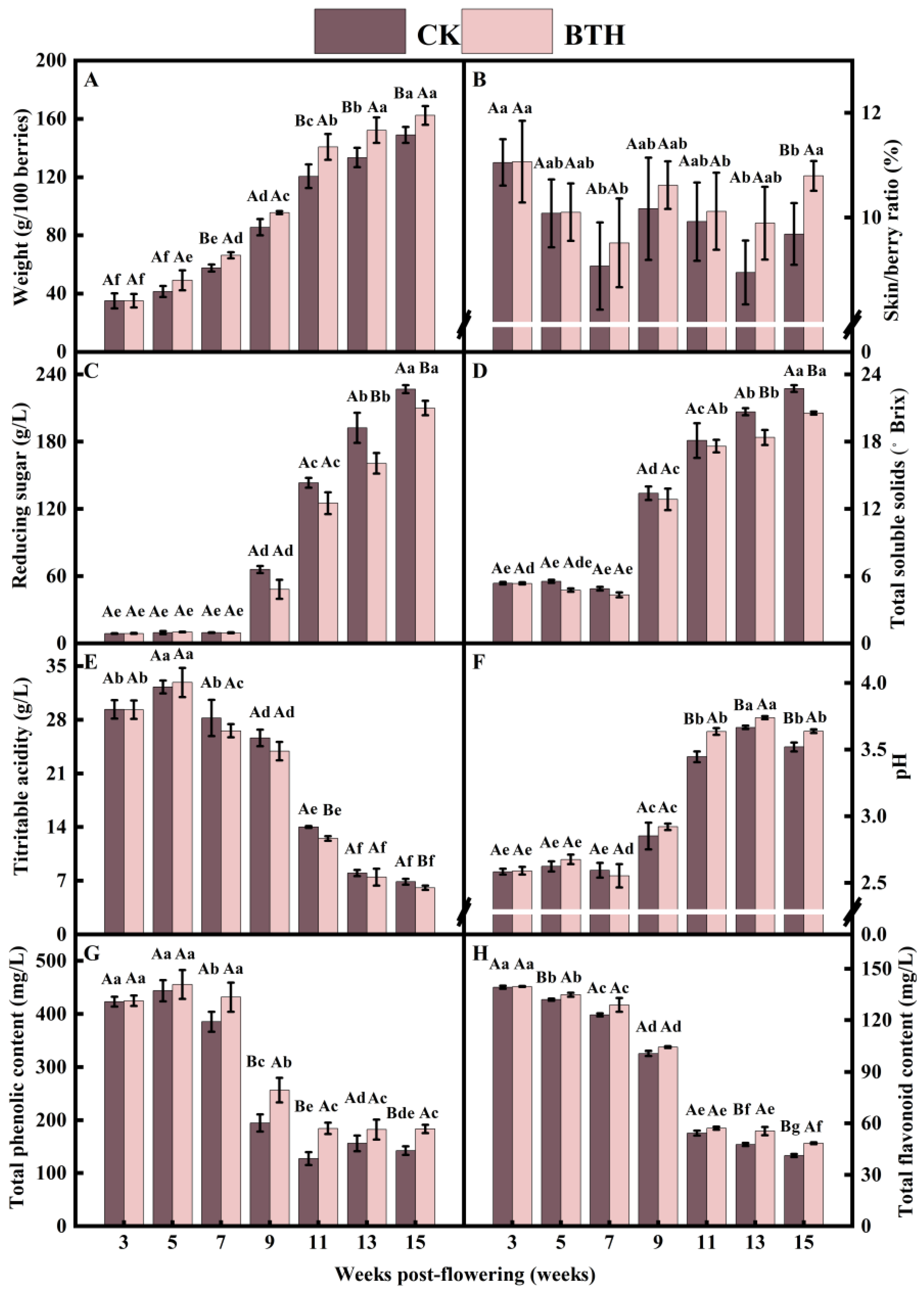
3.1. Influence of BTH Treatment on the Physicochemical Parameters of Cabernet Gernischt Grapes
3.1.1. 100-Berry Weight and Skin/Berry Ratio
3.1.2. Total Soluble Solids and Reducing Sugars
3.1.3. Titratable Acidity and pH
3.1.4. Total Phenolic (TP) and Total Flavonoid (TF) Content
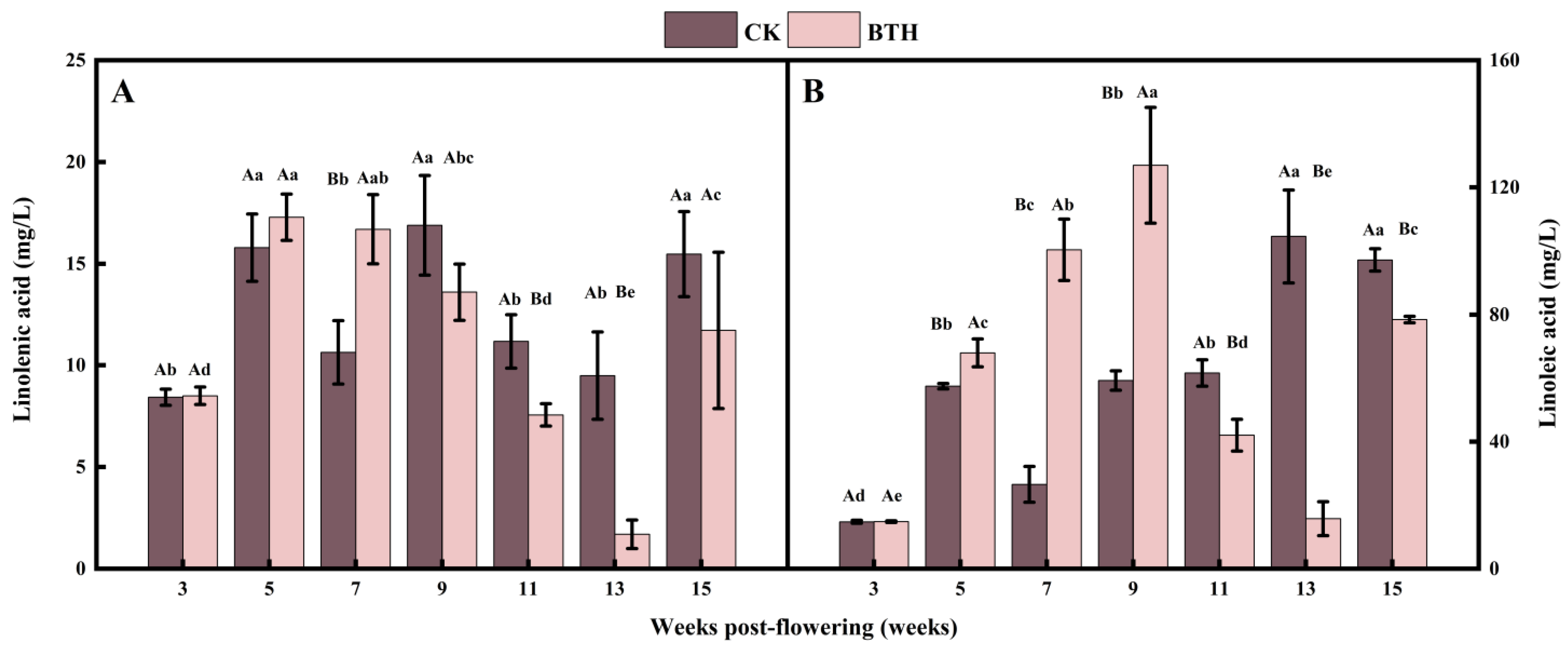
3.2. Effect of BTH on Linolenic and Linoleic Acids in Cabernet Gernischt Grapes
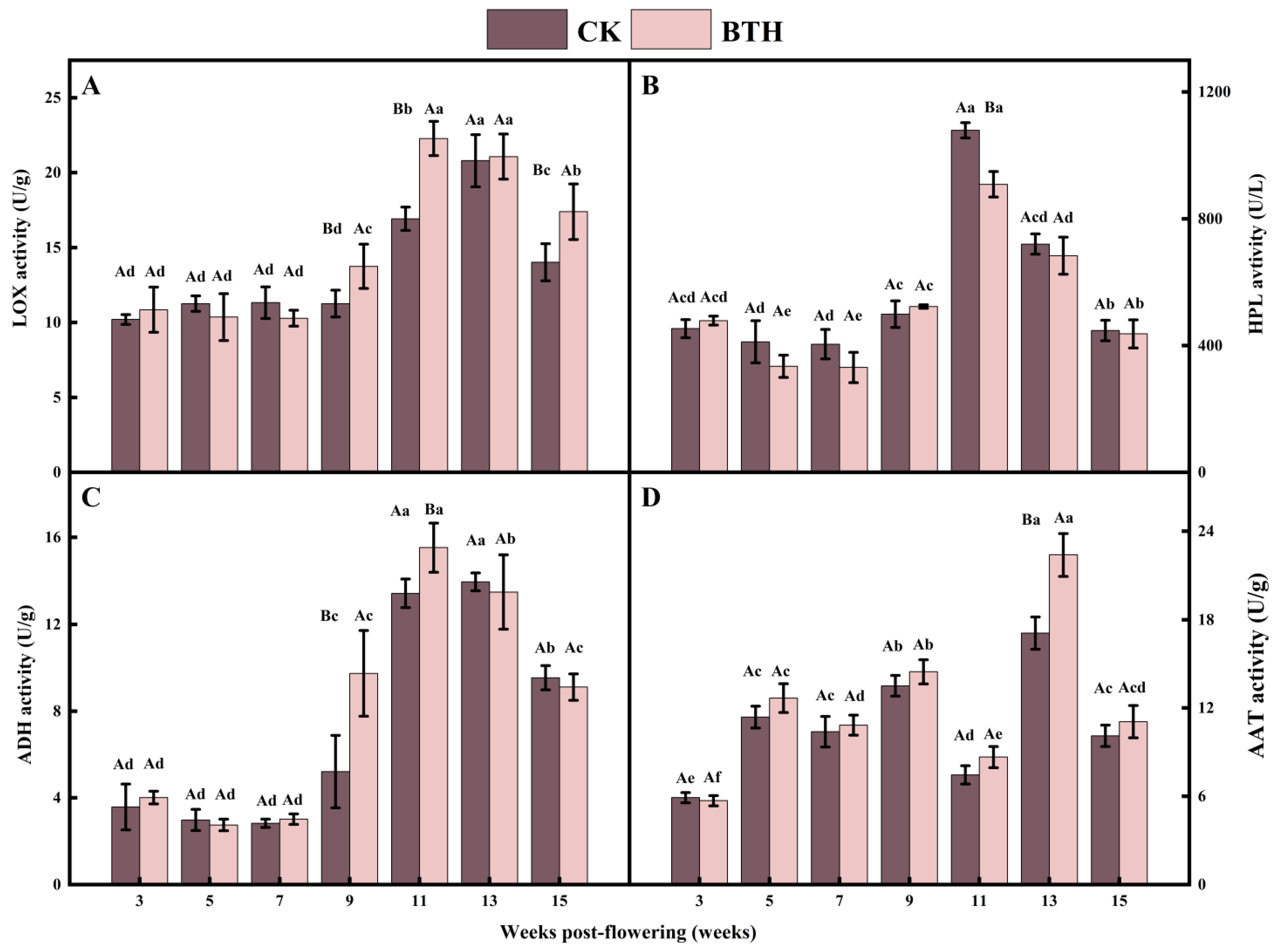
3.3. Effect of BTH Treatment on the Activities of Key Enzymes of LOX Pathway
3.4. Effect of BTH Treatment on the Concentration of C6 Aldehydes, Alcohols, and Esters in Cabernet Gernischt Grapes
3.4.1. C6 Aldehydes
3.4.2. C6 Alcohols
3.4.3. C6 Esters
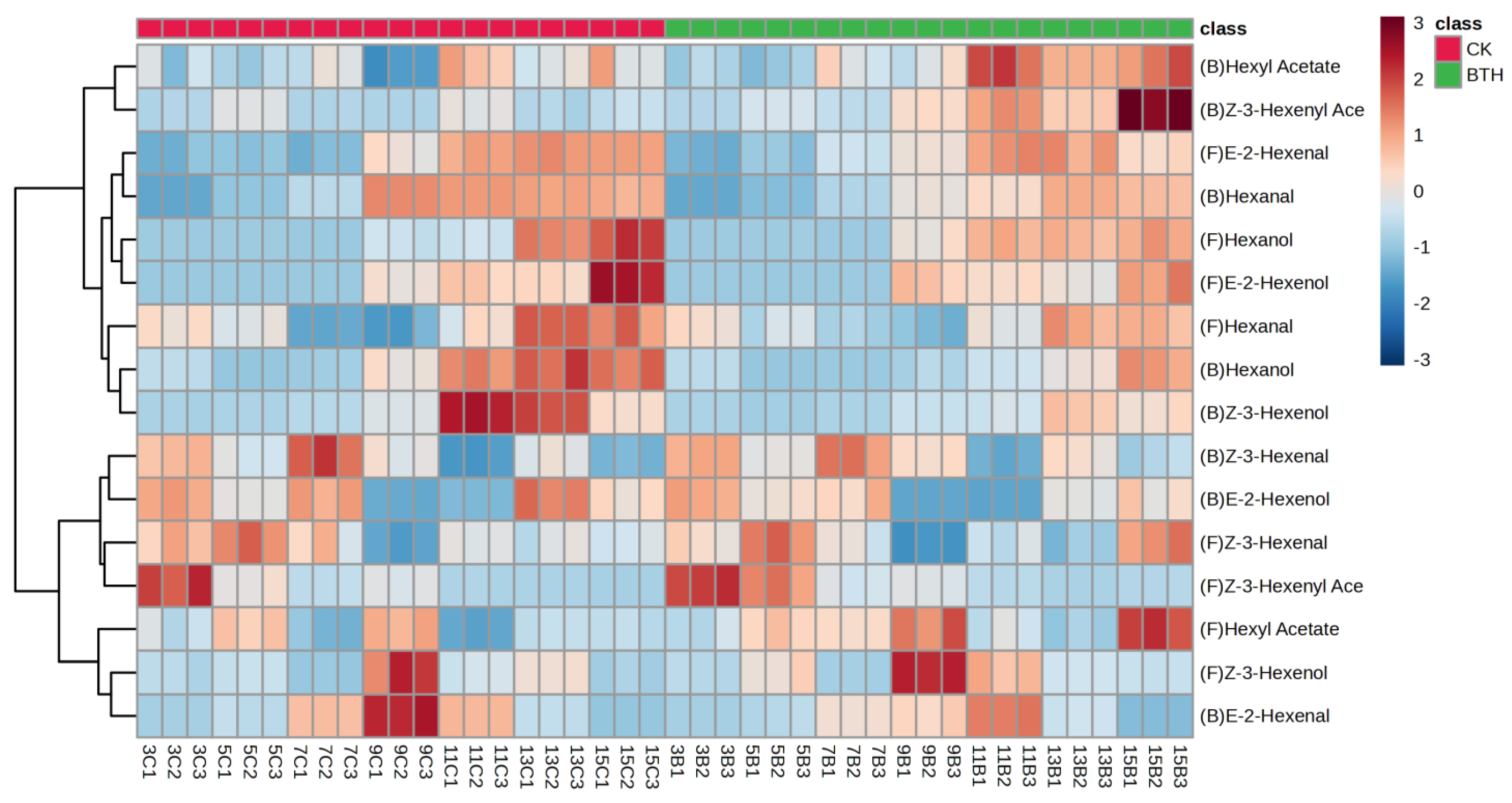
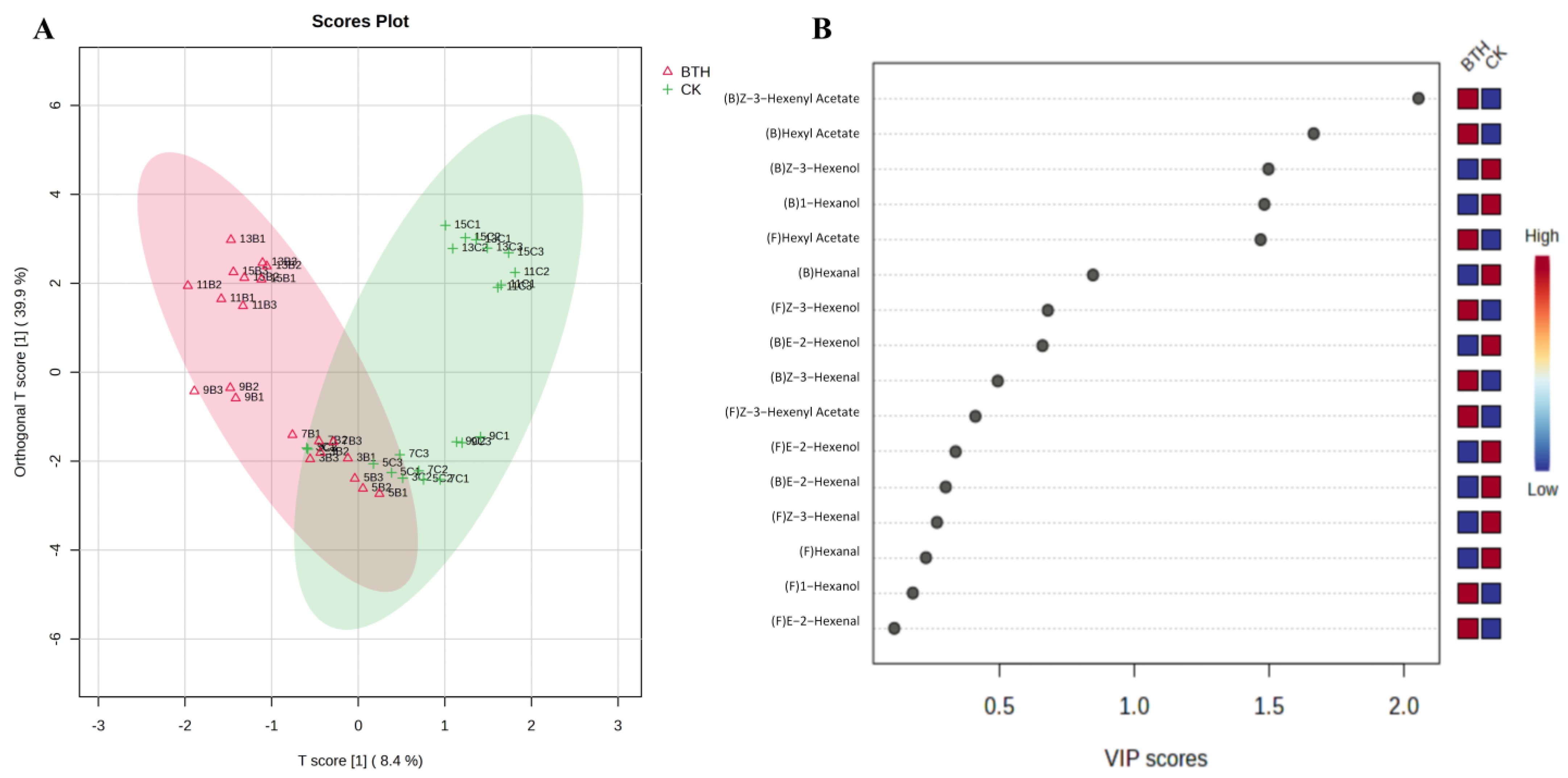
3.4.4. OPLS-DA Analysis of C6 Compounds in Cabernet Gernischt Grapes
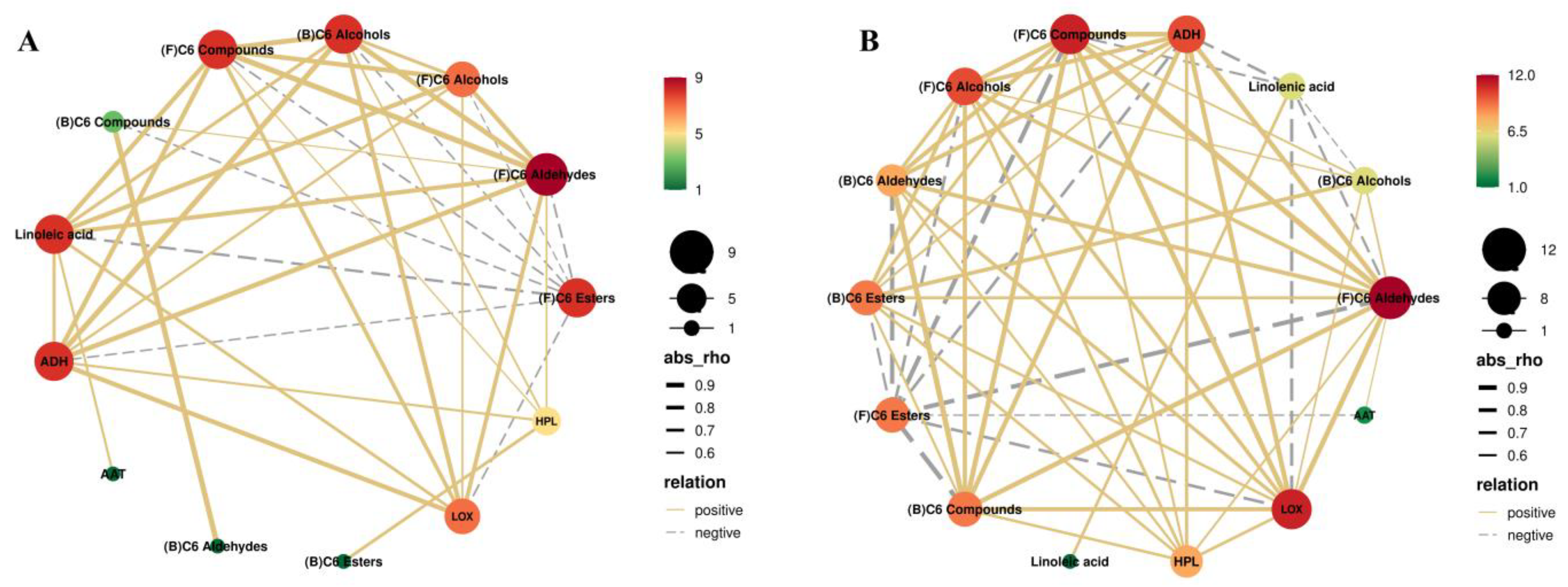
3.5. Pearson Correlation Analysis of Linoleic Acid, Linolenic Acid, Enzyme Activity, and C6 Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, Y.; Zhang, W.; Song, S.; Xu, W.; Zhang, C.; Ma, C.; Wang, L.; Wang, S. Evolution of Volatile Compounds during the Development of Muscat Grape ‘Shine Muscat’ (Vitis labrusca × V. Vinifera). Food Chem. 2020, 309, 125778. [Google Scholar] [CrossRef] [PubMed]
- Alem, H.; Rigou, P.; Schneider, R.; Ojeda, H.; Torregrosa, L. Impact of Agronomic Practices on Grape Aroma Composition: A Review. J. Sci. Food Agric. 2019, 99, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Lund, C.M.; Thompson, M.K.; Benkwitz, F.; Wohler, M.W.; Triggs, C.M.; Gardner, R.; Heymann, H.; Nicolau, L. New Zealand Sauvignon Blanc Distinct Flavor Characteristics: Sensory, Chemical, and Consumer Aspects. Am. J. Enol. Vitic. 2009, 60, 1–12. [Google Scholar] [CrossRef]
- Roland, A.; Vialaret, J.; Razungles, A.; Rigou, P.; Schneider, R. Evolution of S-Cysteinylated and s-Glutathionylated Thiol Precursors during Oxidation of Melon B. and Sauvignon Blanc Musts. J. Agric. Food Chem. 2010, 58, 4406–4413. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K. Green Leaf Volatiles: Hydroperoxide Lyase Pathway of Oxylipin Metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Costabel, M.P.; Wilkinson, K.L.; Bastian, S.E.P.; McCarthy, M.; Ford, C.M.; Dokoozlian, N. Seasonal and Regional Variation of Green Aroma Compounds in Commercial Vineyards of Vitis vinifera L. Merlot in California. Am. J. Enol. Vitic. 2013, 64, 430–436. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Faria, M.; Sá, F.; Barros, F.; Araújo, I.M. C6-Alcohols as Varietal Markers for Assessment of Wine Origin. Anal. Chim. Acta 2006, 563, 300–309. [Google Scholar] [CrossRef]
- Fang, Y.; Qian, M.C. Development of C6 and Other Volatile Compounds in Pinot Noir Grapes Determined by Stir Bar Sorptive Extraction-GC-MS. ACS Symp. Ser. 2012, 1104, 81–99. [Google Scholar] [CrossRef]
- Yuan, F.; Qian, M.C. Development of C13-Norisoprenoids, Carotenoids and Other Volatile Compounds in Vitis Vinifera L. Cv. Pinot Noir Grapes. Food Chem. 2016, 192, 633–641. [Google Scholar] [CrossRef]
- Gamero, E.; Moreno, D.; Vilanova, M.; Uriarte, D.; Prieto, M.H.; Valdés, M.E. Effect of Bunch Thinning and Water Stress on Chemical and Sensory Characteristics of Tempranillo Wines. Aust. J. Grape Wine Res. 2014, 20, 394–400. [Google Scholar] [CrossRef]
- Robinson, A.L.; Boss, P.K.; Heymann, H.; Solomon, P.S.; Trengove, R.D. Influence of Yeast Strain, Canopy Management, and Site on the Volatile Composition and Sensory Attributes of Cabernet Sauvignon Wines from Western Australia. J. Agric. Food Chem. 2011, 59, 3273–3284. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Cheng, G.; Duan, L.; Jiang, R.; Pan, Q.; Duan, C.; Wang, J. Effect of Training Systems on Fatty Acids and Their Derived Volatiles in Cabernet Sauvignon Grapes and Wines of the North Foot of Mt. Tianshan. Food Chem. 2015, 181, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Zoecklein, B.W.; Wolf, T.K.; Pélanne, L.; Miller, M.K.; Birkenmaier, S.S. Effect of Vertical Shoot-Positioned, Smart-Dyson, and Geneva Double-Curtain Training Systems on Viognier Grape and Wine Composition. Am. J. Enol. Vitic. 2008, 59, 11–21. [Google Scholar] [CrossRef]
- Mendez-Costabel, M.P.; Wilkinson, K.L.; Bastian, S.E.P.; Jordans, C.; Mccarthy, M.; Ford, C.M.; Dokoozlian, N. Effect of Winter Rainfall on Yield Components and Fruit Green Aromas of Vitis vinifera L. Cv. Merlot in California. Aust. J. Grape Wine Res. 2014, 20, 100–110. [Google Scholar] [CrossRef]
- Jiang, Y.; Sam, F.E.; Li, J.; Bi, Y.; Ma, T.; Zhang, B. Pre-Harvest Benzothiadiazole Spraying Promotes the Cumulation of Phenolic Compounds in Grapes. Foods 2022, 11, 3345. [Google Scholar] [CrossRef]
- Yue, X.; Ju, Y.; Zhang, T.; Yu, R.; Xu, H.; Zhang, Z. Application of Salicylic Acid to Cv. Muscat Hamburg Grapes for Quality Improvement: Effects on Typical Volatile Aroma Compounds and Anthocyanin Composition of Grapes and Wines. LWT 2023, 182, 114828. [Google Scholar] [CrossRef]
- Lisov, N.; Čakar, U.; Milenković, D.; Čebela, M.; Vuković, G.; Despotović, S.; Petrović, A. The Influence of Cabernet Sauvignon Ripeness, Healthy State and Maceration Time on Wine and Fermented Pomace Phenolic Profile. Fermentation 2023, 9, 695. [Google Scholar] [CrossRef]
- Paladines-Quezada, D.F.; Moreno-Olivares, J.D.; Bleda-Sánchez, J.A.; Gil-Muñoz, R.; Fernández-Fernández, J.I. Application of Elicitors at Two Maturation Stages of Vitis vinifera L. Cv Monastrell: Changes in Skin Cell Walls. Chemistry 2022, 4, 98–111. [Google Scholar] [CrossRef]
- Paladines-Quezada, D.F.; Moreno-Olivares, J.D.; Fernández-Fernández, J.I.; Bautista-Ortín, A.B.; Gil-Muñoz, R. Influence of Methyl Jasmonate and Benzothiadiazole on the Composition of Grape Skin Cell Walls and Wines. Food Chem. 2019, 277, 691–697. [Google Scholar] [CrossRef]
- Kok, D. Influences of Pre- and Post-Veraison Cluster Thinning Treatments on Grape Composition Variables and Monoterpene Levels of Vitis vinifera L. Cv. Sauvignon Blanc. J. Food Agric. Environ. 2011, 9, 22–26. [Google Scholar]
- Miliordos, D.E.; Tsiknia, M.; Kontoudakis, N.; Dimopoulou, M.; Bouyioukos, C.; Kotseridis, Y. Impact of Application of Abscisic Acid, Benzothiadiazole and Chitosan on Berry Quality Characteristics and Plant Associated Microbial Communities of Vitis vinifera L Var. Mouhtaro Plants. Sustainability 2021, 13, 5802. [Google Scholar] [CrossRef]
- Salifu, R. Influence of Fertilizer and Benzothiadiazole (BTH) Treatments on the Aroma Profile of Red Grapes Application. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2022. [Google Scholar]
- Wang, K.; Liao, Y.; Cao, S.; Di, H.; Zheng, Y. Effects of Benzothiadiazole on Disease Resistance and Soluble Sugar Accumulation in Grape Berries and Its Possible Cellular Mechanisms Involved. Postharvest Biol. Technol. 2015, 102, 51–60. [Google Scholar] [CrossRef]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar Sensing and Signaling in Plants: Conserved and Novel Mechanisms. Annu. Rev. Plant. Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [PubMed]
- Paladines-Quezada, D.F.; Fernández-Fernández, J.I.; Moreno-Olivares, J.D.; Bleda-Sánchez, J.A.; Gómez-Martínez, J.C.; Martínez-Jiménez, J.A.; Gil-Muñoz, R. Application of Elicitors in Two Ripening Periods of Vitis vinifera L. Cv Monastrell: Influence on Anthocyanin Concentration of Grapes and Wines. Molecules 2021, 26, 1689. [Google Scholar] [CrossRef]
- Gómez-Plaza, E.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gil-Muñoz, R. Effect of Elicitors on the Evolution of Grape Phenolic Compounds during the Ripening Period. J. Sci. Food Agric. 2017, 97, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Paladines-Quezada, D.F.; Moreno-Olivares, J.D.; Fernández-Fernández, J.I.; Bleda-Sánchez, J.A.; Martínez-Moreno, A.; Gil-Muñoz, R. Elicitors and Pre-Fermentative Cold Maceration: Effects on Polyphenol Concentration in Monastrell Grapes and Wines. Biomolecules 2019, 9, 671. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gómez-Plaza, E. Improving Phenolic and Chromatic Characteristics of Monastrell, Merlot and Syrah Wines by Using Methyl Jasmonate and Benzothiadiazole. Oeno One 2017, 51, 17–27. [Google Scholar] [CrossRef]
- Garcia-Brugger, A.; Lamotte, O.; Vandelle, E.; Bourque, S.; Lecourieux, D.; Poinssot, B.; Wendehenne, D.; Pugin, A. Early Signaling Events Induced by Elicitors of Plant Defenses. Mol. Plant-Microbe Interact. 2006, 19, 711–724. [Google Scholar] [CrossRef]
- Li, X.; Bi, Y.; Wang, J.; Dong, B.; Li, H.; Gong, D.; Zhao, Y.; Tang, Y.; Yu, X.; Shang, Q. BTH Treatment Caused Physiological, Biochemical and Proteomic Changes of Muskmelon (Cucumis melo L.) Fruit during Ripening. J. Proteom. 2015, 120, 179–193. [Google Scholar] [CrossRef]
- Gómez-Plaza, E.; Mestre-Ortuño, L.; Ruiz-García, Y.; Fernández-Fernández, J.I.; López-Roca, J.M. Effect of Benzothiadiazole and Methyl Jasmonate on the Volatile Compound Composition of Vitis vinifera L. Monastrell Grapes and Wines. Am. J. Enol. Vitic. 2012, 63, 394–401. [Google Scholar] [CrossRef]
- Cao, S.; Hu, Z.; Zheng, Y.; Lu, B. Effect of BTH on Anthocyanin Content and Activities of Related Enzymes in Strawberry after Harvest. J. Agric. Food Chem. 2010, 58, 5801–5805. [Google Scholar] [CrossRef]
- Gong, D.; Bi, Y.; Zhang, X.; Han, Z.; Zong, Y.; Li, Y.; Sionov, E.; Prusky, D. Benzothiadiazole Treatment Inhibits Membrane Lipid Metabolism and Straight-Chain Volatile Compound Release in Penicillium expansum-Inoculated Apple Fruit. Postharvest Biol. Technol. 2021, 181, 111671. [Google Scholar] [CrossRef]
- Ju, Y.; Liu, M.; Tu, T.; Zhao, X.; Yue, X.; Zhang, J.; Fang, Y.; Meng, J. Effect of Regulated Deficit Irrigation on Fatty Acids and Their Derived Volatiles in ‘Cabernet Sauvignon’ Grapes and Wines of Ningxia, China. Food Chem. 2018, 245, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Kalua, C.M.; Boss, P.K. Evolution of Volatile Compounds during the Development of Cabernet Sauvignon Grapes (Vitis vinifera L.). J. Agric. Food Chem. 2009, 57, 3818–3830. [Google Scholar] [CrossRef] [PubMed]
- Naziya, B.; Murali, M.; Amruthesh, K.N. Plant Growth-Promoting Fungi (Pgpf) Instigate Plant Growth and Induce Disease Resistance in Capsicum annuum L. upon Infection with Colletotrichum capsici (Syd.) Butler & Bisby. Biomolecules 2020, 10, 41. [Google Scholar] [CrossRef]
- Valero, D.; Díaz-Mula, H.M.; Zapata, P.J.; Castillo, S.; Guillén, F.; Martínez-Romero, D.; Serrano, M. Postharvest Treatments with Salicylic Acid, Acetylsalicylic Acid or Oxalic Acid Delayed Ripening and Enhanced Bioactive Compounds and Antioxidant Capacity in Sweet Cherry. J. Agric. Food Chem. 2011, 59, 5483–5489. [Google Scholar] [CrossRef]
- Zhao, Y.; Song, C.; Brummell, D.A.; Qi, S.; Lin, Q.; Bi, J.; Duan, Y. Salicylic Acid Treatment Mitigates Chilling Injury in Peach Fruit by Regulation of Sucrose Metabolism and Soluble Sugar Content. Food Chem. 2021, 358, 129867. [Google Scholar] [CrossRef]
- Hu, B.; Gao, J.; Xu, S.; Zhu, J.; Fan, X.; Zhou, X. Quality Evaluation of Different Varieties of Dry Red Wine Based on Nuclear Magnetic Resonance Metabolomics. Appl. Biol. Chem. 2020, 63, 24. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Wang, B.; Han, Y.; Li, Y.; Prusky, D.; Bi, Y. Soluble Sugars, Organic Acids and Energy Metabolism Involved in the Wound Healing of Muskmelons Elicited by Benzothiadiazole. Postharvest Biol. Technol. 2023, 199, 112277. [Google Scholar] [CrossRef]
- Trefely, S.; Lovell, C.D.; Snyder, N.W.; Wellen, K.E. Compartmentalised Acyl-CoA Metabolism and Roles in Chromatin Regulation. Mol. Metab. 2020, 38, 100941. [Google Scholar] [CrossRef]
- Choi, I.; Son, H.; Baek, J.H. Tricarboxylic Acid (Tca) Cycle Intermediates: Regulators of Immune Responses. Life 2021, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Castrejón, A.D.R.; Eichholz, I.; Rohn, S.; Kroh, L.W.; Huyskens-Keil, S. Phenolic Profile and Antioxidant Activity of Highbush Blueberry (Vaccinium corymbosum L.) during Fruit Maturation and Ripening. Food Chem. 2008, 109, 564–572. [Google Scholar] [CrossRef]
- Fumagalli, F.; Rossoni, M.; Iriti, M.; Di Gennaro, A.; Faoro, F.; Borroni, E.; Borgo, M.; Scienza, A.; Sala, A.; Folco, G. From Field to Health: A Simple Way to Increase the Nutraceutical Content of Grape as Shown by NO-Dependent Vascular Relaxation. J. Agric. Food Chem. 2006, 54, 5344–5349. [Google Scholar] [CrossRef]
- Hu, Y.; Li, J.; Wang, Y.; Wang, B.; Zhang, G.; Zhang, R.; Jiang, Y. Effect of Postharvest Benzothiadiazole Treatment on Membrane Phospholipid Metabolism of Thick-Skinned Muskmelon (in Chinese with English Abstract). Food Sci. 2018, 39, 165–173. [Google Scholar] [CrossRef]
- Ullah, C.; Tsai, C.J.; Unsicker, S.B.; Xue, L.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Salicylic Acid Activates Poplar Defense against the Biotrophic Rust Fungus Melampsora Larici-Populina via Increased Biosynthesis of Catechin and Proanthocyanidins. New Phytol. 2019, 221, 960–975. [Google Scholar] [CrossRef]
- Fang, F.; Huang, W.D. Salicylic Acid Modulated Flavonol Biosynthesis in Three Key Phases during Grape Berry Development. Eur. Food Res. Technol. 2013, 237, 441–448. [Google Scholar] [CrossRef]
- Liang, C.; Yang, B.; Wei, Y.; Zhang, P.; Wen, P. SA Incubation Induced Accumulation of Flavan-3-Ols through Activated VvANR Expression in Grape Leaves. Sci. Hortic. 2021, 287, 110269. [Google Scholar] [CrossRef]
- Iriti, M.; Rossoni, M.; Borgo, M.; Faoro, F. Benzothiadiazole Enhances Resveratrol and Anthocyanin Biosynthesis in Grapevine, Meanwhile Improving Resistnce to Botrytis Cinerea. J. Agric. Food Chem. 2004, 52, 4406–4413. [Google Scholar] [CrossRef]
- Gozzo, F. Systemic Acquired Resistance in Crop Protection. Outlooks Pest. Manag. 2004, 15, 20–23. [Google Scholar] [CrossRef]
- Gao, Q.M.; Venugopal, S.; Navarre, D.; Kachroo, A. Low Oleic Acid-Derived Repression of Jasmonic Acid-Inducible Defense Responses Requires the WRKY50 and WRKY51 Proteins. Plant. Physiol. 2011, 155, 464–476. [Google Scholar] [CrossRef]
- Upchurch, R.G. Fatty Acid Unsaturation, Mobilization, and Regulation in the Response of Plants to Stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Shemi, R.G.M.I. Effects of the Foliar-Applied Salicylic Acid, Zinc, and Glycine Betaine in Improving Drought-Tolerance of Wheat and Maize during Critical Growth Stages. Ph.D. Thesis, Southwest University, Chongqing, China, 2020. [Google Scholar]
- Deytieux, C.; Geny, L.; Lapaillerie, D.; Claverol, S.; Bonneu, M.; Donèche, B. Proteome Analysis of Grape Skins during Ripening. J. Exp. Bot. 2007, 58, 1851–1862. [Google Scholar] [CrossRef] [PubMed]
- OuYang, Y.N.; Gao, J.S.; Li, R.L.; Zhu, M.R.; Hu, X.H.; Min, Z.; Chen, S.X.; Zhang, Z.W.; Fang, Y.L. Changes in Volatile Profiles and Activity of Hydroperoxide Lyase and Alcohol Dehydrogenase during the Development of Cabernet Sauvignon Grapes (Vitis vinifera L.). S. Afr. J. Enol. Vitic. 2015, 36, 328–342. [Google Scholar] [CrossRef]
- Vancanneyt, G.; Sanz, C.; Farmaki, T.; Paneque, M.; Ortego, F.; Castañera, P.; Sánchez-Serrano, J.J. Hydroperoxide Lyase Depletion in Transgenic Potato Plants Leads to an Increase in Aphid Performance. Proc. Natl. Acad. Sci. USA 2001, 98, 8139–8144. [Google Scholar] [CrossRef] [PubMed]
- Bate, N.J.; Sivasankar, S.; Moxon, C.; Riley, J.M.C.; Thompson, J.E.; Rothstein, S.J. Molecular Characterization of an Arabidopsis Gene Encoding Hydroperoxide Lyase, a Cytochrome P-450 That Is Wound Inducible. Plant Physiol. 1998, 117, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.A.; Lee, G.I.; Itoh, A.; Li, L.; DeRocher, A.E. Cytochrome P450-Dependent Metabolism of Oxylipins in Tomato. Cloning and Expression of Allene Oxide Synthase and Fatty Acid Hydroperoxide Lyase. Plant Physiol. 2000, 123, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Díaz de León-Sánchez, F.; Pelayo-Zaldívar, C.; Rivera-Cabrera, F.; Ponce-Valadez, M.; Ávila-Alejandre, X.; Fernández, F.J.; Escalona-Buendía, H.B.; Pérez-Flores, L.J. Effect of Refrigerated Storage on Aroma and Alcohol Dehydrogenase Activity in Tomato Fruit. Postharvest Biol. Technol. 2009, 54, 93–100. [Google Scholar] [CrossRef]
- Defilippi, B.G.; Kader, A.A.; Dandekar, A.M. Apple Aroma: Alcohol Acyltransferase, a Rate Limiting Step for Ester Biosynthesis, Is Regulated by Ethylene. Plant Sci. 2005, 168, 1199–1210. [Google Scholar] [CrossRef]
- Kumar, N.; Tokas, J.; Raghavendra, M.; Singal, H.R. Impact of Exogenous Salicylic Acid Treatment on the Cell Wall Metabolism and Ripening Process in Postharvest Tomato Fruit Stored at Ambient Temperature. Int. J. Food Sci. Technol. 2021, 56, 2961–2972. [Google Scholar] [CrossRef]
- Madžgalj, V.; Petrović, A.; Čakar, U.; Maraš, V.; Sofrenić, I.; Tešević, V. The Influence of Different Enzymatic Preparations and Skin Contact Time on Aromatic Profile of Wines Produced from Autochthonous Grape Varieties Krstač and Žižak. J. Serb. Chem. Soc. 2022, 88, 11–23. [Google Scholar] [CrossRef]
- Salinas, M.R.; Zalacain, A.; Pardo, F.; Alonso, G.L. Stir Bar Sorptive Extraction Applied to Volatile Constituents Evolution during Vitis Vinifera Ripening. J. Agric. Food Chem. 2004, 52, 4821–4827. [Google Scholar] [CrossRef] [PubMed]
- Feussner, I.; Wasternack, C. The Lipoxygenase Pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Ma, L.; Hao, H.; Yu, X. Volatile Precursors and Aroma-Related Enzyme Activities during Fruit Maturation of Muskmelon. Fresh Prod. 2011, 5, 77–79. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Han, Y.; Chen, C.; Sam, F.E.; Guan, R.; Wang, K.; Zhang, Y.; Zhao, M.; Chen, C.; Liu, X.; et al. Influence of Benzothiadiazole on the Accumulation and Metabolism of C6 Compounds in Cabernet Gernischt Grapes (Vitis vinifera L.). Foods 2023, 12, 3710. https://doi.org/10.3390/foods12193710
Wang J, Han Y, Chen C, Sam FE, Guan R, Wang K, Zhang Y, Zhao M, Chen C, Liu X, et al. Influence of Benzothiadiazole on the Accumulation and Metabolism of C6 Compounds in Cabernet Gernischt Grapes (Vitis vinifera L.). Foods. 2023; 12(19):3710. https://doi.org/10.3390/foods12193710
Chicago/Turabian StyleWang, Jianfeng, Yuqi Han, Chunxia Chen, Faisal Eudes Sam, Ruwen Guan, Kai Wang, Yu Zhang, Man Zhao, Changxia Chen, Xuan Liu, and et al. 2023. "Influence of Benzothiadiazole on the Accumulation and Metabolism of C6 Compounds in Cabernet Gernischt Grapes (Vitis vinifera L.)" Foods 12, no. 19: 3710. https://doi.org/10.3390/foods12193710
APA StyleWang, J., Han, Y., Chen, C., Sam, F. E., Guan, R., Wang, K., Zhang, Y., Zhao, M., Chen, C., Liu, X., & Jiang, Y. (2023). Influence of Benzothiadiazole on the Accumulation and Metabolism of C6 Compounds in Cabernet Gernischt Grapes (Vitis vinifera L.). Foods, 12(19), 3710. https://doi.org/10.3390/foods12193710