
Phenotypic and Genotypic Characteristics of Non-Hemolytic L. monocytogenes Isolated from Food and Processing Environments
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Reference Strains
2.3. Isolation and Identification of L. monocytogenes
2.4. Storage and Revival of Reference LM Strains and aLM Isolates
2.5. DNA Isolation
2.6. Multiplex PCR Molecular Serotyping
2.7. Gene Virulence and Specific Gene for L. monocytogenes
2.8. Electrophoresis of Multiplex PCR and PCR Reaction Products
2.9. Antibiotic-Resistant L. monocytogenes
2.10. Statistical Analysis
3. Results and Discussion
3.1. Isolation Environment of Non-Hemolytic L. monocytogenes
3.2. Phenotypic Characteristics
3.3. Ability to Ferment Sugars

{kind=link}
{kind=link}
{kind=link}
| Sample | D-arabitol | Arabinose | Galactose | Glucose-1-Phosphate | Glucose | Lactose | D- Mannitol | Methyl-L-D-Glucoside | Rhamnose | Ribose | Sorbitol | Sucrose | D -Tagatose | D-Xylose |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1/2a | − | − | + | − | + | + | + | + | + | − | − | + | − | − |
| 1/2b | − | − | + | − | + | + | + | + | + | − | − | + | − | − |
| 1/2c | − | − | + | − | + | + | − | + | + | − | − | + | − | − |
| 3a | − | − | + | − | + | + | − | + | + | − | − | + | − | − |
| 3b | − | − | + | − | + | + | + | + | + | − | − | + | − | − |
| 3c | − | − | + | − | + | + | − | + | + | − | − | + | − | − |
| 4a | − | − | + | − | + | + | − | + | + | − | − | + | − | − |
| 4b | − | − | + | − | + | + | − | + | + | − | − | + | − | − |
| 4c | − | − | + | − | + | + | + | + | + | − | − | + | − | − |
| 4d | − | − | + | − | + | + | − | + | + | − | − | + | − | − |
| 4e | − | − | + | − | + | + | − | + | + | − | − | + | − | − |
| 7 | − | − | + | − | + | + | + | + | + | − | − | + | − | − |
| 112 | + | − | − | − | − | + | − | + | + | − | − | + | − | − |
| 245 | + | − | − | − | − | + | − | + | + | − | − | + | − | + |
| 273 | + | − | − | − | − | + | + | − | + | + | − | + | − | − |
| 275 | + | − | − | − | − | + | − | + | + | − | + | + | − | − |
| 280 | + | − | − | − | + | − | + | − | + | + | − | + | − | − |
| 281 | + | − | − | + | + | − | − | + | + | + | − | + | − | + |
| 288 | + | − | − | + | + | − | − | + | + | + | − | + | − | + |
| 324 | + | − | − | − | − | + | − | + | + | − | − | + | − | + |
| 352 | + | − | − | − | + | − | − | + | + | + | − | + | − | + |
| 69 | + | − | + | − | − | + | − | + | + | − | − | + | − | − |
| 111 | + | + | − | + | − | + | + | + | + | + | − | + | + | + |
| 140 | + | − | − | + | + | − | − | + | + | + | − | + | − | + |
| B15 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| B54 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| B56 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| B16 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| RS26 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| RS29 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| B67 | + | − | − | − | − | + | − | + | + | − | − | − | − | + |
| B68 | + | − | − | − | − | + | − | + | + | − | − | − | − | + |
| D20 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| KS68 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| T8 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| T16 | + | − | − | − | − | + | + | + | + | − | + | − | − | + |
| T18 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| CA11 | + | − | − | − | − | + | − | + | + | − | − | − | − | + |
| P54 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
| SW1 | + | + | − | + | − | + | − | + | + | + | − | − | + | + |
| SW3 | + | − | − | − | + | − | − | + | + | − | − | − | + | − |
| SW10 | + | − | − | + | + | − | − | + | + | + | − | − | − | − |
3.4. Antibiotic Susceptibility of aLM
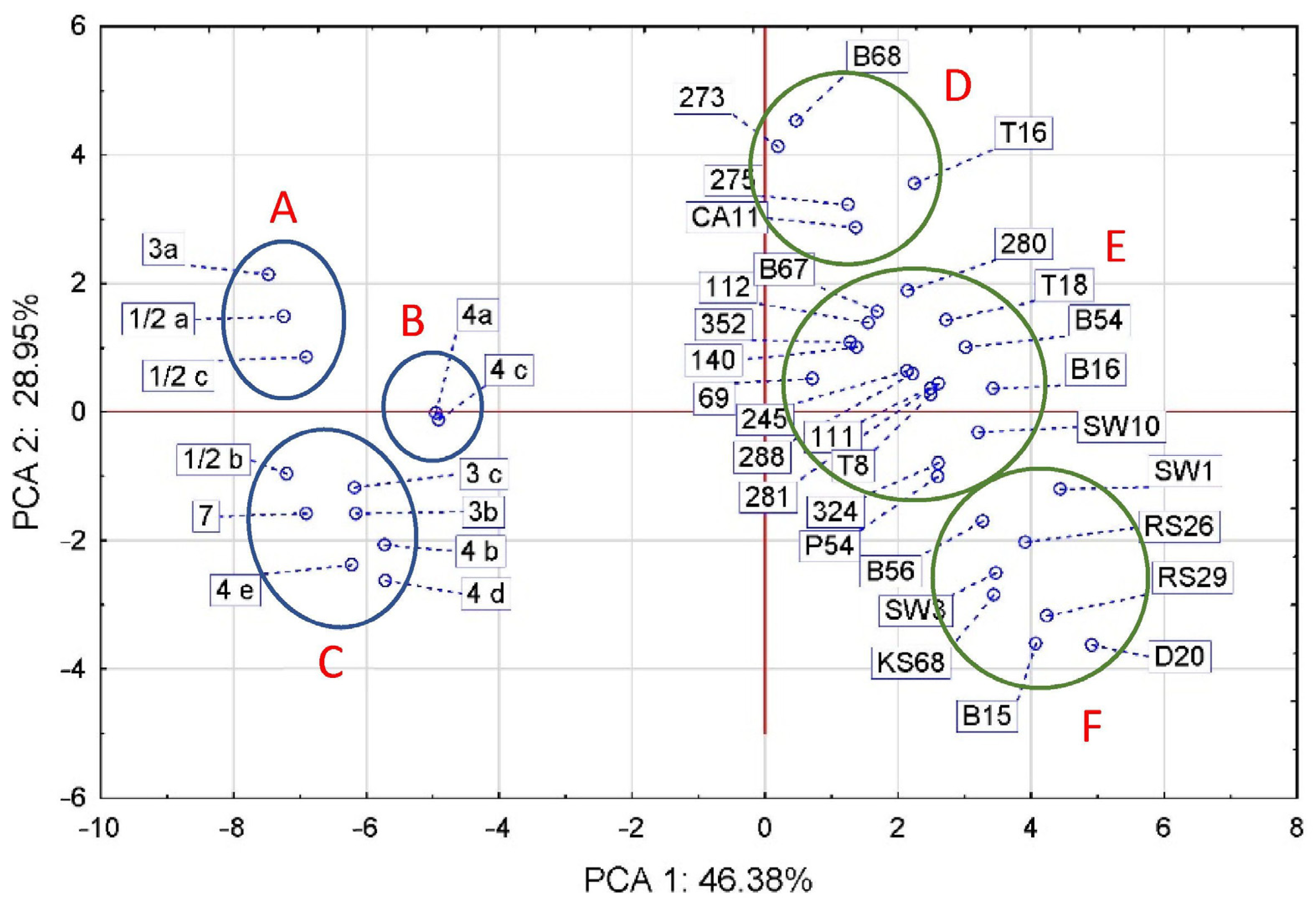
3.5. Gene Virulence and Serological Classification
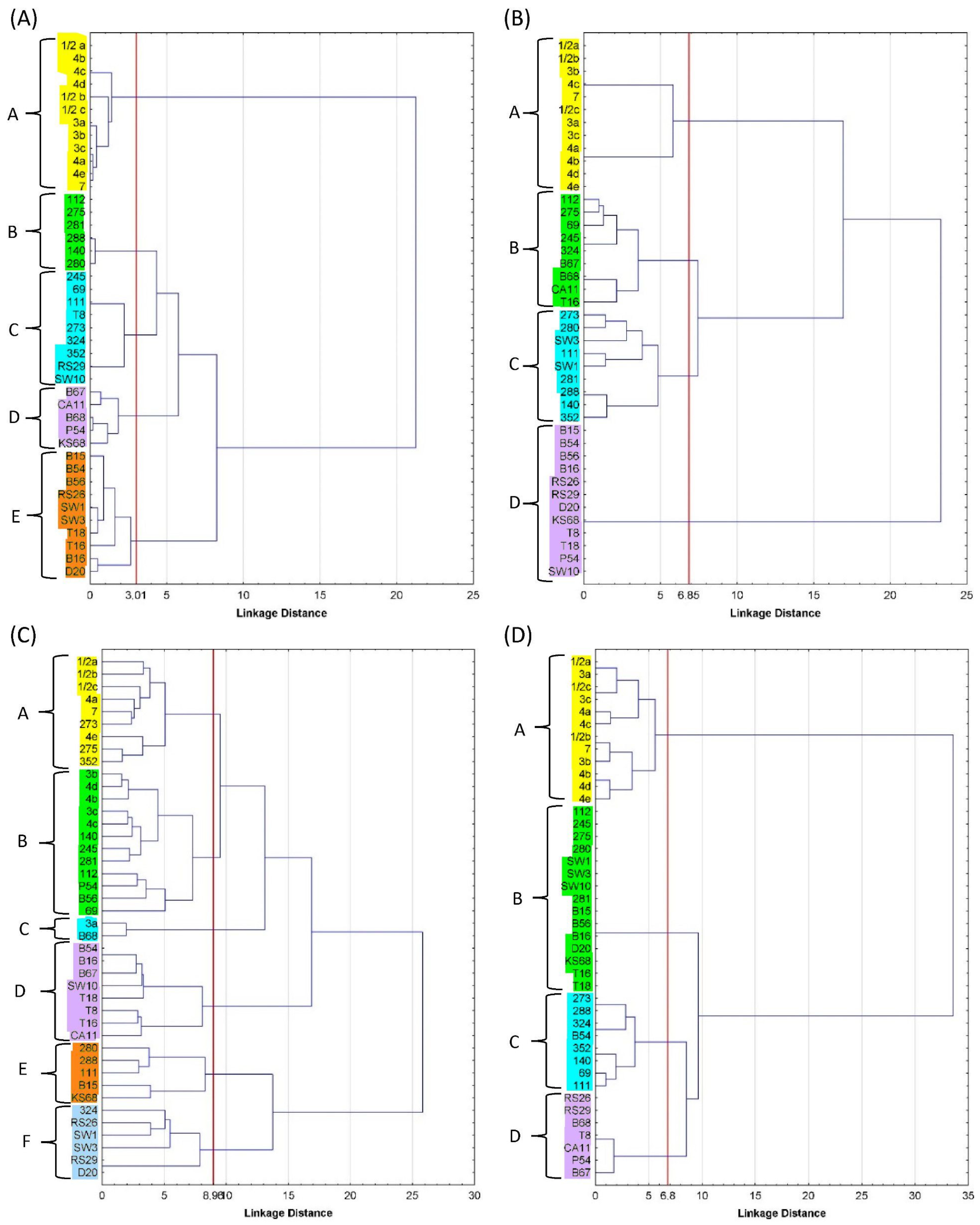
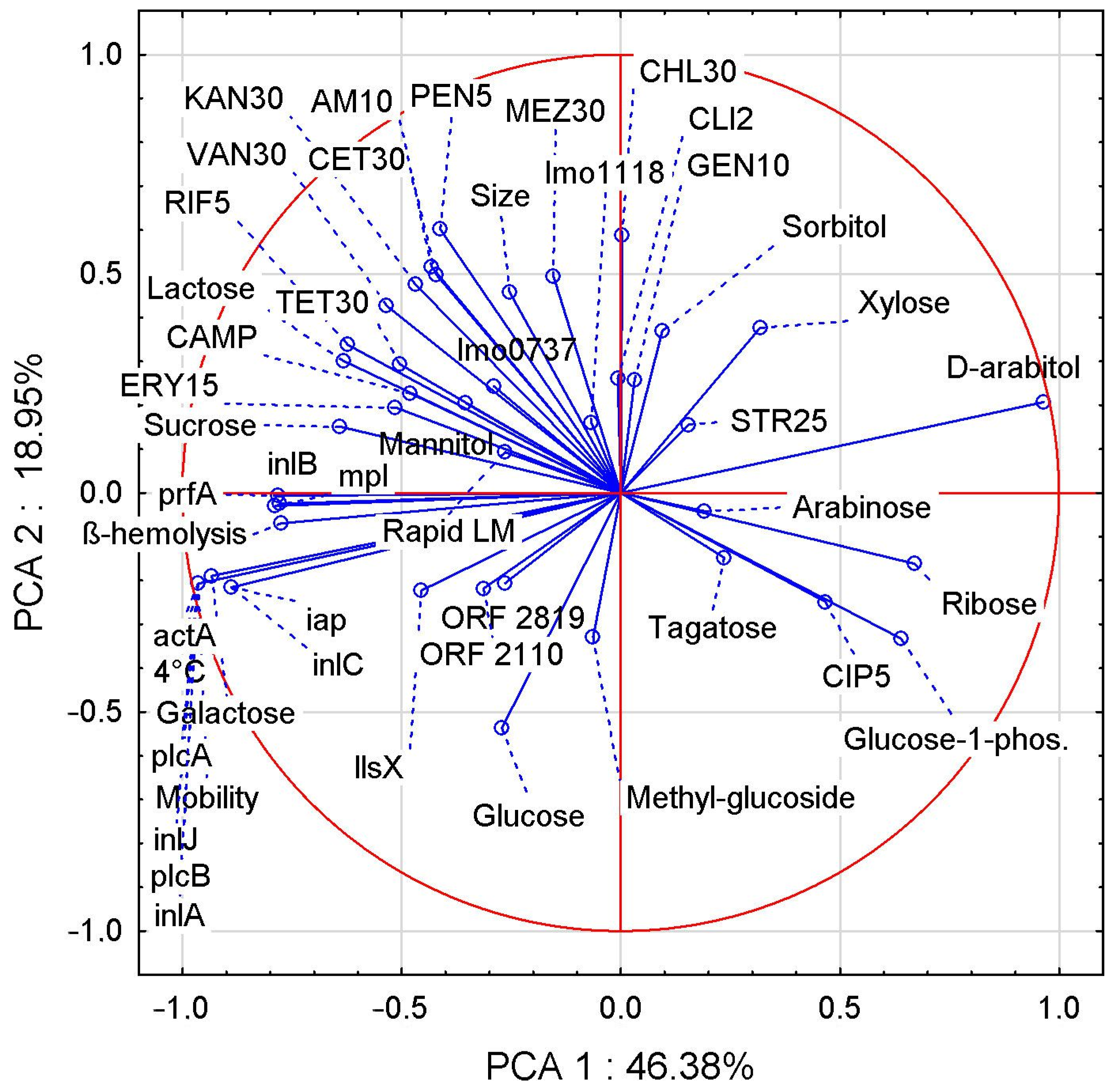
3.6. PCA Correlation Analysis
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- CDC. Listeria Outbreaks, Centers for Disease Control and Prevention. 2023. Available online: https://www.cdc.gov/listeria/outbreaks/index.html (accessed on 15 February 2023).
- EFSA. European Food Safety Authority and European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2020, 19, 6971. [Google Scholar]
- Carlin, C.R.; Liao, J.; Weller, D.; Guo, X.; Orsi, R.; Wiedmann, M. Listeria cossartiae sp. nov., Listeria immobilis sp. nov., Listeria portnoyi sp. nov. and Listeria rustica sp. nov., isolated from agricultural water and natural environments. Int. J. Syst. Evol. Microbiol. 2021, 71, 004795. [Google Scholar] [CrossRef] [PubMed]
- Den Bakker, H.C.; Warchocki, S.; Wright, E.M.; Allred, A.F.; Ahlstrom, C.; Manuel, C.S.; Stasiewicz, M.J.; Burrell, A.; Roof, S.; Strawn, L.K.; et al. Listeria floridensis sp. nov., Listeria aquatica sp. nov., Listeria cornellensis sp. nov., Listeria riparia sp. nov and Listeria grandensis sp. nov., from agricultural and natural environments. Int. J. Syst. Evol. Microbiol. 2014, 64, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomic, evolution, ecology and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Vines, A.; Swaminathan, B. Identification and characterization of nucleotide sequence differences in three virulence-associated genes of Listeria monocytogene strains representing clinically important serotypes. Curr. Microbiol. 1998, 36, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Burall, L.S.; Grim, C.J.; Datta, A.R. A clade of Listeria monocytogenes serotype 4b variant strains linked to recent listeriosis outbreaks associated with produce from a defined geographic region in the US. PLoS ONE 2017, 12, e0176912. [Google Scholar] [CrossRef]
- Laksanalamai, P.; Steyert, S.R.; Burall, L.S.; Datta, A.R. Genome sequences of Listeria monocytogenes serotype 4b variant strains isolated from clinical and environmental sources. Genome Announc. 2013, 1, 713–771. [Google Scholar] [CrossRef]
- Guidi, F.; Lorenzetti, C.; Centrorotola, G.; Torresi, M.; Camma, C.; Chiaverini, A.; Pomillio, F.; Blasi, G. Atypical serogroup IVb-v1 of Listeria monocytogenes assigned to new ST 2801 widely spread and persistent in the environment of a pork-meat producting plant of central Italy. Front. Microbiol. 2022, 13, 930895. [Google Scholar] [CrossRef]
- Baquero, F.; Lanza, V.F.; Duval, M.; Coque, T.M. Ecogenics of antibiotic resistance in Listeria monocytogenes. Mol. Microbiol. 2020, 113, 570–579. [Google Scholar] [CrossRef]
- ISO 11290-1; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection and Enumeration of Listeria monocytogenes—Part 1: Detection Method. International Organization for Standardization: Geneva, Switzerland, 2017.
- Rijpens, N.; Vlaemynck, G.; Rossau, R.; Herman, L.; Jankes, G. Unidentified Listeria-like bacteria isolated from cheese. Let. Appl. Microbiol. 1998, 27, 198–202. [Google Scholar] [CrossRef]
- Moreno, L.Z.; Paixão, R.; Sena de Gobbi, D.D.; Raimundo, D.C.; Porfida Ferreira, T.S.; Moreno, A.M.; Hofer, E.; Falavina dos Reis, C.M.; Rogério Matté, G.; Matté, M.H. Phenotypic and genotypic characterization of atypical Listeria monocytogenes and Listeria innocua isolated from swine slaughterhouses and meat markets. BioMed Res. Int. 2014, 2014, 742032. [Google Scholar] [CrossRef]
- Kawacka, I.; Olejnik-Schmidt, A.; Schmidt, M. Nonhemolitic Listeria monocytogenes–prevalence rate, reasons underlying atypical phenotype, and methods for accurate hemolysis assessment. Microorganisms 2022, 10, 483. [Google Scholar] [CrossRef]
- Szymczak, B.; Szymczak, M.; Trafiałek, J. Prevalence of Listeria species and L. monocytogenes in ready-to-eat foods in the West Pomeranian region of Poland: Correlations between the contamination level, serogroups, ingredients, and producers. Food Microbiol. 2020, 91, 103532. [Google Scholar] [CrossRef] [PubMed]
- ISO 16140-2; Microbiology of the Food Chain-Method Validation-Part 2: Protocol for the Validation of Alternative (Proprietary) Methods against a Reference Method. International Organization for Standardization: Geneva, Switzerland, 2016.
- Szymczak, B. Method of Modifying the Revitalization and DNA Isolation of Atypical Listeria monocytogenes Bacteria. Patent P.441965, 8 August 2022. (In Polish). [Google Scholar]
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J. Clin. Microbiol. 2004, 42, 3819–3822. [Google Scholar] [CrossRef] [PubMed]
- Kérouanton, A.; Marault, M.; Petit, L.; Grout, J.; Dao, T.T.; Brisabois, A. Evaluation of a multiplex PCR assay as an alternative method for Listeria monocytogenes serotyping. J. Microbiol. Methods 2010, 80, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Paziak-Domańska, B.; Bogusławska, E.; Wieckowska-Szakiel, M.; Kotłowski, R.; Różalska, B.; Chmiela, M.; Kur, J.; Dąbrowski, W.; Rudnicka, W. Evaluation of the API test, phosphatidylinositol-specific phospholipase C activity and PCR method in identification of Listeria monocytogenes in meat foods. FEMS Microbiol. Lett. 1999, 15, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Rawool, D.B.; Malik, S.V.S.; Barbuddhe, S.B.; Shakuntala, I.; Aurora, R. A multiplex PCR for detection of virulence associated genes in Listeria monocytogenes. Internet J. Food Saf. 2007, 9, 56–62. [Google Scholar] [CrossRef]
- Lomonaco, S.; Patti, R.; Knabel, S.J.; Civera, T. Detection of virulence-associated genes and epidemic clone markers in Listeria monocytogenes isolates from PDO Gorgonzola cheese. Int. J. Food Microbiol. 2012, 160, 76–79. [Google Scholar] [CrossRef]
- Jaradat, Z.W.; Schutze, G.E.; Bhunia, A.K. Genetic homogeneity among Listeria monocytogenes strains from infected patients and meat products from two geographic locations determined by phenotyping, ribotyping and PCR analysis of virulence genes. Int. J. Food Microbiol. 2002, 76, 1–10. [Google Scholar] [CrossRef]
- Tao, T.; Chen, Q.; Bie, X.; Lu, F.; Lu, Z. Mining of novel species-specific primers for PCR detection of Listeria monocytogenes based on genomic approach. World J. Microbiol. Biotech. 2015, 31, 1955–1966. [Google Scholar] [CrossRef]
- Zhou, X.; Jiao, X. Prevalence and lineages of Listeria monocytogenes in Chinese food products. Lett. Appl. Microbiol. 2006, 43, 554–559. [Google Scholar] [CrossRef]
- Liu, D.; Lawrence, M.L.; Ainsworth, A.J.; Austin, F.W. Multiplex PCR for species- and virulence-specific determination of Listeria monocytogenes. J. Microbiol. Methods 2007, 71, 133–140. [Google Scholar] [CrossRef]
- Wu, S.; Wu, Q.; Zhang, J.; Chen, M.; Yan, Z. Prevalence, antibiotic resistance and genetic diversity of Listeria monocytogenes isolated from retail ready-to-eat foods in China. Food Control 2015, 47, 340–347. [Google Scholar] [CrossRef]
- Chen, J.Q.; Healey, S.; Regan, P.; Laksanalamai, P.; Hu, Z. PCR-based methodologies for detection and characterization of Listeria monocytogenes and Listeria ivanovii in foods and environmental sources. Food Sci. Hum. Wellness 2017, 6, 39–59. [Google Scholar] [CrossRef]
- EUCAST-Disk Diffusion Method of Antimicrobial Susceptibility Testing, Version 12.0. 2022. Available online: https://www.eucast.org (accessed on 18 March 2022).
- EUCAST. European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. 2022. Version 12.0. Available online: https://www.eucast.org (accessed on 21 March 2022).
- Kwiatkowska, B. Occurence of L. monocytogenes in Soil and Plants. Ph.D. Thesis, Akademia Rolnicza, Szczecin, Poland, 2008. (In Polish). [Google Scholar]
- Colagiorgi, A.; Bruini, I.; Di Ciccio, P.A.; Zanardi, E.; Ghidini, S.; Ianieri, A. Listeria monocytogenes biofilms in the wonderland of food industry. Pathogens 2017, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Jakubowska-Gawlik, K.; Kolanowski, W.; Trafiałek, J. Evaluating suppliers of spices, casings and packaging to a meat processing plant using food safety audits data gathered during a 13-year period. Food Control 2021, 127, 108138. [Google Scholar] [CrossRef]
- Trafiałek, J.; Lehrke, M.; Lücke, F.K.; Kołożyn-Krajewska, D.; Janssen, J. HACCP-based procedures in Germany and Poland. Food Control 2015, 55, 66–74. [Google Scholar] [CrossRef]
- Trafiałek, J.; Zwoliński, M.; Kolanowski, W. Assessing hygiene practices during fish selling in retail stores. Br. Food J. 2016, 118, 2053–2067. [Google Scholar] [CrossRef]
- Denis, M.; Ziebal, C.; Boscher, E.; Picard, S.; Perrot, M.; Vila Nova, M.; Roussel, S.; Diara, A.; Pourcher, A.M. Occurrence and diversity of Listeria monocytogenes isolated from two pig manure treatment plants in France. Microb. Environ. 2022, 37, ME22019. [Google Scholar] [CrossRef]
- Szymczak, B.; Szymczak, M.; Sawicki, W.; Dąbrowski, W. Anthropogenic impact on the presence of L. monocytogenes in soil, fruits and vegetables. Folia Microbiol. 2013, 58, 4. [Google Scholar] [CrossRef]
- Smith, A.; Moorhouse, E.; Monaghan, J.; Taylor, C.; Singleton, I. Sources and survival of Listeria monocytogenes on fresh, leafy produce. J. Appl. Microbiol. 2018, 125, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Spor, A.; Camargo, A.R.O.; Bru, D.; Gaba, S.; Garmyn, D.; Gal, L.; Pivetaeu, P. Habitat disturbances modulate the barrier effect of resident soil microbiota on Listeria monocytogenes invasion success. Front. Microbiol. 2020, 11, 927. [Google Scholar] [CrossRef] [PubMed]
- Srinivasiah, S.; Bhavsar, J.; Thapar, K.; Liles, M.; Schoenfeld, T.; Wommack, K.E. Phages across the biosphere: Contrasts of viruses in soil and aquatic environments. Res. Microbiol. 2008, 159, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.D. Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar] [CrossRef]
- Leong, D.; NicAogáin, K.; Luque-Sastre, L.; McManamon, O.; Hunt, K.; Alvarez-Ordóñez, A.; Scollard, J.; Schmalenberger, A.; Fanning, S.; O’Byrne, C.; et al. A 3-year multi-food study of the presence and persistence of Listeria monocytogenes in 54 small food businesses in Ireland. Int. J. Food Microbiol. 2017, 249, 18–26. [Google Scholar] [CrossRef]
- Sevellec, Y.; Ascencio, E.; Douarre, P.E.; Felix, B.; Gal, L.; Garmyn, D.; Guiller, L.; Piveteau, P.; Roussel, S. Listeria monocytogenes: Investigation of fitness in soil does not support the relevance of ecotypes. Front. Microbiol. 2022, 13, 917588. [Google Scholar] [CrossRef]
- Bhalla, M.G.S.; Kumar, C.M.; Mahaja, M.P.; Sahai, B.K. Listerial meningitis; atypical strain and misidentification by the automated identification system (vitek2). Med. J. Armed Forces India 2019, 75, 99–101. [Google Scholar] [CrossRef]
- Palerme, J.S.; Ching Pan, P.; Parsons, C.T.; Kathariou, S.; Ward, T.J.; Jacob, M.E. Isolation and characterization of atypical Listeria monocytogenes associated with a carine urinary tract infection. J. Vet. Diagn. Investig. 2016, 28, 604–607. [Google Scholar] [CrossRef]
- Orsi, R.H.; Wiedmann, M. Characteristics and distribution of Listeria spp., including Listeria species newly described since 2009. Appl. Microbiol. Biotechnol. 2016, 100, 5273–5287. [Google Scholar] [CrossRef]
- De Vos, P.; Garrity, G.M.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.H.; Whitman, W.B. Bergey’s Manual of Systematic Bacteriology. The Firmicutes; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; Volume 3. [Google Scholar] [CrossRef]
- Maury, M.M.; Chenal-Francisque, V.; Bracq-Dieye, H.; Han, L.; Leclercq, A.; Vales, G.; Moura, A.; Gouin, E.; Scortti, M.; Disson, O.; et al. Spontaneous loss of virulence in natural populations of Listeria monocytogenes. Infect. Immun. 2017, 85, e00541-17. [Google Scholar] [CrossRef]
- Lindback, T.; Secic, I.; Rorvick, L.M. A contingency locus in prfA in a Listeria monocytogenes subgroup allows reactivation of the prfA virulence regulator during infection in mice. Appl. Environ. Microbiol. 2011, 77, 3478–3483. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tiensuu, T.; Guerreiro, D.N.; Olivieira, A.H.; O’Byrne, C.; Johansson, J. Flick of a switch: Regulatory mechanisms allowing Listeria monocytogenes to transition from a saprophyte to a killer. Microbiology 2019, 165, 819–833. [Google Scholar] [CrossRef]
- Halter, E.L.; Neuhaus, K.; Scharter, S. Listeria weihenstephanensis sp. nov., isolated from the water plant Lemna trisultica of a German fresh water pond. Int. J. Syst. Evol. Microbiol. 2013, 63, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, W.; Schleifer, K.H.; Whitman, W.B. Listeriaceae. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Vos, P., Garrity, G., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.H., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2009; pp. 244–268. [Google Scholar]
- Mafuna, T.; Matle, I.; Magwedere, K.; Pierneef, R.E.; Reva, O.N. Comparative genomics of Listeria species recovered from meat and food processing facilities. Microbiol. Spectr. 2022, 10, 5. [Google Scholar] [CrossRef]
- Burall, L.S.; Grim, C.; Gopinath, G.; Laksanalamai, P.; Datta, A.R. Whole-genome sequencing identifies an atypical Listeria monocytogenes strain isolated from pet foods. Genome Announc. 2014, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Levin, P.A.; Angert, E.R. Small but Mighty: Cell size and bacteria. Cold Spring Harb. Perspect. Biol. 2015, 7, a019216. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J.; Aké, F.; Zebre, A.; Cao, T.; Kentache, T.; Pham, Q.; Mokhtari, A.; Joyet, P.; Milohanic, E. Carbohydrate utilization by Listeria monocytogenes and its influence on virulence gene expression. In Listeria monocytogenes: Food Sources, Prevalence and Management Strategies; Hambrick, E.C., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2014; pp. 49–76. [Google Scholar]
- Gasanov, U.; Hughes, D.; Hansbro, P.M. Methods for the isolation and identification of Listeria spp. and Listeria monocytogenes: A review. FEMS Microbiol. Rev. 2005, 29, 851–875. [Google Scholar] [CrossRef]
- Jarvis, N.A.; O’Bryan, C.A.; Ricke, S.C.; Johnson, M.G.; Crandall, P.G. A review of minimal and defined media for growth of Listeria monocytogenes. Food Control 2016, 66, 256–269. [Google Scholar] [CrossRef]
- Johnson, J.; Jinneman, K.; Stelma, G.; Smith, B.G.; Lye, D.; Messer, J.; Ulaszek, J.; Evsen, L.; Gendel, S.; Bennett, R.W.; et al. Natural atypical Listeria innocua Strains with Listeria monocytogenes pathogenicity island 1 genes. Appl. Environ. Microbiol. 2004, 70, 4256–4266. [Google Scholar] [CrossRef]
- Moura, A.; Disson, O.; Lavina, M.; Thouvenot, P.; Huang, L.; Leclercq, A.; Fredriksson-Ahomaa, M.; Eshwar, A.K.; Stephan, R.; Lecuit, M. Atypical hemolytic Listeria innocua isolates are virulent, albeit less than Listeria monocytogenes. Infect. Immun. 2019, 87, e00758-18. [Google Scholar] [CrossRef]
- Gómez, D.; Azón, E.; Marco, N.; Carramiñana, J.J.; Rota, C.; Ariño, A.; Yangüela, J. Antimicrobial resistance of Listeria monocytogenes and Listeria innocua from meat products and meat-processing environment. Food Microbiol. 2014, 42, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Osaili, T.M.; Al-Nabulsi, A.A.; Taha, M.H.; Al-Holy, M.A.; Alaboudi, A.R.; Al-Rousan, W.M.; Shaker, R.R. Occurrence and antimicrobial susceptibility of Listeria monocytogenes isolated from brined white cheese in Jordan. J. Food Sci. 2012, 77, M528–M532. [Google Scholar] [CrossRef] [PubMed]
- Thedieck, K.; Hain, T.; Mohamed, W.; Tindall, B.J.; Nimtz, M.; Chakraborty, T.; Wehland, J.; Jänsch, L. The MprF protein is required for lysinylation of phospholipids in listerial membranes and confers resistance to cationic antimicrobial peptides (CAMPs) on Listeria monocytogenes. Mol. Microbiol. 2006, 62, 1325–1339. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.J.; Stasiewicz, M.J. CRISPR–Based subtyping using whole genome sequence data does not improve differentiation of persistent and sporadic Listeria monocytogenes strains. J. Food Sci. 2019, 84, 319–326. [Google Scholar] [CrossRef]
- Noll, M.; Kleta, S.; Dahouk, S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J. Infect. Public Health 2018, 11, 572–577. [Google Scholar] [CrossRef]
- Wu, S.; Wu, Q.; Zhang, J.; Chen, M.; Yan, Z.A.; Hu, H. Listeria monocytogenes prevalence and characteristics in retail raw foods in China. PLoS ONE 2015, 10, e0136682. [Google Scholar] [CrossRef]
- Soni, D.K.; Singh, M.; Singh, D.V.; Dubey, S.K. Virulence and genotypic characterization of Listeria monocytogenes isolated from vegetable and soil samples. BMC Microbiol. 2014, 14, 241. [Google Scholar] [CrossRef]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria monocytogenes clones’ adaption to mammalian gut accounts for their association with dairy products. Nat. Commun. 2019, 10, 2488. [Google Scholar] [CrossRef]
- Quereda, J.J.; Morón-García, A.; Palacios-Gorba, C.; Dessaux, C.; García-Del Portillo, F.; Pucciarelli, M.G.; Ortega, A.D. Pathogenicity and virulence of Listeria monocytogenes: A trip from environmental to medical microbiology. Virulence 2021, 12, 2509–2545. [Google Scholar] [CrossRef]
- Feng, Y.; Yao, H.; Chen, S.; Sun, X.; Yin, Y.; Jiao, X. Rapid detection of hypervirulent serovar 4 h Listeria monocytogenes by multiplex PCR. Front. Microbiol. 2020, 11, 1309. [Google Scholar] [CrossRef]



| Environmental | Sampling Locations | Number of Analyzed Samples | Count of LM Isolates | Count of aLM Isolates | aLM Isolate Number |
|---|---|---|---|---|---|
| Soil (S) | arable soil with natural fertilization (S1) | 180 | 7 | 4 | 245, 280, 281, 288 |
| arable soil artificially fertilized (S2) | 180 | ||||
| waste soil | 180 | ||||
| garden plots (S3) | 180 | 8 | 2 | 324, 112 | |
| orchards | 30 | ||||
| meadows | 35 | 4 | |||
| intensive grazing of cattle | 50 | 18 | 2 | 273, 275 | |
| usable meadows | 50 | ||||
| forest (S4) | 79 | 20 | 2 | 352, 69 | |
| area around meat processing plant | 36 | 9 | 2 | 111, 140 | |
| Fruit | strawberry from S1 | 20 | 3 | 1 | T8 |
| strawberry from S2 | 20 | ||||
| strawberry from S3 | 20 | 5 | 2 | T16, T18 | |
| raspberry from S2 | 20 | ||||
| raspberry from S3 | 20 | ||||
| blackberry from S4 | 40 | ||||
| blueberry from S3 | 20 | ||||
| Vegetables | beetroot from S1 | 10 | 5 | 4 | B15, B16, B67, B68 |
| beetroot from S2 | 10 | ||||
| beetroot from S3 | 10 | 4 | 2 | B54, B56 | |
| cabbage from S1 | 10 | ||||
| cabbage from S2 | 10 | ||||
| cabbage from S3 | 10 | ||||
| carrot from S1 | 10 | 3 | 1 | CA11 | |
| carrot from S2 | 10 | ||||
| carrot from S3 | 10 | 2 | |||
| lettuce from S1 | 10 | ||||
| lettuce from S2 | 10 | ||||
| lettuce from S3 | 10 | ||||
| parsley from S1 | 10 | 2 | |||
| parsley from S2 | 10 | ||||
| parsley from S3 | 10 | 1 | |||
| potato from S1 | 10 | 3 | |||
| potato from S2 | 10 | ||||
| potato from S3 | 10 | 2 | 1 | P54 | |
| tomato from S1 | 10 | ||||
| tomato from S2 | 10 | ||||
| tomato from S3 | 10 | ||||
| RTE food | dumplings | 200 | 41 | 1 | D20 |
| croquettes | 60 | 14 | |||
| salads | 40 | 14 | |||
| sandwiches | 40 | 2 | |||
| sushi | 40 | 8 | |||
| salted fish | 5 | ||||
| smoked fish | 5 | 1 | |||
| fish paste | 5 | 2 | |||
| cold marinated fish | 30 | 4 | |||
| desserts | 210 | ||||
| vegetable juice | 40 | 2 | |||
| Meat raw materials | pork-shoulder | 30 | 10 | ||
| neck of pork | 30 | 5 | |||
| pork ham | 30 | 8 | |||
| intestines | 30 | 12 | |||
| pork loin | 30 | 7 | |||
| Fish raw materials | Norwegian salmon | 40 | 10 | ||
| trout | 20 | ||||
| tuna | 20 | ||||
| Salads ingredients | iceberg lettuce | 20 | |||
| kale sprouts | 20 | 5 | 1 | KS68 | |
| radish sprouts | 20 | 6 | 2 | RS26, RS29 | |
| radish | 20 | 3 | |||
| tomato | 20 | ||||
| feta cheese | 20 | ||||
| Swabs in fish plants | tunnel for the pasteurizer | 5 | |||
| production line floor | 5 | 1 | |||
| water drainage system | 5 | ||||
| pasteurizer turbine | 5 | ||||
| seal in the hall door | 5 | 3 | 1 | SW10 | |
| floor drain | 5 | 2 | |||
| transport box | 5 | 1 | |||
| pasteurizer | 5 | 2 | |||
| knife | 5 | ||||
| floor in the raw material cold store | 5 | 1 | |||
| Swabs in meat plants | pasteurizer belt | 5 | 2 | 1 | SW1 |
| pasteurizer rollers | 5 | 1 | 1 | SW3 | |
| seals for cold store doors | 5 | 1 | |||
| sluice gaskets for the production hall | 5 | 2 | |||
| knife | 5 | ||||
| shelf for storing knives after sterilization | 5 | ||||
| floor drain | 5 | 1 | |||
| production line floor | 5 | 1 | |||
| transport box for raw material | 5 | 2 | |||
| product transport box | 5 | 0 | |||
| All samples | total: | 2495 | 262 | 30 |
| Target Gene | Size of Amplified Product (bp) | Primer Sequences (5′–3′) | PCR Conditions | Reference |
|---|---|---|---|---|
| prfA | 274 | F: GATACAGAAACATCGGTTGGC | 94 °C—3 min; 36 cycles: 94 °C—40 s, 60 °C—45 s, 72 °C—1,15 min; 72 °C—7 min | [19] |
| R: GTGTAATCTTGATGCCATCAGG | ||||
| hly | 456 | F: GCAGTTGCAAGCGCTTGGAGTGAA | 95 °C—2 min; 35 cycles: 95 °C—15 s, 59,5 °C—30 s, 72 °C—1,30 min; 72 °C—10 min | [20] |
| R: GCAACGTATCCTCCAGAGTGATCG | ||||
| iap | 131 | F: ACAAGCTGCACCTGTTGCAG | 95 °C—2 min; 35 cycles: 95 °C—15 s, 58 °C—30 s, 72 °C—1,30 min; 72 °C—10 min | [21] |
| R: TGACAGCGTGTGTAGTAGCA | ||||
| plcA | 326 | F: CTCGGACCATTGTAGTCATCTT | 95 °C—2 min; 35 cycles: 95 °C—15 s, 62 °C—30 s, 72 °C—1,30 min; 72 °C—10 min | [22] |
| R: CACTTTCAGGCGTATTAGAAACGA | ||||
| plcB | 261 | F: GGG AAA TTT GAC ACA GCG TT | 94 °C—3 min; 35 cycles: 94 °C—1 min, 62 °C—2 min, 72 °C—1 min; 72 °C—7 min | [23] |
| R: ATT TTC GGG TAG TCC GCT TT | ||||
| mpl | 679 | F: TGATGAAATAAAGGTCCACG | 94 °C—3 min; 35 cycles: 94 °C—30 s, 60 °C—30 s, 72 °C—40 s; 72 °C—10 min | [24] |
| R: CAAGCCATAATGAACAAACG | ||||
| actA | 827 | F: GCTGATTTAAGAGATAGAGGAACA | 95 °C—2 min; 40 cycles: 95 °C—10 s, 60 °C—30 s, 72 °C—30 s; 72 °C—10 min | [25] |
| R: TTTATGTGGTTATTTGCTGTC | ||||
| inlA | 800 | F: ACGAGTAACGGGACAAATGC | 94 °C—2 min; 30 cycles: 94 °C—20 s, 60 °C—20 s, 72 °C—50 s; 72 °C—7 min | [26] |
| R: CCCGACAGTGGTGCTAGATT | ||||
| inlB | 884 | F: TGGGAGAGTAACCCAACCAC | 94 °C—2 min; 30 cycles: 94 °C—20 s, 65 °C—20 s, 72 °C—50 s; 72 °C—7 min | |
| R: GTTGACCTTCGATGGTTGCT | ||||
| inlC | 517 | F: AATTCCCACAGGACACAACC | 94 °C—2 min; 30 cycles: 94 °C—20 s, 60 °C—20 s, 72 °C—50 s; 72 °C—7 min | |
| R: CGGGAATGCAATTTTTCACTA | ||||
| inlJ | 238 | F: TGTAACCCCGCTTACACAGTT | 94 °C—2 min; 30 cycles: 94 °C—20 s, 60 °C—20 s, 72 °C—50 s; 72 °C—7 min | |
| R: AGCGGCTTGGCAGTCTAATA | ||||
| llsX | 200 | F: TTATTGCATCAATTGTTCTAGGG | 95 °C—3 min; 45 cycles: 95 °C—30 s, 60 °C—1 min, 72 °C—1 min; 72 °C—10 min | [27] |
| R: CCCCTATAAACATCATGCTAGTG | ||||
| prs | 370 | F: GCTGAAGAGATTGCGAAAGAAG | 94 °C—3 min, 35 cycle: 94 °C—40s, 53 °C—1.15 min, 72 °C—1.15 min;72 °C—7 min | [18] |
| R: CAAAGAAACCTTGGATTTGCGG | ||||
| ORF2819 | 471 | F: AGCAAAATGCCAAAACTCGT | ||
| R: CATCACTAAAGCCTCCCATTG | ||||
| ORF2110 | 597 | F: AGTGGACAATTGATTGGTGAA | ||
| R: CATCCATCCCTTACTTTGGAC | ||||
| lmo0737 | 691 | F: AGGGCTTCAAGGACTTACCC | ||
| R: ACGATTTCTGCTTGCCATTC | ||||
| lmo1118 | 906 | F: AGGGGTCTTAAATCCTGGAA | ||
| R: CGGCTTGTTCGGCATACTTA | ||||
| LMOf2365_0970 | 386 | F: GCTCAGCGGCAAATCAAAC | 94 °C—3 min; 35 cycles: 94 °C—30 s, 60 °C—30 s, 72 °C—40 s; 72 °C—10 min | [24] |
| R: GGCACTCGCAACAGAAACG | ||||
| LMOf2365_2721 | 583 | F: GTTCGTCGGTCCGTGGTA | 94 °C—3 min; 35 cycles: 94 °C—30 s, 60 °C—30 s, 72 °C—40 s; 72 °C—10 min | [28] |
| R: TTGGCAAGCAAGCAGTTCA |
| Sample | Cell Size | Mobility | Growth at (°C) | β—Hemolysis | CAMP Test | Chromogenic Medium | Esculin, Voges–Proskauer, Methyl Red | Nitrate Reduction | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 | 30 and 37 | S. aureus | R. equi | OCLA and ALOA | Rapid L. mono | ||||||
| 1/2a | 3.0 | + | + | + | + | + | − | BH | GB | + | − |
| 1/2b | 2.0 | + | + | + | + | + | − | BH | GB | + | − |
| 1/2c | 2.8 | + | + | + | + | + | − | BH | GB | + | − |
| 3a | 2.8 | + | + | + | + | + | − | BH | GB | + | − |
| 3b | 2.8 | + | + | + | + | + | − | BH | GB | + | − |
| 3c | 2.7 | + | + | + | + | + | − | BH | GB | + | − |
| 4a | 2.7 | + | + | + | + | + | − | BH | GB | + | − |
| 4b | 3.0 | + | + | + | + | + | − | BH | GB | + | − |
| 4c | 3.0 | + | + | + | + | + | − | BH | GB | + | − |
| 4d | 3.0 | + | + | + | + | + | − | BH | GB | + | − |
| 4e | 2.6 | + | + | + | + | + | − | BH | GB | + | − |
| 7 | 2.6 | + | + | + | + | + | − | BH | GB | + | − |
| 112 | 3.0 | − | − | + | − | + | − | BH | GB | + | − |
| 245 | 2.0 | − | − | + | − | + | − | BH | GB | + | − |
| 273 | 2.5 | − | − | + | − | + | − | BH | GB | + | − |
| 275 | 3.0 | − | − | + | − | + | − | BH | GB | + | − |
| 280 | 3.2 | − | − | + | − | + | − | BH | GB | + | − |
| 281 | 3.0 | − | − | + | − | + | − | BH | GB | + | − |
| 288 | 3.0 | − | − | + | − | + | − | BH | GB | + | − |
| 324 | 2.5 | − | − | + | − | + | − | BH | GB | + | − |
| 352 | 2.5 | − | − | + | − | + | − | BH | GB | + | − |
| 69 | 2.0 | − | − | + | − | + | − | BH | GB | + | − |
| 111 | 2.0 | − | − | + | − | + | − | BH | GB | + | − |
| 140 | 3.0 | − | − | + | − | + | − | BH | GB | + | − |
| B15 | 2.0 | − | − | + | − | − | − | BH | GB | + | − |
| B54 | 2.5 | − | − | + | − | − | − | BH | GB | + | − |
| B56 | 2.5 | − | − | + | − | − | − | BH | GB | + | − |
| B16 | 2.5 | − | − | + | − | − | − | BH | W | + | − |
| RS26 | 2.5 | − | − | + | − | − | − | BH | GB | + | − |
| RS29 | 2.5 | − | − | + | − | + | − | BH | GB | + | − |
| B67 | 2.5 | − | − | + | + | + | − | BH | GB | + | − |
| B68 | 3.0 | − | − | + | + | + | − | BH | W | + | − |
| D20 | 2.0 | − | − | + | − | − | − | BH | W | + | − |
| KS68 | 2.0 | − | − | + | + | + | − | BH | W | + | − |
| T8 | 2.0 | − | − | + | − | + | − | BH | GB | + | − |
| T16 | 3.5 | − | − | + | − | − | − | BH | GB | + | − |
| T18 | 2.8 | − | − | + | − | − | − | BH | GB | + | − |
| CA11 | 3.2 | − | − | + | + | + | − | BH | GB | + | − |
| P54 | 2.8 | − | − | + | + | + | − | BH | W | + | − |
| SW1 | 2.5 | − | − | + | − | − | − | BH | GB | + | − |
| SW3 | 2.5 | − | − | + | − | − | − | BH | GB | + | − |
| SW10 | 2.5 | − | − | + | − | + | − | BH | GB | + | − |
| Sample | Antibiotics | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GEN 10 | STR 25 | KAN 30 | CHL 30 | RIF 5 | CET 30 | VAN 30 | CLI 2 | ERY 15 | AMP 10 | MEZ 75 | PEN 10 | CIP 5 | TET 30 | |
| 1/2a | 25 | 23 | 30 | 29 | 32 | 28 | 30 | 13 | 33 | 28 | 31 | 40 | 25 | 30 |
| 1/2b | 20 | 21 | 24 | 28 | 30 | 29 | 26 | 15 | 36 | 33 | 32 | 28 | 23 | 30 |
| 1/2c | 27 | 23 | 26 | 27 | 34 | 28 | 26 | 21 | 32 | 39 | 27 | 31 | 20 | 27 |
| 3a | 29 | 30 | 32 | 26 | 25 | 32 | 31 | 13 | 38 | 41 | 37 | 30 | 18 | 37 |
| 3b | 26 | 20 | 26 | 26 | 28 | 24 | 26 | 18 | 28 | 24 | 24 | 21 | 22 | 28 |
| 3c | 23 | 18 | 28 | 21 | 30 | 26 | 23 | 12 | 26 | 27 | 28 | 25 | 19 | 26 |
| 4a | 28 | 22 | 28 | 24 | 28 | 26 | 26 | 12 | 32 | 31 | 30 | 28 | 21 | 30 |
| 4b | 28 | 19 | 25 | 24 | 28 | 26 | 24 | 10 | 26 | 26 | 25 | 25 | 22 | 28 |
| 4c | 22 | 20 | 28 | 23 | 26 | 24 | 25 | 14 | 29 | 30 | 31 | 30 | 19 | 26 |
| 4d | 24 | 20 | 24 | 21 | 28 | 23 | 26 | 14 | 28 | 24 | 24 | 22 | 21 | 25 |
| 4e | 23 | 25 | 20 | 24 | 28 | 20 | 28 | 15 | 29 | 40 | 28 | 24 | 25 | 26 |
| 7 | 25 | 15 | 27 | 25 | 26 | 26 | 28 | 19 | 31 | 27 | 25 | 25 | 20 | 24 |
| 112 | 20 | 17 | 21 | 28 | 21 | 21 | 26 | 12 | 30 | 21 | 29 | 29 | 20 | 27 |
| 245 | 23 | 24 | 23 | 23 | 22 | 23 | 21 | 14 | 30 | 27 | 23 | 27 | 22 | 24 |
| 273 | 24 | 20 | 24 | 29 | 27 | 26 | 28 | 22 | 31 | 32 | 31 | 29 | 22 | 28 |
| 275 | 23 | 15 | 24 | 26 | 27 | 27 | 27 | 12 | 26 | 33 | 31 | 27 | 23 | 27 |
| 280 | 33 | 28 | 26 | 31 | 31 | 20 | 24 | 0 | 30 | 25 | 31 | 23 | 30 | 12 |
| 281 | 23 | 17 | 21 | 23 | 25 | 23 | 23 | 10 | 29 | 31 | 28 | 29 | 22 | 22 |
| 288 | 31 | 22 | 32 | 29 | 24 | 16 | 23 | 10 | 22 | 19 | 30 | 22 | 23 | 12 |
| 324 | 23 | 21 | 23 | 18 | 20 | 14 | 22 | 10 | 26 | 19 | 17 | 18 | 27 | 28 |
| 352 | 22 | 20 | 25 | 26 | 26 | 24 | 28 | 13 | 26 | 30 | 28 | 27 | 22 | 22 |
| 69 | 21 | 0 | 23 | 24 | 25 | 25 | 23 | 15 | 28 | 30 | 26 | 27 | 17 | 10 |
| 111 | 29 | 24 | 32 | 25 | 27 | 18 | 26 | 0 | 27 | 20 | 30 | 24 | 27 | 15 |
| 140 | 22 | 22 | 25 | 26 | 26 | 25 | 22 | 14 | 32 | 28 | 28 | 27 | 17 | 29 |
| B15 | 22 | 13 | 24 | 23 | 22 | 18 | 18 | 0 | 28 | 10 | 27 | 10 | 23 | 25 |
| B54 | 30 | 27 | 20 | 31 | 22 | 26 | 25 | 25 | 28 | 31 | 31 | 27 | 28 | 20 |
| B56 | 15 | 14 | 18 | 30 | 22 | 17 | 26 | 13 | 32 | 24 | 25 | 17 | 23 | 27 |
| B16 | 30 | 27 | 20 | 31 | 22 | 26 | 25 | 25 | 28 | 31 | 31 | 27 | 28 | 20 |
| RS26 | 24 | 27 | 13 | 25 | 23 | 22 | 21 | 20 | 25 | 29 | 27 | 23 | 26 | 0 |
| RS29 | 26 | 28 | 10 | 18 | 13 | 23 | 24 | 20 | 25 | 19 | 27 | 15 | 29 | 0 |
| B67 | 28 | 32 | 16 | 30 | 24 | 25 | 27 | 25 | 29 | 32 | 29 | 28 | 30 | 10 |
| B68 | 28 | 30 | 30 | 26 | 24 | 32 | 28 | 18 | 34 | 39 | 38 | 32 | 18 | 34 |
| D20 | 26 | 28 | 10 | 18 | 13 | 23 | 24 | 20 | 25 | 19 | 27 | 15 | 29 | 0 |
| KS68 | 26 | 20 | 28 | 24 | 24 | 20 | 24 | 0 | 28 | 11 | 29 | 0 | 30 | 30 |
| T8 | 27 | 27 | 21 | 31 | 32 | 29 | 26 | 22 | 24 | 25 | 26 | 23 | 21 | 25 |
| T16 | 27 | 27 | 31 | 30 | 25 | 28 | 23 | 23 | 21 | 27 | 25 | 22 | 20 | 22 |
| T18 | 27 | 26 | 22 | 32 | 28 | 24 | 25 | 21 | 28 | 36 | 34 | 33 | 29 | 27 |
| CA11 | 26 | 25 | 31 | 29 | 27 | 29 | 27 | 21 | 24 | 28 | 32 | 23 | 25 | 14 |
| P54 | 22 | 15 | 20 | 26 | 21 | 23 | 23 | 20 | 29 | 27 | 30 | 17 | 19 | 24 |
| SW1 | 25 | 29 | 19 | 25 | 18 | 23 | 23 | 20 | 23 | 21 | 17 | 23 | 25 | 19 |
| SW3 | 24 | 28 | 18 | 12 | 24 | 20 | 22 | 0 | 26 | 29 | 27 | 26 | 27 | 0 |
| SW10 | 27 | 28 | 16 | 27 | 23 | 25 | 22 | 23 | 27 | 30 | 27 | 26 | 29 | 28 |
| Sample | Virulence Genes | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| hly | iap | prfA | inlA | inlB | inlC | inlJ | plcA | plcB | llsX | actA | mpl | LMOf 2365_ 0970 | LMOf 2365_ 2721 | Serogroup | |
| 1/2a | + | + | + | + | + | + | + | + | + | − | + | + | + | + | nc |
| 1/2b | + | + | + | + | + | + | + | + | + | + | + | + | + | + | nc |
| 1/2c | + | + | + | + | + | + | + | + | + | − | + | + | + | + | nc |
| 3a | + | + | + | + | + | + | + | + | + | − | + | + | + | + | nc |
| 3b | + | + | + | + | + | + | + | + | + | − | + | + | + | + | nc |
| 3c | + | + | + | + | + | + | + | + | + | − | + | + | + | + | nc |
| 4a | + | − | + | + | + | − | + | + | + | − | + | − | + | + | nc |
| 4b | + | + | + | + | − | + | + | + | + | − | + | + | + | + | nc |
| 4c | + | − | + | + | − | − | + | + | + | − | + | + | + | + | nc |
| 4d | + | + | + | + | − | + | + | + | + | − | + | + | + | + | nc |
| 4e | + | + | + | + | − | + | + | + | + | + | + | + | + | + | nc |
| 7 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | nc |
| 112 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| 245 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| 273 | + | − | + | − | − | − | − | − | − | − | − | + | + | + | 1/2c−3c |
| 275 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| 280 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| 281 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| 288 | + | − | + | − | − | − | − | − | − | − | − | + | + | + | 1/2c−3c |
| 324 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| 352 | + | − | + | − | − | − | − | − | − | − | − | + | + | + | 4a−4c |
| 69 | + | − | + | − | − | − | − | − | − | − | − | − | − | + | nc |
| 111 | + | − | + | − | − | − | − | − | − | − | − | − | − | + | 4a−4c |
| 140 | + | − | + | − | − | − | − | − | − | − | − | + | + | + | 4a−4c |
| B15 | + | − | − | − | − | − | − | − | − | − | − | − | + | + | nc |
| B54 | + | − | − | − | − | − | − | − | − | − | − | − | + | + | nc |
| B56 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| B16 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| RS26 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| RS29 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| B67 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| B68 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| D20 | + | − | − | − | − | − | − | − | − | − | − | − | + | nc | |
| KS68 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| T8 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| T16 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| T18 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| CA11 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| P54 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| SW1 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| SW3 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
| SW10 | + | − | − | − | − | − | − | − | − | − | − | − | − | + | nc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szymczak, B. Phenotypic and Genotypic Characteristics of Non-Hemolytic L. monocytogenes Isolated from Food and Processing Environments. Foods 2023, 12, 3630. https://doi.org/10.3390/foods12193630
Szymczak B. Phenotypic and Genotypic Characteristics of Non-Hemolytic L. monocytogenes Isolated from Food and Processing Environments. Foods. 2023; 12(19):3630. https://doi.org/10.3390/foods12193630
Chicago/Turabian StyleSzymczak, Barbara. 2023. "Phenotypic and Genotypic Characteristics of Non-Hemolytic L. monocytogenes Isolated from Food and Processing Environments" Foods 12, no. 19: 3630. https://doi.org/10.3390/foods12193630
APA StyleSzymczak, B. (2023). Phenotypic and Genotypic Characteristics of Non-Hemolytic L. monocytogenes Isolated from Food and Processing Environments. Foods, 12(19), 3630. https://doi.org/10.3390/foods12193630