



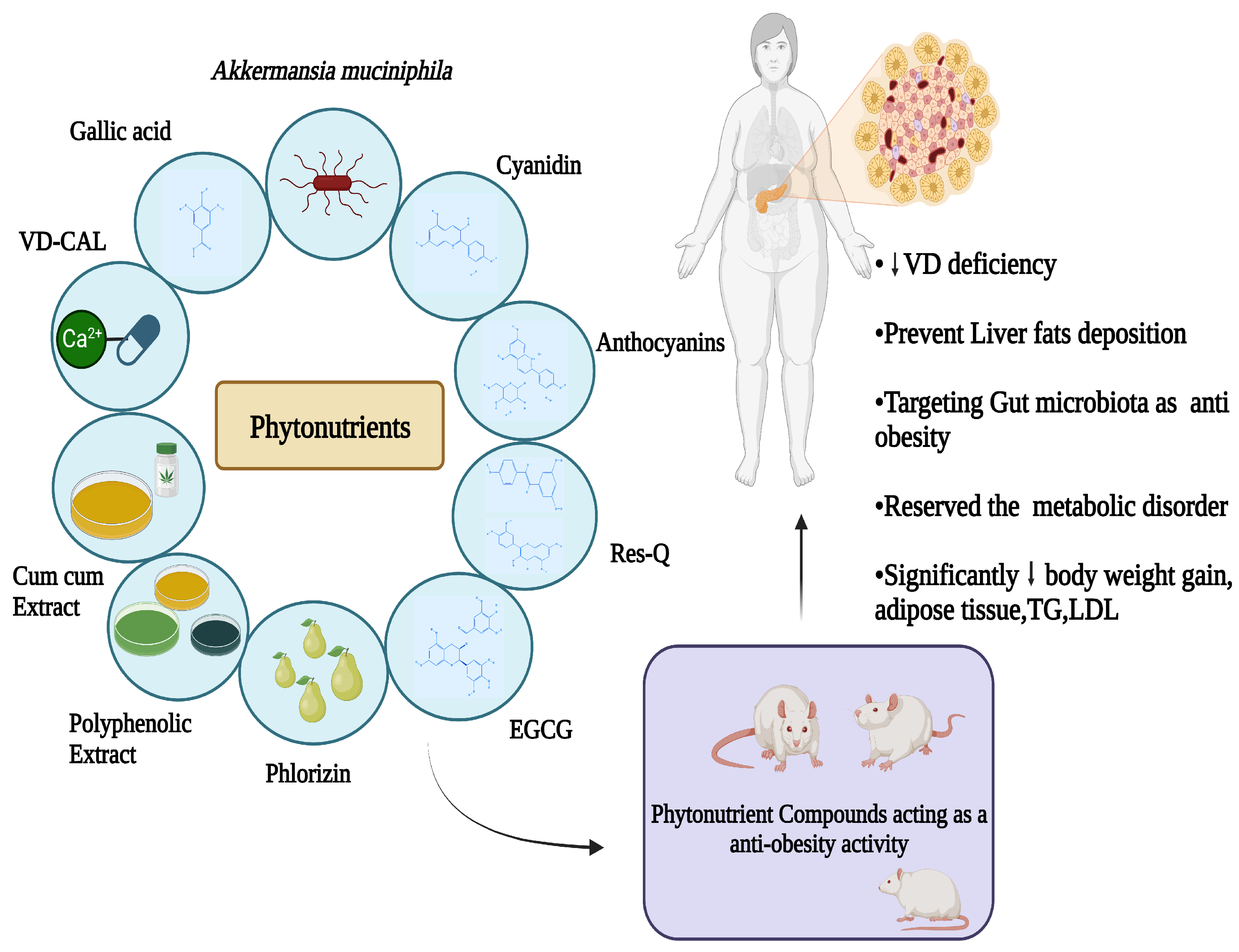
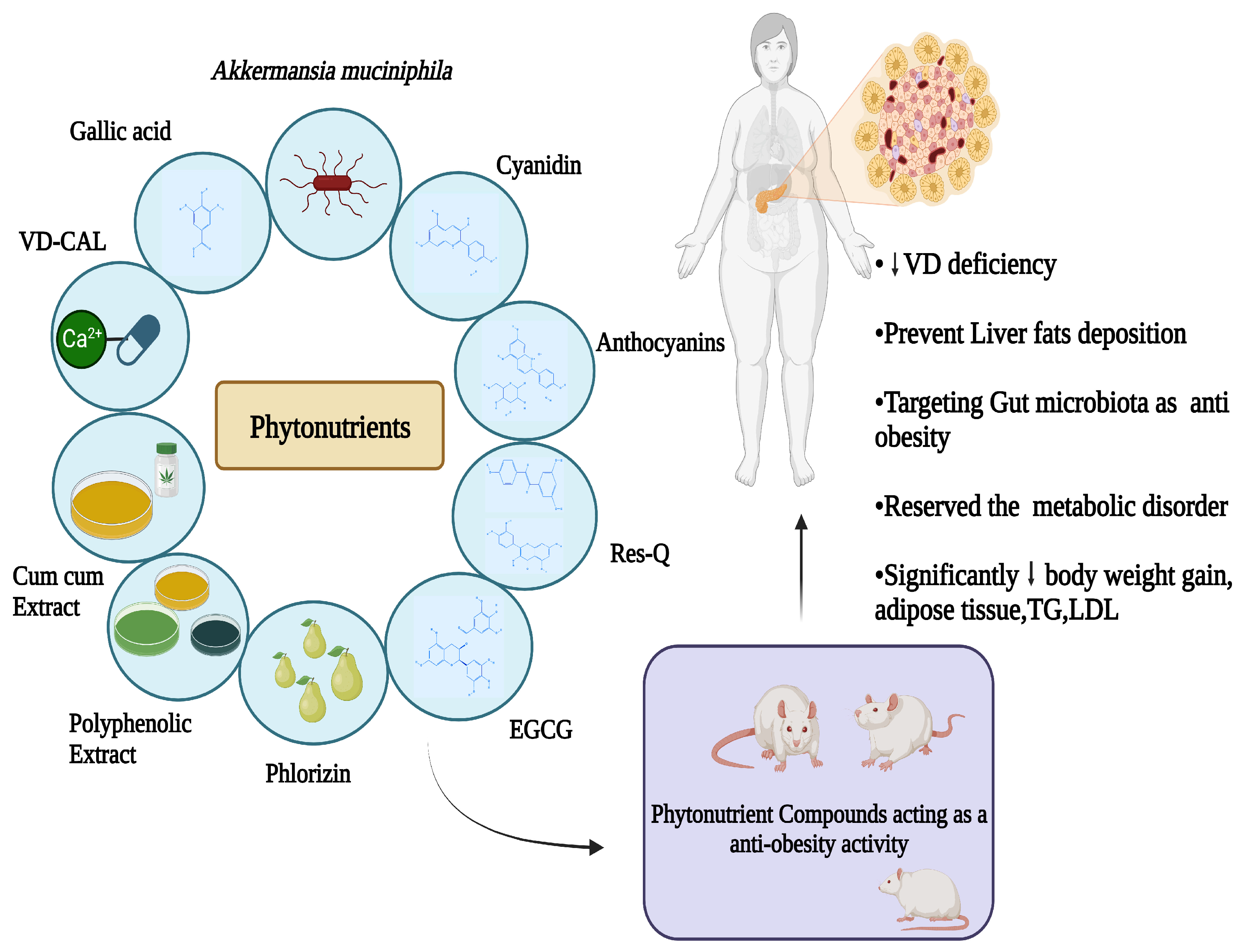
A Comprehensive Review of Phytonutrients as a Dietary Therapy for Obesity

 , , ,
, , , 
Abstract
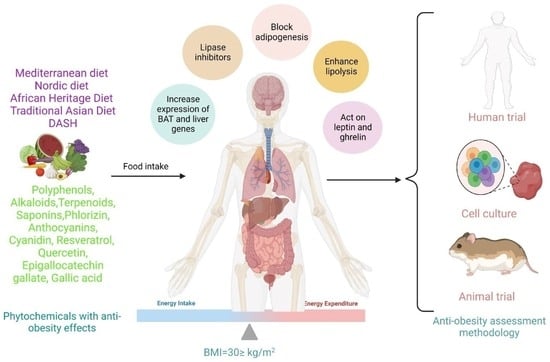
:
1. Introduction
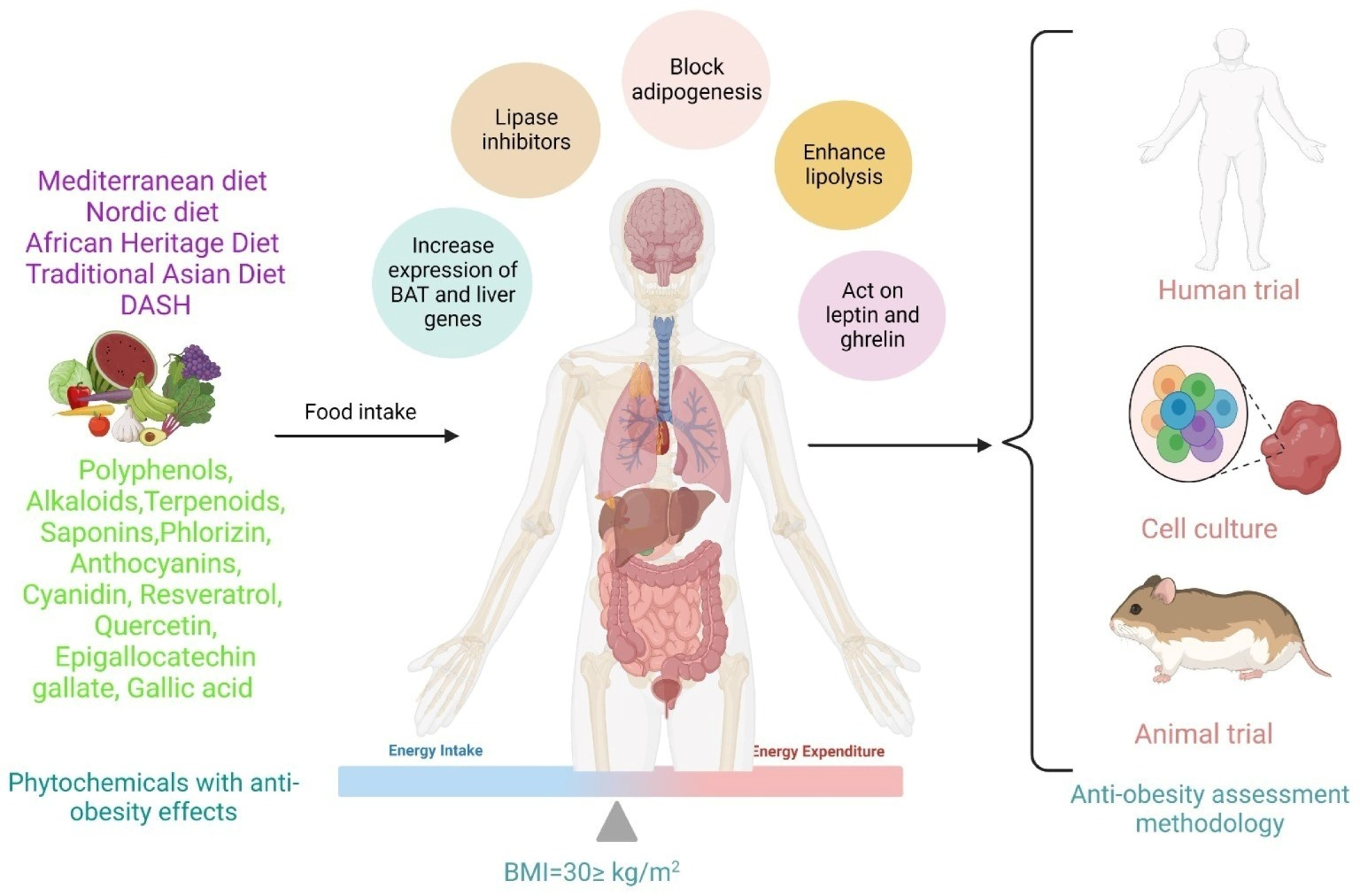
2. Phytonutrients in Regional Diet
2.1. Mediterranean Diet
2.2. Nordic Diet
2.3. African Heritage Diet
2.4. Traditional Asian Diets
2.5. Dietary Approaches to Stop Hypertension (DASH)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
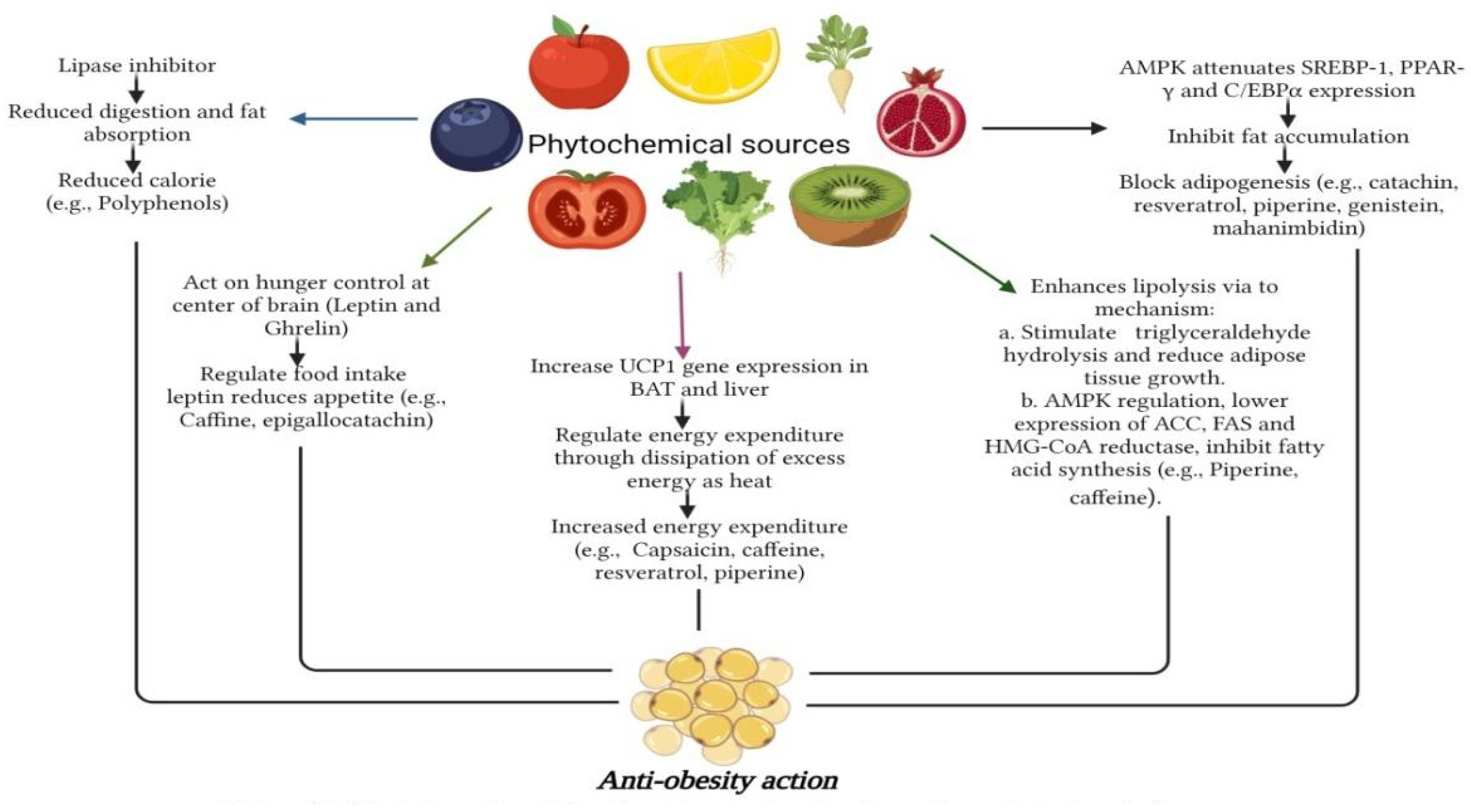{kind=link}
{kind=link}
| Diets | Phytonutrients | Regions | Outcomes | References |
|---|---|---|---|---|
| Mediterranean diet | Fruit, vegetables, breads, other forms of cereals, potatoes, beans, nuts, and seeds | Africa, Asia, and Europe | Monitoring diet, predominantly low in saturated fat; reduced amount 7–8% | [28,29,30] |
| Nordic diet | Apples, pears, berries, root and cruciferous vegetables, cabbages, whole grains, rye bread, intake of fish, low-fat dairy products, potatoes, and vegetable fats | Northern Europe and Europe | Stimulate low glycemic action, the contented of high glycemic index, lower effect, and blood sugar level | [33,35,36] |
| African Heritage diet | A plant-heavy diet with fruits, vegetables, tubers, grains, beans, nuts, healthy oils, and seafood | African, African-American, Caribbean, and South American | Reduced body weight and waist edgesand reduced blood pressure | [41,42,43] |
| Traditional Asian diets | Rice, whole grains, fermented food, indigenous land and sea vegetables, proteins(legumes and fish meat), medicinal herbs (garlic, green onions, andginger), sesame, and perilla oils | Asia | Lower risk of metabolic syndrome, obesity, hypertriglyceridemia, and hypertension | [44,45,48] |
| Dietary Approaches to Stop Hypertension (DASH) | Rich in fruits, vegetables, and low-fat dairy | East Asia | Developments to lower overweight/obesity, low-density lipoprotein cholesterol (LDL-C), and total cholesterol | [52,53] |
3. Bioavailability of Phytonutrients in the Human Metabolism System
3.1. Digestion and Absorption
3.2. Interaction with Other Substances
3.3. Individual Differences
3.4. Cooking and Food Processing
3.5. Bioavailability of Phytonutrients as Anti-Obesity Activities
4. Phytonutrients with Anti-Obesity Effects
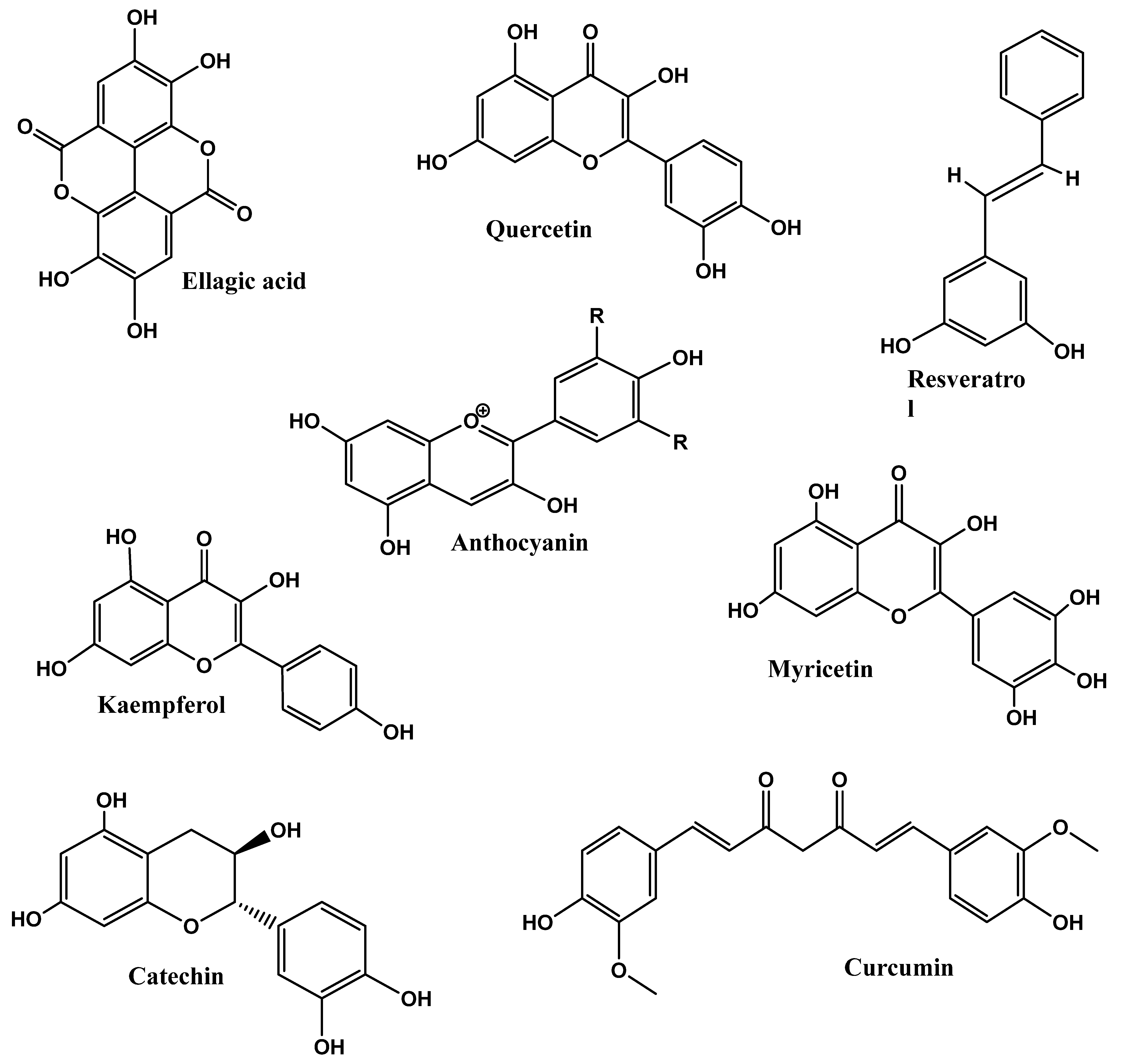
4.1. Polyphenols
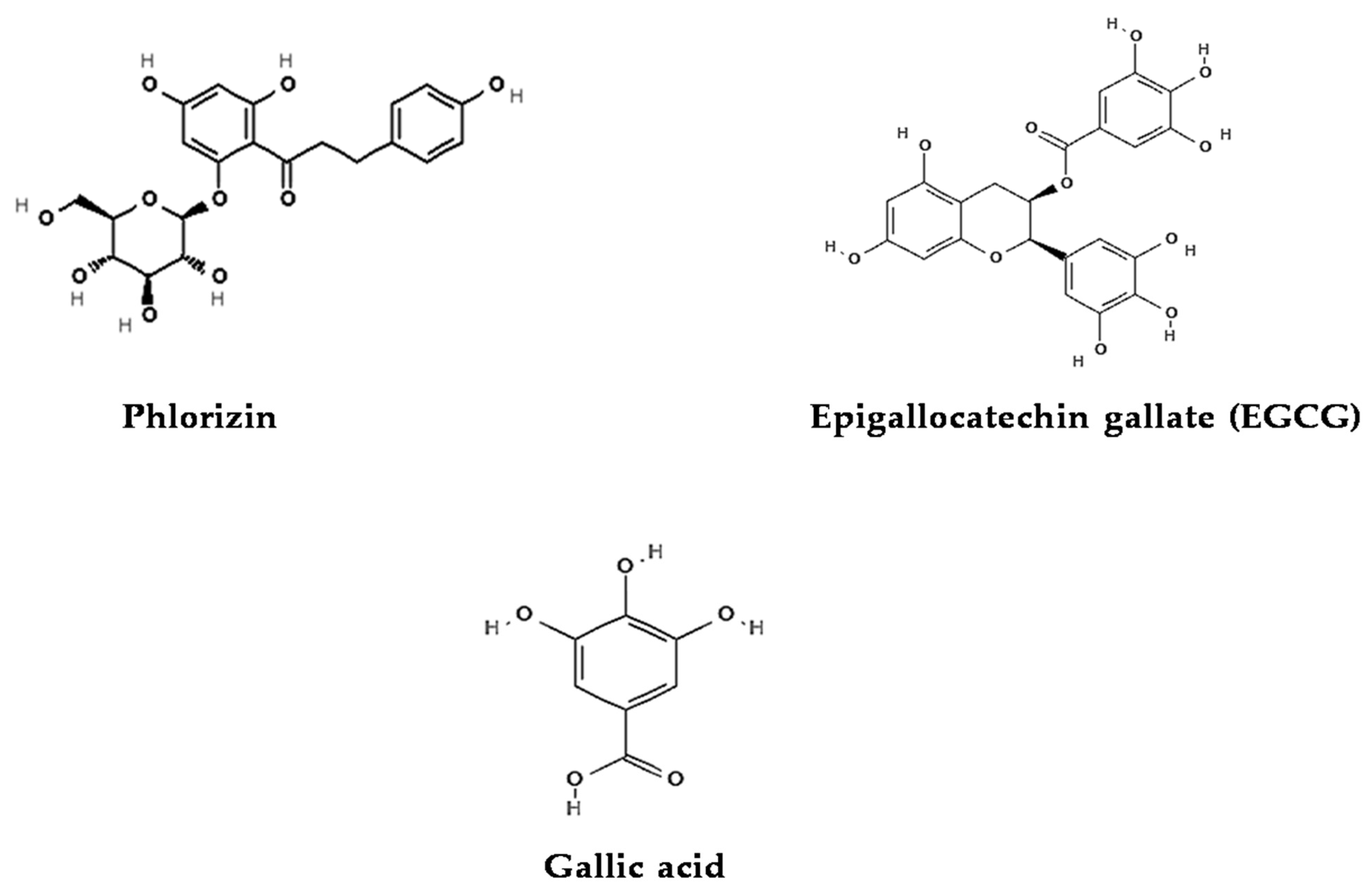
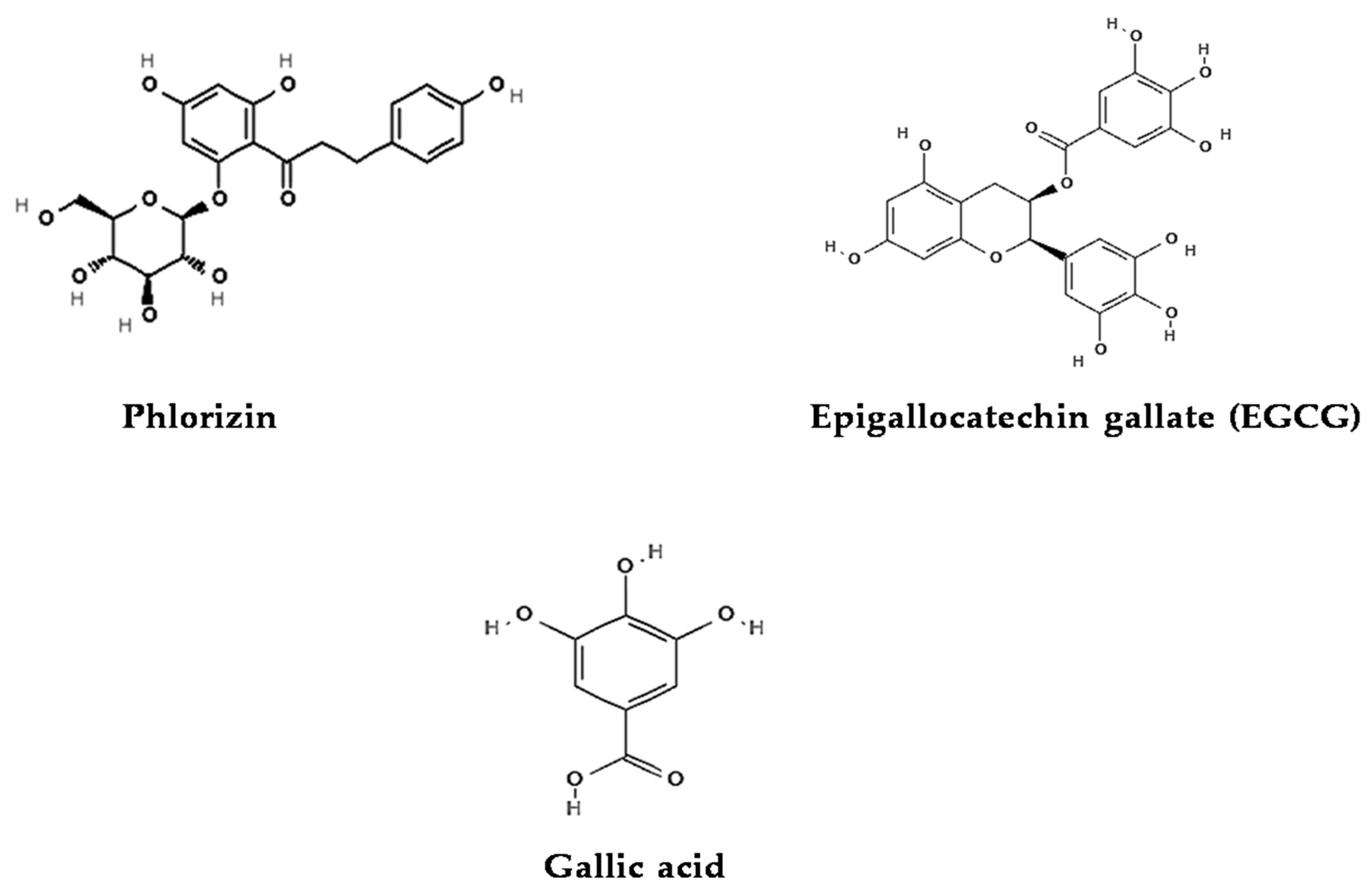
4.1.1. Phlorizin
4.1.2. Epigallocatechin Gallate (EGCG)
4.1.3. Gallic Acid
4.1.4. Resveratrol and Quercetin
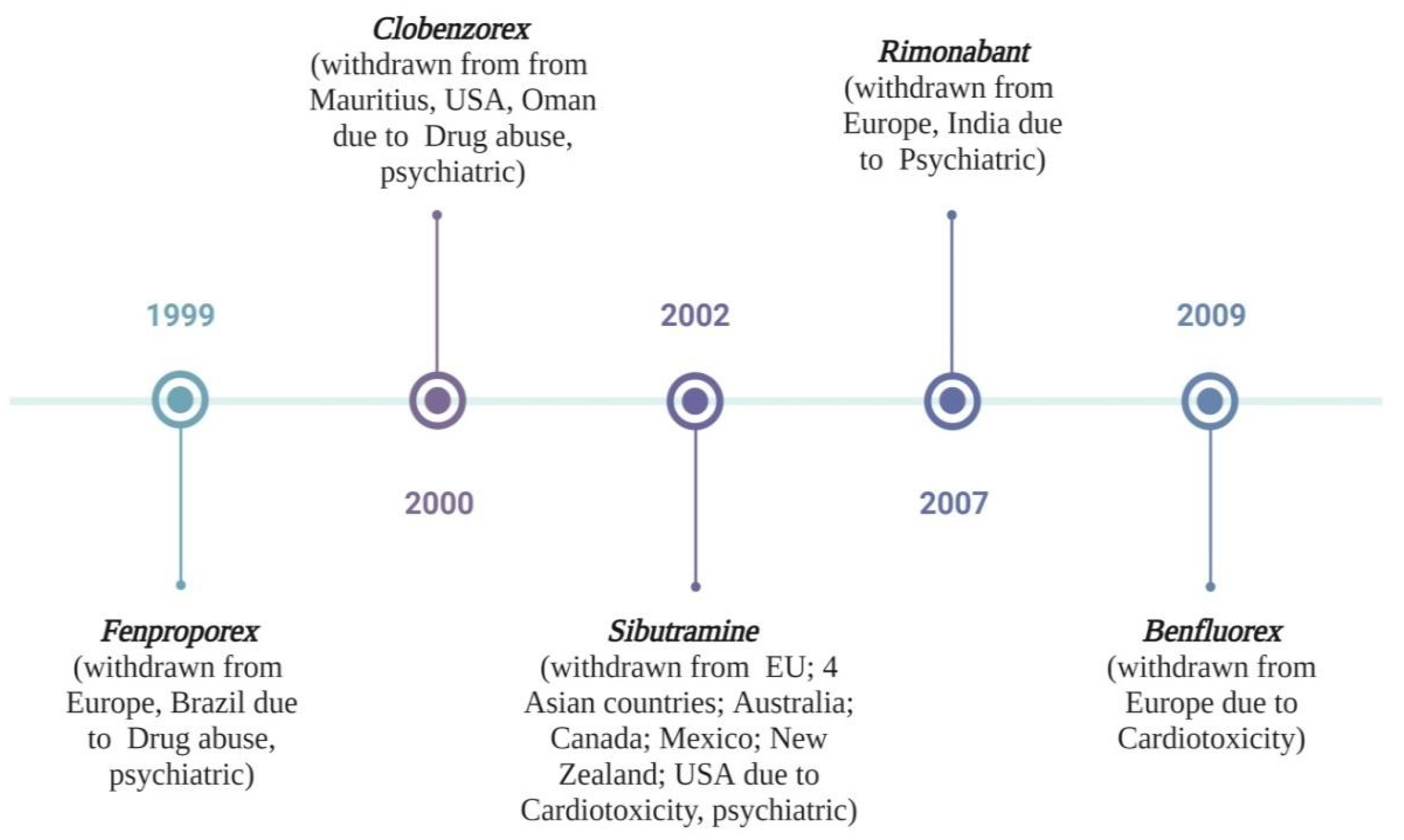
4.2. Alkaloids
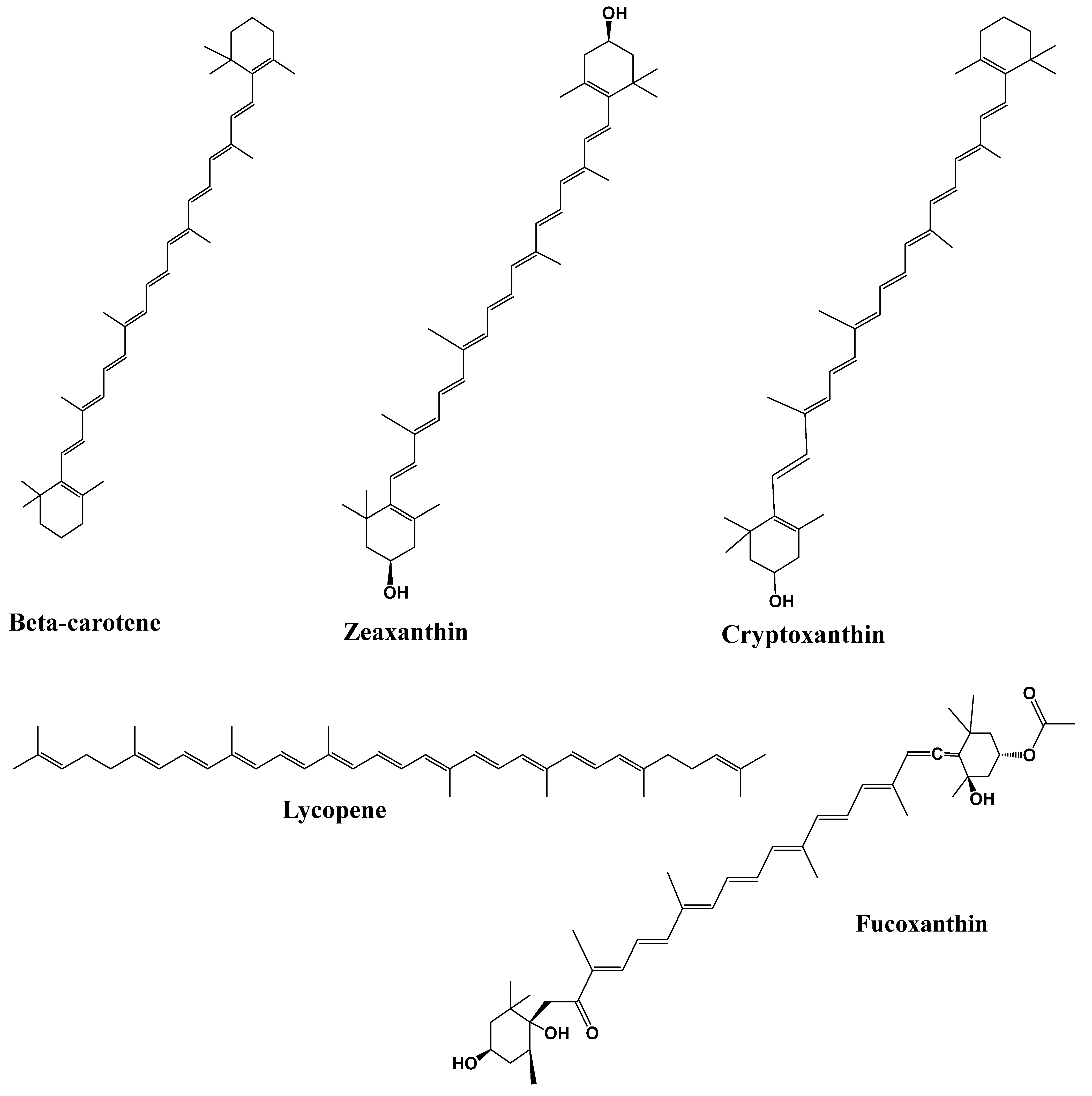
4.3. Terpenoids
4.4. Saponins
4.5. Anthocyanins
Cyanidin
5. Application of Phytonutrient-Based Anti-Obesity Food Supplement Products and Market Trends
6. Future Perspectives and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Apovian, C.M. Obesity: Definition, comorbidities, causes, and burden. Am. J. Manag. Care 2016, 22, 176–185. [Google Scholar]
- Pati, S.; Irfan, W.; Jameel, A.; Ahmed, S.; Shahid, R.K. Obesity and cancer: A current overview of epidemiology, pathogenesis, outcomes, and management. Cancers 2023, 15, 485. [Google Scholar] [CrossRef]
- Llewellyn, A.; Simmonds, M.; Owen, C.G.; Woolacott, N. Childhood obesity as a predictor of morbidity in adulthood: A systematic review and meta-analysis. Obes. Rev. 2016, 17, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Engin, A.B.; Engin, A. The definition and prevalence of obesity and metabolic syndrome. Adv. Exp. Med. Biol. 2017, 960, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Urasaki, Y.; Le, T.T. Functional Complementation of Anti-Adipogenic Phytonutrients for Obesity Prevention and Management. Nutrients 2022, 14, 4325. [Google Scholar] [CrossRef] [PubMed]
- Shubham, K.; Anukiruthika, T.; Dutta, S.; Kashyap, A.V.; Moses, J.A.; Anandharamakrishnan, C. Iron deficiency anemia: A comprehensive review on iron absorption, bioavailability and emerging food fortification approaches. Trends Food Sci. Technol. 2020, 99, 58–75. [Google Scholar] [CrossRef]
- Tran, T.; Bønløkke, P.; Rodríguez-Rodríguez, C.; Nosrati, Z.; Esquinas, P.L.; Borkar, N.; Plum, J.; Strindberg, S.; Karagiozov, S.; Rades, T.; et al. Using in vitro lipolysis and SPECT/CT in vivo imaging to understand oral absorption of fenofibrate from lipid-based drug delivery systems. J. Control. Release 2020, 317, 375–384. [Google Scholar] [CrossRef]
- Ogden, C.L.; Carroll, M.D.; Kit, B.K.; Flegal, K.M. Prevalence of childhood and adult obesity in the United States, 2011–2012. JAMA 2014, 311, 806–814. [Google Scholar] [CrossRef]
- Hendriks, S.L.; Viljoen, A.; Marais, D.; Wenhold, F.A.; McIntyre, A.M.; Ngidi, M.S.; Annandale, J.G.; Stewart, D. Considerations for the design of nutrition-sensitive production programmes in rural South Africa. BMC Public Health 2020, 20, 1383. [Google Scholar] [CrossRef] [PubMed]
- Tharifkhan, S.A.; Perumal, A.B.; Elumalai, A.; Moses, J.A.; Anandharamakrishnan, C. Improvement of nutrient bioavailability in millets: Emphasis on the application of enzymes. J. Sci. Food Agric. 2021, 101, 4869–4878. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Mu, M.; Liu, K.; He, Y. Screen time and childhood overweight/obesity: A systematic review and meta-analysis. Child Care Health Dev. 2019, 45, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Neve, K.L.; Isaacs, A. How does the food environment influence people engaged in weight management? A systematic review and thematic synthesis of the qualitative literature. Obes. Rev. 2022, 23, e13398. [Google Scholar] [CrossRef]
- Rebello, C.J.; Greenway, F.L.; Finley, J.W. A review of the nutritional value of legumes and their effects on obesity and its related co-morbidities. Obes. Rev. 2014, 15, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Plow, M.A.; Moore, S.; Husni, M.E.; Kirwan, J.P. A systematic review of behavioural techniques used in nutrition and weight loss interventions among adults with mobility-impairing neurological and musculoskeletal conditions. Obes. Rev. 2014, 15, 945–956. [Google Scholar] [CrossRef]
- Anderson, E.; Durstine, J.L. Physical activity, exercise, and chronic diseases: A brief review. Sports Med. Health Sci. 2014, 1, 3–10. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Yadav, P.; Vashishth, D.; Sharma, K.; Kumar, A.; Chahal, J.; Dalal, S.; Kataria, S.K. A review on obesity management through natural compounds and a green nanomedicine-based approach. Molecules 2021, 26, 3278. [Google Scholar] [CrossRef]
- Bibi, R.; Jahan, S.; Afsar, T.; Almajwal, A.; Hammadeh, M.E.; Alruwaili, N.W.; Ra/zak, S.; Amor, H. The influence of paternal overweight on sperm chromatin integrity, fertilization rate and pregnancy outcome among males attending fertility clinic for IVF/ICSI treatment. BMC Pregnancy Childbirth 2022, 22, 620. [Google Scholar] [CrossRef]
- Orgeron III, R.; Pope, J.; Erickson, D.; Green, V. Phytonutrients: A Potential Role in Obesity (P08-047-19). Curr. Dev. Nutr. 2019, 3, nzz044.P08-047-19. [Google Scholar] [CrossRef]
- World Obesity. Available online: https://data.worldobesity.org/maps/?area=trends&group=M&year=2020 (accessed on 15 July 2023).
- Martin, C.; Zhang, Y.; Tonelli, C.; Petroni, K. Plants, diet, and health. Annu. Rev. Plant Biol. 2013, 64, 19–46. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.S.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Nisar, A.; Jagtap, S.; Vyavahare, S.; Deshpande, M.; Harsulkar, A.; Ranjekar, P.; Prakash, O. Phytochemicals in the treatment of inflammation-associated diseases: The journey from preclinical trials to clinical practice. Front. Pharmacol. 2013, 14, 1177050. [Google Scholar] [CrossRef] [PubMed]
- Issaoui, M.; Delgado, A.M.; Caruso, G.; Micali, M.; Barbera, M.; Atrous, H.; Ouslati, A.; Chammem, N. Phenols, flavors, and the mediterranean diet. J. AOAC Int. 2020, 103, 915–924. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Morze, J.; Hoffmann, G. Mediterranean diet and health status: Active ingredients and pharmacological mechanisms. Br. J. Pharmacol. 2020, 177, 1241–1257. [Google Scholar] [CrossRef]
- Clodoveo, M.L.; Camposeo, S.; De Gennaro, B.; Pascuzzi, S.; Roselli, L. In the ancient world, virgin olive oil was called “liquid gold” by Homer and “the great healer” by Hippocrates. Why has this mythic image been forgotten? Food Res. Int. 2014, 62, 1062–1068. [Google Scholar] [CrossRef]
- Delgado-Floody, P.; Alvarez, C.; Caamaño-Navarrete, F.; Jerez-Mayorga, D.; Latorre-Román, P. Influence of Mediterranean diet adherence, physical activity patterns, and weight status on cardiovascular response to cardiorespiratory fitness test in Chilean school children. Nutrition 2020, 71, 110621. [Google Scholar] [CrossRef]
- Servili, M.; Sordini, B.; Esposto, S.; Urbani, S.; Veneziani, G.; Maio, I.D.; Selvaggini, R.; Taticchi, A. Biological activities of phenolic compounds of extra virgin olive oil. Antioxidants 2013, 3, 1–23. [Google Scholar] [CrossRef]
- Aloo, S.O.; Ofosu, F.K.; Kim, N.H.; Kilonzi, S.M.; Oh, D.H. Insights on Dietary Polyphenols as Agents against Metabolic Disorders: Obesity as a Target Disease. Antioxidants 2023, 12, 416. [Google Scholar] [CrossRef]
- Jin, T.; Song, Z.; Weng, J.; Fantus, I.G. Curcumin and other dietary polyphenols: Potential mechanisms of metabolic actions and therapy for diabetes and obesity. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E201–E205. [Google Scholar] [CrossRef]
- Omar, S.H. Oleuropein in olive and its pharmacological effects. Sci. Pharm. 2010, 78, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Galbete, C.; Kröger, J.; Jannasch, F.; Iqbal, K.; Schwingshackl, L.; Schwedhelm, C.; Weikert, C.; Boeing, H.; Schulze, M.B. Nordic diet, Mediterranean diet, and the risk of chronic diseases: The EPIC-Potsdam study. BMC Med. 2018, 16, 99. [Google Scholar] [CrossRef] [PubMed]
- Willcox, D.C.; Scapagnini, G.; Willcox, B.J. Healthy aging diets other than the Mediterranean: A focus on the Okinawan diet. Mech. Ageing Dev. 2014, 136, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Muroyama, K.; Yamamoto, Y.; Murosaki, S. Erratum to: Oral intake of a combination of glucosyl hesperidin and caffeine elicits an antiobesity effect in healthy, moderately obese subjects: A randomized double-blind placebo-controlled trial. Nutr. J. 2017, 16, 28. [Google Scholar] [CrossRef] [PubMed]
- Darwiche, G.; Höglund, P.; Roth, B.; Larsson, E.; Sjöberg, T.; Wohlfart, B.; Steen, S.; Ohlsson, B. An Okinawan-based Nordic diet improves anthropometry, metabolic control, and health-related quality of life in Scandinavian patients with type 2 diabetes: A pilot trial. Food Nutr. Res. 2016, 60, 32594. [Google Scholar] [CrossRef]
- Kårlund, A.; Kolehmainen, M.; Landberg, R.; Poutanen, K. Traditional and new sources of grain protein in the healthy and sustainable Nordic diet. J. Cereal Sci. 2022, 105, 103462. [Google Scholar] [CrossRef]
- FAO/WHO. FAOSTAT. 2018. Available online: https://www.fao.org/faostat/en/#data/FBS (accessed on 20 May 2023).
- Sang, S.; Chu, Y. Whole grain oats, more than just a fiber: Role of unique phytochemicals. Mol. Nutr. Food Res. 2017, 61, 1600715. [Google Scholar] [CrossRef]
- Cena, H.; Calder, P.C. Defining a healthy diet: Evidence for the role of contemporary dietary patterns in health and disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef]
- Turner-McGrievy, G.M.; Wilcox, S.; Frongillo, E.A.; Murphy, E.A.; Hutto, B.; Wilson, M.; Davey, M.; Bernhart, J.A.; Okpara, N.; Bailey, S.; et al. Effect of a Plant-Based vs. Omnivorous Soul Food Diet on Weight and Lipid Levels Among African American Adults: A Randomized Clinical Trial. JAMA Netw. 2023, 6, e2250626. [Google Scholar] [CrossRef]
- Bello, A. The Role of Diet in the Control of Hypertension: What Africans Should Know. Available online: https://www.datelinehealthafrica.org/the-role-of-diet-in-the-control-of-hypertension-what-africans-should-know (accessed on 14 September 2023).
- Kuehn, B.M. Heritage diets and culturally appropriate dietary advice may help combat chronic diseases. JAMA 2019, 322, 2271–2273. [Google Scholar] [CrossRef]
- Medagama, A.; Widanapathirana, H. A traditional Asian diet modified to meet nutritional requirements of diabetes, has anything changed? A cross-sectional dietary survey. BMC Nutr. 2015, 1, 4–9. [Google Scholar] [CrossRef]
- Kim, G.N.; Shin, M.R.; Shin, S.H.; Lee, A.R.; Lee, J.Y.; Seo, B.I.; Kim, M.Y.; Kim, T.H.; Noh, J.S.; Rhee, M.H.; et al. Study of antiobesity effect through inhibition of pancreatic lipase activity of Diospyros kaki fruit and Citrus unshiu peel. BioMed Res. Int. 2016, 2016, 1723042. [Google Scholar] [CrossRef] [PubMed]
- Marchiori, D.; Papies, E.K. A brief mindfulness intervention reduces unhealthy eating when hungry, but not the portion size effect. Appetite 2014, 75, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.J.; Park, S.H.; Choi, E.K.; Cha, Y.S.; Cho, B.H.; Kim, Y.G.; Kim, M.G.; Song, W.O.; Park, T.S.; Ko, J.K.; et al. Beneficial Effects of Korean Traditional Diets in Hypertensive and Type 2 Diabetic Patients. J. Med. Food 2014, 17, 161–171. [Google Scholar] [CrossRef]
- Ma, G. Food, eating behavior, and culture in Chinese society. J. Ethn. Foods 2015, 2, 195–199. [Google Scholar] [CrossRef]
- Leonetti, F.; Liguori, A.; Petti, F.; Rughini, S.; Silli, L.; Liguori, S.; Bangrazi, S. Effects of basic traditional Chinese diet on body mass index, lean body mass, and eating and hunger behaviours in overweight or obese individuals. J. Tradit. Chin. Med. ChungTsa Chih Ying Wen Pan 2016, 36, 456–463. [Google Scholar] [CrossRef]
- Niu, K.; Momma, H.; Kobayashi, Y.; Guan, L.; Chujo, M.; Otomo, A.; Ouchi, E.; Nagatomi, R. The traditional Japanese dietary pattern and longitudinal changes in cardiovascular disease risk factors in apparently healthy Japanese adults. Eur. J. Nutr. 2016, 55, 267–279. [Google Scholar] [CrossRef]
- Milinovic, J.; Mata, P.; Diniz, M.; Noronha, J.P. Umami taste in edible seaweeds: The current comprehension and perception. Int. J. Gastron. Food Sci. 2021, 23, 100301. [Google Scholar] [CrossRef]
- Campbell, A.P. DASH Eating Plan: An Eating Pattern for Diabetes Management. Diabetes Spectr. 2017, 30, 76–81. [Google Scholar] [CrossRef]
- Salehi-Abargouei, A.; Maghsoudi, Z.; Shirani, F.; Azadbakht, L. Effects of Dietary Approaches to Stop Hypertension (DASH)-style diet on fatal or nonfatal cardiovascular diseases—Incidence: A systematic review and meta-analysis on observational prospective studies. Nutrition 2013, 29, 611–618. [Google Scholar] [CrossRef]
- Bohn, T.; McDougall, G.J.; Alegría, A.; Alminger, M.; Arrigoni, E.; Aura, A.M.; Brito, C.; Cilla, A.; El, S.N.; Karakaya, S.; et al. Mind the gap—Deficits in our knowledge of aspects impacting the bioavailability of phytochemicals and their metabolites—A position paper focusing on carotenoids and polyphenols. Mol. Nutr. Food Res. 2015, 59, 1307–1323. [Google Scholar] [CrossRef] [PubMed]
- Niaz, K.; Shah, M.A.; Khan, F.; Saleem, U.; Vargas, C.; Panichayupakaranant, P. Bioavailability and safety of phytonutrients. In Phytonutrients in Food; Elsevier: Amsterdam, The Netherlands, 2020; pp. 117–136. [Google Scholar] [CrossRef]
- Beane, K.E.; Redding, M.C.; Wang, X.; Pan, J.H.; Le, B.; Cicalo, C.; Young, S.J.; Kim, J.; Lee, J.H.; Shin, E.C.; et al. Effects of dietary fibers, micronutrients, and phytonutrients on gut microbiome: A review. Appl. Biol. Chem. 2021, 64, 36. [Google Scholar] [CrossRef]
- Thakur, N.; Raigond, P.; Singh, Y.; Mishra, T.; Singh, B.; Lal, M.K.; Dutt, S. Recent updates on bioaccessibility of phytonutrients. Trends Food Sci. Technol. 2020, 97, 366–380. [Google Scholar] [CrossRef]
- Martin, C. The Interface between Plant Metabolic Engineering and Human Health. Curr. Opin. Biotechnol. 2013, 24, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.M.; Barraj, L.M.; Herman, D.; Bi, X.; Cheatham, R.; Randolph, R.K. Phytonutrient intake by adults in the United States in relation to fruit and vegetable consumption. J. Acad. Nutr. Diet. 2012, 112, 222–229. [Google Scholar] [CrossRef]
- Elsamanoudy, A.Z.; Neamat-Allah, M.A.M.; Mohammad, F.A.H.; Hassanien, M.; Nada, H.A. The role of nutrition related genes and nutrigenetics in understanding the pathogenesis of cancer. J. Microsc. Ultrastruct. 2016, 4, 115. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Mengxue, G.; Guangbo, K.; He, H. The Potential Role of Phytonutrients Flavonoids Influencing Gut Microbiota in the Prophylaxis and Treatment of Inflammatory Bowel Disease. Front. Nutr. 2021, 8, 798038. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Gary, W. Nutrients and Phytochemicals: From Bioavailability to Bioefficacy beyond Antioxidants. Curr. Opin. Biotechnol. 2008, 19, 73–82. [Google Scholar] [CrossRef]
- Liu, R.H. Health Benefits of Fruit and Vegetables Are from Additive and Synergistic Combinations of Phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.; Zhang, B.; Deng, Z. The Synergistic and Antagonistic Antioxidant Interactions of Dietary Phytochemical Combinations. Crit. Rev. Food Sci. Nutr. 2003, 62, 5658–5677. [Google Scholar] [CrossRef]
- de Kok, T.M.; van Breda, S.G.; Manson, M.M. Mechanisms of combined action of different chemopreventive dietary compounds: A review. Eur. J. Nutr. 2008, 47, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Esquivel, M.K. Nutrition Benefits and Considerations for Whole Foods Plant-Based Eating Patterns. Am. J. Lifestyle Med. 2022, 16, 284–290. [Google Scholar] [CrossRef]
- Ribas-Agustí, A.; Martín-Belloso, O.; Soliva-Fortuny, R.; Elez-Martínez, P. Food Processing Strategies to Enhance Phenolic Compounds Bioaccessibility and Bioavailability in Plant-Based Foods. Crit. Rev. Food Sci. Nutr. 2018, 58, 2531–2548. [Google Scholar] [CrossRef] [PubMed]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of in vitro digestion on composition, bioaccessibility and antioxidant activity of food polyphenols—A non-systematic review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef] [PubMed]
- Alminger, M.; Aura, A.M.; Bohn, T.; Dufour, C.; El, S.N.; Gomes, A.; Karakaya, S.; Martínez-Cuesta, M.C.; McDougall, G.J.; Requena, T.; et al. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413–436. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Yeo, J.D. Insoluble-Bound Phenolics in Food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef]
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Złotek, U.; Zielińska, E. Digestion and bioavailability of bioactive phytochemicals. Int. J. Food Sci. Technol. 2017, 52, 291–305. [Google Scholar] [CrossRef]
- Santhiravel, S.; Bekhit, A.E.D.A.; Mendis, E.; Jacobs, J.L.; Dunshea, F.R.; Rajapakse, N.; Ponnampalam, E.N. The impact of plant phytochemicals on the gut microbiota of humans for a balanced life. Int. J. Mol. Sci. 2022, 23, 8124. [Google Scholar] [CrossRef]
- McGhie, T.K.; Walton, M.C. The bioavailability and absorption of anthocyanins: Towards a better understanding. Mol. Nutr. Food Res. 2022, 51, 702–713. [Google Scholar] [CrossRef]
- Walle, T. Absorption and Metabolism of Flavonoids. Free. Radic. Biol. Med. 2004, 36, 829–837. [Google Scholar] [CrossRef]
- Kan, J.; Wu, F.; Wang, F.; Zheng, J.; Cheng, J.; Li, Y.; Yang, Y.; Du, J. Phytonutrients: Sources, bioavailability, interaction with gut microbiota, and their impacts on human health. Front. Nutr. 2022, 9, 960309. [Google Scholar] [CrossRef] [PubMed]
- Sharif, M.K.; Zahid, A.; Shah, F.H. Role of food product development in increased food consumption and value addition. In Food Processing for Increased Quality and Consumption; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 455–479. [Google Scholar] [CrossRef]
- Xiao, J.Y.C.; Huang, Q. Edible Nanoencapsulation Vehicles for Oral Delivery of Phytochemicals: A Perspective Paper. J. Agric. Food Chem. 2017, 65, 6727–6735. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Li, H.; Shahidi, F.; Luo, T.; Deng, Z. Interactions among dietary phytochemicals and nutrients: Role of cell membranes. Trends Food Sci. Technol. 2022, 124, 38–50. [Google Scholar] [CrossRef]
- Harahap, I.A.; Sobral, M.M.C.; Casal, S.; Pinho, S.; Faria, M.A.; Suliburska, J.; Ferreira, I.M. Fat oxidation of fatty fish vs. meat meal diets under in vitro standardized semi-dynamic gastric digestion. Front. Nutr. 2022, 9, 901006. [Google Scholar] [CrossRef] [PubMed]
- Kritchevsky, D. Dietary Fibre in Health and Disease. In Advanced Dietary Fibre Technology; Wiley: Hoboken, NJ, USA, 2000; pp. 147–161. [Google Scholar] [CrossRef]
- Shahidi, F.; Pan, Y. Influence of Food Matrix and Food Processing on the Chemical Interaction and Bioaccessibility of Dietary Phytochemicals: A Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 6421–6445. [Google Scholar] [CrossRef]
- Palafox-Carlos, H.; Ayala-Zavala, J.F.; González-Aguilar, G.A. The role of dietary fiber in the bioaccessibility and bioavailability of fruit and vegetable antioxidants. J. Food Sci. 2011, 76, R6–R15. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Whole grains and phenolic acids: A review on bioactivity, functionality, health benefits and bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef]
- Tosh, S.M.; Bordenave, N. Emerging science on benefits of whole grain oat and barley and their soluble dietary fibers for heart health, glycemic response, and gut microbiota. Nutr. Rev. 2021, 78, 13–20. [Google Scholar] [CrossRef]
- McClements, D.J. Advances in Nanoparticle and Microparticle Delivery Systems for Increasing the Dispersibility, Stability, and Bioactivity of Phytochemicals. Biotechnol. Adv. 2020, 38, 107287. [Google Scholar] [CrossRef]
- Padayachee, A.; Day, L.; Howell, K.; Gidley, M.J. Complexity and health functionality of plant cell wall fibers from fruits and vegetables. Crit. Rev. Food Sci. Nutr. 2017, 57, 59–81. [Google Scholar] [CrossRef]
- Borel, P.; Dangles, O.; Kopec, R.E. Fat-soluble vitamin and phytochemical metabolites: Production, gastrointestinal absorption, and health effects. Prog. Lipid Res. 2023, 90, 101220. [Google Scholar] [CrossRef] [PubMed]
- Mashurabad, P.C.; Palika, R.; Jyrwa, Y.W.; Bhaskarachary, K.; Pullakhandam, R. Dietary fat composition, food matrix and relative polarity modulate the micellarization and intestinal uptake of carotenoids from vegetables and fruits. J. Food Sci. Technol. 2017, 54, 333–341. [Google Scholar] [CrossRef] [PubMed]
- McClements, D.J.; Xiao, H. Excipient foods: Designing food matrices that improve the oral bioavailability of pharmaceuticals and nutraceuticals. Food Funct. 2014, 5, 1320–1333. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Yu, H.; Ru, Q. Bioavailability and delivery of nutraceuticals using nanotechnology. J. Food Sci. 2010, 75, R50–R57. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Lin, Q.; Zhao, H.; Li, X.; Sang, S.; McClements, D.J.; Long, J.; Jin, Z.; Wang, J.; Qiu, C. Bioaccessibility and bioavailability of phytochemicals: Influencing factors, improvements, and evaluations. Food Hydrocoll. 2022, 135, 108165. [Google Scholar] [CrossRef]
- Porrini, M.; Patrizia, R. Factors Influencing the Bioavailability of Antioxidants in Foods: A Critical Appraisal. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 647–650. [Google Scholar] [CrossRef]
- Boronat, A.; Rodriguez-Morató, J.; Serreli, G.; Fitó, M.; Tyndale, R.F.; Deiana, M.; de la Torre, R. Contribution of biotransformations carried out by the microbiota, drug-metabolizing enzymes, and transport proteins to the biological activities of phytochemicals found in the diet. Adv. Nutr. 2021, 12, 2172–2189. [Google Scholar] [CrossRef]
- Borel, P.; Charles, D. Bioavailability of Fat-Soluble Vitamins and Phytochemicals in Humans: Effects of Genetic Variation. Annu. Rev. Nutr. 2018, 38, 69–96. [Google Scholar] [CrossRef]
- Hoda, M.; Hemaiswarya, S.; Doble, M. Role of phenolic phytochemicals in diabetes management. Role of Phenolic Phytochemicals in Diabetes Management; Springer: Singapore, 2019; pp. 159–173. [Google Scholar] [CrossRef]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Vadhanam, M.V. Bioavailability of phytochemicals and its enhancement by drug delivery systems. Cancer Lett. 2013, 334, 133–141. [Google Scholar] [CrossRef]
- Qin, Z.; Wang, P.; Duan, S.; Wan, X.; Xing, H.; Yang, J.; Zhang, X.; Yao, Z.; Yao, X. Potential determinants for metabolic fates and inhibitory effects of isobavachalcone involving in human cytochrome P450, UDP-glucuronosyltransferase enzymes, and efflux transporters. J. Pharm. Sci. 2021, 110, 2285–2294. [Google Scholar] [CrossRef]
- vel Szic, K.S.; Declerck, K.; Vidaković, M.; Vanden Berghe, W. From inflammaging to healthy aging by dietary lifestyle choices: Is epigenetics the key to personalized nutrition? Clin. Epigenet. 2015, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Giovanni, S. Interactions between Dietary Polyphenols and Aging Gut Microbiota: A Review. BioFactors 2022, 48, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Bhatia, S.; Chunduri, V.; Kaur, S.; Sharma, S.; Kapoor, P.; Kumari, A.; Garg, M. Pathogenesis of celiac disease and other gluten related disorders in wheat and strategies for mitigating them. Front. Nutr. 2020, 7, 6. [Google Scholar] [CrossRef]
- Theethira, T.G.; Dennis, M. Celiac disease and the gluten-free diet: Consequences and recommendations for improvement. Dig. Dis. 2015, 33, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Caeiro, C.; Pragosa, C.; Cruz, M.C.; Pereira, C.D.; Pereira, S.G. The role of pseudocereals in celiac disease: Reducing nutritional deficiencies to improve well-being and health. J. Nutr. Metab. 2022, 2022, 8502169. [Google Scholar] [CrossRef]
- Shabbir, M.A.; Mehak, F.; Khan, Z.M.; Ahmad, W.; Khan, M.R.; Zia, S.; Rahaman, A.; Aadil, R.M. Interplay between ceramides and phytonutrients: New insights in metabolic syndrome. Trends Food Sci. Technol. 2021, 111, 483–494. [Google Scholar] [CrossRef]
- Rémond, D.; Shahar, D.R.; Gille, D.; Pinto, P.; Kachal, J.; Peyron, M.A.; Dos Santos, C.N.; Walther, B.; Bordoni, A.; Dupont, D.; et al. Understanding the gastrointestinal tract of the elderly to develop dietary solutions that prevent malnutrition. Oncotarget 2015, 6, 13858–13898. [Google Scholar] [CrossRef]
- Shahmohammadi, H.A.; Hosseini, S.A.; Hajiani, E.; Malehi, A.S.; Alipour, M. Effects of green coffee bean extract supplementation on patients with non-alcoholic fatty liver disease: A randomized clinical trial. Hepat. Mon. 2017, 17, e12299. [Google Scholar] [CrossRef]
- Gerges, S.H.; Wahdan, S.A.; Elsherbiny, D.A.; El-Demerdash, E. Non-alcoholic fatty liver disease: An overview of risk factors, pathophysiological mechanisms, diagnostic procedures, and therapeutic interventions. Life Sci. 2021, 271, 119220. [Google Scholar] [CrossRef]
- Ota, S.; Morita, A.; Ohnuki, S.; Hirata, A.; Sekida, S.; Okuda, K.; Ohya, Y.; Kawano, S. Carotenoid dynamics and lipid droplet containing astaxanthin in response to light in the green alga Haematococcus pluvialis. Sci. Rep. 2018, 8, 5617. [Google Scholar] [CrossRef]
- Battino, M.; Giampieri, F.; Cianciosi, D.; Ansary, J.; Chen, X.; Zhang, D.; Gil, E.; Forbes-Hernández, T. The roles of strawberry and honey phytochemicals on human health: A possible clue on the molecular mechanisms involved in the prevention of oxidative stress and inflammation. Phytomedicine 2021, 86, 153170. [Google Scholar] [CrossRef] [PubMed]
- Mercali, G.D.; Jaeschke, D.P.; Tessaro, I.C.; Marczak, L.D.F. Study of vitamin C degradation in acerola pulp during ohmic and conventional heat treatment. LWT-Food Sci. Technol. 2012, 47, 91–95. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Aoyagi, H. Thermal and UV degradation kinetics of water-soluble extracellular pigment produced by Talaromyces purpurogenus. Food Bioprocess Technol. 2022, 15, 606–619. [Google Scholar] [CrossRef]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agric. 2014, 94, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Chen, J.; Lv, F.; Chen, S.; Chen, J.; Liu, D.; Ye, X. Domestic cooking methods affect the phytochemical composition and antioxidant activity of purple-fleshed potatoes. Food Chem. 2016, 197, 1264–1270. [Google Scholar] [CrossRef]
- Ioannou, I.; Chekir, L.; Ghoul, M. Effect of heat treatment and light exposure on the antioxidant activity of flavonoids. Processes 2020, 8, 1078. [Google Scholar] [CrossRef]
- Shahidi, F.; Chandrasekara, A.; Zhong, Y. Bioactive phytochemicals in vegetables. In Handbook of Vegetables and Vegetable Processing; Wiley: Hoboken, NJ, USA, 2018; Volume 1–2, pp. 181–222. [Google Scholar] [CrossRef]
- Thamkaew, G.; Sjöholm, I.; Galindo, F.G. A review of drying methods for improving the quality of dried herbs. Crit. Rev. Food Sci. Nutr. 2021, 61, 1763–1786. [Google Scholar] [CrossRef]
- Tiwari, U.; Cummins, E. Factors influencing levels of phytochemicals in selected fruit and vegetables during pre-and post-harvest food processing operations. Food Res. Int. 2013, 50, 497–506. [Google Scholar] [CrossRef]
- Maqsood, S.; Benjakul, S.; Abushelaibi, A.; Alam, A. Phenolic compounds and plant phenolic extracts as natural antioxidants in prevention of lipid oxidation in seafood: A detailed review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1125–1140. [Google Scholar] [CrossRef]
- Robards, K.; Prenzler, P.D.; Tucker, G.; Swatsitang, P.; Glover, W. Phenolic compounds and their role in oxidative processes in fruits. Food Chem. 1999, 66, 401–436. [Google Scholar] [CrossRef]
- Sommano, S.R.; Chanasut, U.; Kumpoun, W. Enzymatic browning and its amelioration in fresh-cut tropical fruits. In Fresh-Cut Fruits and Vegetables; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 51–76. [Google Scholar] [CrossRef]
- Poljsak, B. Strategies for Reducing or Preventing the Generation of Oxidative Stress. Oxidative Med. Cell. Longev. 2011, 2011, 194586. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, S. Current Evidence on the Effect of Dietary Polyphenols Intake on Brain Health. Curr. Nutr. Food Sci. 2020, 16, 1170–1182. [Google Scholar] [CrossRef]
- Jabeen, N.S.; Kiruthiga, V.; Vinodhini, A.; Rudrapal, M. Herbs, Spices, and Dietary Constituents as Sources of Phytoantioxidants. In Phytoantioxidants and Nanotherapeutics; Wiley: Hoboken, NJ, USA, 2022; pp. 55–76. [Google Scholar] [CrossRef]
- Duarte, L.; Gasaly, N.; Poblete-Aro, C.; Uribe, D.; Echeverria, F.; Gotteland, M.; Garcia-Diaz, D.F. Polyphenols and their anti-obesity role mediated by the gut microbiota: A comprehensive review. Rev. Endocr. Metab. Disord. 2021, 22, 367–388. [Google Scholar] [CrossRef] [PubMed]
- Boccellino, M.; D’Angelo, S. Anti-obesity effects of polyphenol intake: Current status and future possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef] [PubMed]
- Jack, B.U.; Malherbe, C.J.; Mamushi, M.; Muller, C.J.; Joubert, E.; Louw, J.; Pheiffer, C. Adipose tissue as a possible therapeutic target for polyphenols: A case for Cyclopia extracts as anti-obesity nutraceuticals. Biomed. Pharmacother. 2019, 120, 109439. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, X. Interactions of tea polyphenols with intestinal microbiota and their implication for anti-obesity. J. Sci. Food Agric. 2020, 100, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.K.; Abdal Dayem, A.; Han, J.; Yin, Y.; Kim, K.; Saha, S.K.; Yang, G.M.; Choi, H.Y.; Cho, S.G. Molecular mechanisms of the anti-obesity and anti-diabetic properties of flavonoids. Int. J. Mol. Sci. 2016, 17, 569. [Google Scholar] [CrossRef]
- Song, D.; Cheng, L.; Zhang, X.; Wu, Z.; Zheng, X. The modulatory effect and the mechanism of flavonoids on obesity. J. Food Biochem. 2019, 43, e12954. [Google Scholar] [CrossRef]
- Oliveira, A.K.D.S.; de Oliveira e Silva, A.M.; Pereira, R.O.; Santos, A.S.; Barbosa Junior, E.V.; Bezerra, M.T.; Barreto, R.S.S.; Quintans-Junior, L.J.; Quintans, J.S. Anti-obesity properties and mechanism of action of flavonoids: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 7827–7848. [Google Scholar] [CrossRef]
- Varshney, R.; Mishra, R.; Das, N.; Sircar, D.; Roy, P. A comparative analysis of various flavonoids in the regulation of obesity and diabetes: An in vitro and in vivo study. J. Funct. Foods 2019, 59, 194–205. [Google Scholar] [CrossRef]
- Kurniawan, R.; Nurkolis, F.; Taslim, N.A.; Subali, D.; Surya, R.; Gunawan, W.B.; Alisaputra, D.; Mayulu, N.; Salindeho, N.; Kim, B. Carotenoids Composition of Green Algae Caulerpa racemosa and Their Antidiabetic, Anti-Obesity, Antioxidant, and Anti-Inflammatory Properties. Molecules 2023, 28, 3267. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, K. Anti-Obesity Therapy by Food Component: Unique Activity of Marine Carotenoid, Fucoxanthin. Obes. Control. Ther. Open Access 2013, 1, 3267. [Google Scholar] [CrossRef]
- Kim, H.J.; Koo, K.A.; Park, W.S.; Kang, D.M.; Kim, H.S.; Lee, B.Y.; Woo, D.K.; Kwak, S.S.; Ahn, M.J. Anti-obesity activity of anthocyanin and carotenoid extracts from color-fleshed sweet potatoes. J. Food Biochem. 2020, 44, e13438. [Google Scholar] [CrossRef]
- Stefania, D.S.; Clodoveo, M.L.; Cariello, M.; D’Amato, G.; Franchini, C.; Faienza, M.F.; Corbo, F. Polyphenols and obesity prevention: Critical insights on molecular regulation, bioavailability and dose in preclinical and clinical settings. Crit. Rev. Food Sci. Nutr. 2021, 61, 1804–1826. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Russo, G.L. The flavonoid quercetin in disease prevention and therapy: Facts and fancies. Biochem. Pharmacol. 2012, 83, 6–15. [Google Scholar] [CrossRef]
- Goktas, Z.; Zu, Y.; Abbasi, M.; Galyean, S.; Wu, D.; Fan, Z.; Wang, S. Recent advances in nanoencapsulation of phytochemicals to combat obesity and its comorbidities. J. Agric. Food Chem. 2020, 68, 8119–8131. [Google Scholar] [CrossRef]
- Pekkinen, J.; Rosa, N.N.; Savolainen, O.I.; Keski-Rahkonen, P.; Mykkänen, H.; Poutanen, K.; Micard, V.; Hanhineva, K. Disintegration of wheat aleurone structure has an impact on the bioavailability of phenolic compounds and other phytochemicals as evidenced by altered urinary metabolite profile of diet-induced obese mice. Nutr. Metab. 2014, 11, 1. [Google Scholar] [CrossRef]
- Azhar, Y.; Parmar, A.; Miller, C.N.; Samuels, J.S.; Rayalam, S. Phytochemicals as novel agents for the induction of browning in white adipose tissue. Nutr. Metab. 2016, 13, 89. [Google Scholar] [CrossRef]
- Mele, L.; Bidault, G.; Mena, P.; Crozier, A.; Brighenti, F.; Vidal-Puig, A.; Del Rio, D. Dietary (Poly) phenols, brown adipose tissue activation, and energy expenditure: A narrative review. Adv. Nutr. 2017, 8, 694–704. [Google Scholar] [CrossRef]
- Mopuri, R.; Islam, M.S. Medicinal plants and phytochemicals with anti-obesogenic potentials: A review. Biomed. Pharmacother. 2017, 89, 1442–1452. [Google Scholar] [CrossRef]
- Salvadó, M.J.; Casanova, E.; Fernández-Iglesias, A.; Arola, L.; Bladé, C. Roles of proanthocyanidin rich extracts in obesity. Food Funct. 2015, 6, 1053–1071. [Google Scholar] [CrossRef]
- Balaji, M.; Ganjayi, M.S.; Kumar, G.E.H.; Parim, B.N.; Mopuri, R.; Dasari, S. A review on possible therapeutic targets to contain obesity: The role of phytochemicals. Obes. Res. Clin. Pract. 2016, 10, 363–380. [Google Scholar] [CrossRef]
- Saha, M.; Mukherjee, S.; Das, S.; Chowdhury, M.; Das, M.; Sarkar, S.; Chatterjee, S. Phytonutrient screening and evaluation of in-vitro antibacterial activity of onion and garlic peels: A comparative study with the prospects of waste to wealth. Int. J. Herb. Med. 2022, 10, 39–44. [Google Scholar] [CrossRef]
- Carpene, C.; Gomez-Zorita, S.; Deleruyelle, S.; Carpene, M.A. Novel strategies for preventing diabetes and obesity complications with natural polyphenols. Curr. Med. Chem. 2015, 22, 150–164. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Jiang, H.; Jiang, B.; Zhang, T.; Cui, S.W.; Jin, Z. Phytonutrients for controlling starch digestion: Evaluation of grape skin extract. Food Chem. 2014, 145, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Shimizu, H.; Okazaki, Y.; Sakaguchi, H.; Taira, T.; Suzuki, T.; Chiji, H. Anthocyanin-rich phytochemicals from aronia fruits inhibit visceral fat accumulation and hyperglycemia in high-fat diet-induced dietary obese rats. J. Oleo Sci. 2015, 64, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Saad, B.; Ghareeb, B.; Kmail, A. Metabolic and epigenetics action mechanisms of antiobesity medicinal plants and phytochemicals. Evid. Based Complement. Altern. Med. 2021, 2021, 9995903. [Google Scholar] [CrossRef] [PubMed]
- Mounien, L.; Tourniaire, F.; Landrier, J.F. Anti-obesity effect of carotenoids: Directimpact on adipose tissue and adipose tissue-driven indirect effects. Nutrients 2019, 11, 1562. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, B.; Zhang, C.; Sun, G.; Sun, X. Effect of Panax notoginseng saponins and major anti-obesity components on weight loss. Front. Pharmacol. 2021, 11, 601751. [Google Scholar] [CrossRef]
- Singh, M.; Thrimawithana, T.; Shukla, R.; Adhikari, B. Managing obesity through natural polyphenols: A review. Future Foods 2020, 1–2, 100002. [Google Scholar] [CrossRef]
- Weng, G.; Duan, Y.; Zhong, Y.; Song, B.; Zheng, J.; Zhang, S.; Yin, Y.; Deng, J. Plant extracts in obesity: A role of gut microbiota. Front. Nutr. 2021, 8, 727951. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.C.; Hsieh, P.H.; Pan, M.H.; Ho, C.T. Cellular models for the evaluation of the antiobesity effect of selected phytochemicals from food and herbs. J. Food Drug Anal. 2017, 25, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Alkhalidy, H.; Moore, W.; Zhang, Y.; McMillan, R.; Wang, A.; Ali, M.; Suh, K.S.; Zhen, W.; Cheng, Z.; Jia, Z.; et al. Small molecule kaempferol promotes insulin sensitivity and preserved pancreatic β-cell mass in middle-aged obese diabetic mice. J. Diabetes Res. 2015, 2015, 532984. [Google Scholar] [CrossRef]
- Gourineni, V.; Shay, N.F.; Chung, S.; Sandhu, A.K.; Gu, L. Muscadine grape (Vitis rotundifolia) and wine phytochemicals prevented obesity-associated metabolic complications in C57BL/6J mice. J. Agric. Food Chem. 2012, 60, 7674–7681. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Miranda-Garcia, O.; Adamson, A.; Sasaki, G.; Shay, N.F. Development of obesity is reduced in high-fat fed mice fed whole raspberries, raspberry juice concentrate, and a combination of the raspberry phytochemicals ellagic acid and raspberry ketone. J. Berry Res. 2016, 6, 213–223. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, M.; Wu, T.; Dai, S.; Xu, J.; Zhou, Z. The anti-obesity effect of green tea polysaccharides, polyphenols and caffeine in rats fed with a high-fat diet. Food Funct. 2015, 6, 297–304. [Google Scholar] [CrossRef]
- Suzuki, T.; Pervin, M.; Goto, S.; Isemura, M.; Nakamura, Y. Beneficial effects of tea and the green tea catechin epigallocatechin-3-gallate on obesity. Molecules 2016, 21, 1305. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Chen, J.; Yi, K.; Peng, L.; Xie, J.; Gou, X.; Peng, T.; Tang, L. Phlorizin ameliorates obesity-associated endotoxemia and insulin resistance in high-fat diet-fed mice by targeting the gut microbiota and intestinal barrier integrity. Gut Microbes 2020, 12, 1842990. [Google Scholar] [CrossRef]
- Thilakarathna, S.H.; Rupasinghe, H.V.; Needs, P.W. Apple peel bioactive rich extracts effectively inhibit in vitro human LDL cholesterol oxidation. Food Chem. 2013, 138, 463–470. [Google Scholar] [CrossRef]
- Khanam, S.; Mishra, A.; Shahid, A.; Pujari, N.M. Therapeutic indication of Phloridzin: A new Gleam for metabolic disorders. Phytomed. Plus 2022, 2, 100200. [Google Scholar] [CrossRef]
- Yun, Y.R.; Park, B.Y.; Kim, S.H.; Jung, J.H. Antioxidant, anti-obesity, and anti-aging activities of Jeju citrus blended vinegar. Foods 2021, 10, 1441. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.K.; Cho, S.J.; Jung, U.J.; Ryu, R.; Choi, M.S. Phlorizin supplementation attenuates obesity, inflammation, and hyperglycemia in diet-induced obese mice fed a high-fat diet. Nutrients 2016, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Mao, L.; Xu, P.; Wang, Y. Effects of (−)-epigallocatechin gallate (EGCG) on energy expenditure and microglia-mediated hypothalamic inflammation in mice fed a high-fat diet. Nutrients 2018, 10, 1681. [Google Scholar] [CrossRef]
- Li, F.; Gao, C.; Yan, P.; Zhang, M.; Wang, Y.; Hu, Y.; Wu, X.; Wang, X.; Sheng, J. EGCG Reduces Obesity and White Adipose Tissue Gain Partly Through AMPK Activation in Mice. Front. Pharmacol. 2018, 9, 1366. [Google Scholar] [CrossRef]
- Chen, I.J.; Liu, C.Y.; Chiu, J.P.; Hsu, C.H. Therapeutic effect of high-dose green tea extract on weight reduction: A randomized, double-blind, placebo-controlled clinical trial. Clin. Nutr. 2016, 35, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Shahrajabian, M.H. Therapeutic potential of phenolic compounds in medicinal plants—Natural health products for human health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Dludla, P.V.; Nkambule, B.B.; Jack, B.; Mkandla, Z.; Mutize, T.; Silvestri, S.; Orlando, P.; Tiano, L.; Louw, J.; Mazibuko-Mbeje, S.E. Inflammation and oxidative stress in an obese state and the protective effects of gallic acid. Nutrients 2019, 11, 23. [Google Scholar] [CrossRef]
- Doan, K.V.; Ko, C.M.; Kinyua, A.W.; Yang, D.J.; Choi, Y.H.; Oh, I.Y.; Nguyen, N.M.; Ko, A.; Choi, J.W.; Joeng, Y.; et al. Gallic acid regulates body weight and glucose homeostasis through AMPK activation. Endocrinology 2015, 156, 157–168. [Google Scholar] [CrossRef]
- Tanaka, M.; Sugama, A.; Sumi, K.; Shimizu, K.; Kishimoto, Y.; Kondo, K.; Iida, K. Gallic acid regulates adipocyte hypertrophy and suppresses inflammatory gene expression induced by the paracrine interaction between adipocytes and macrophages in vitro and in vivo. Nutr. Res. 2020, 73, 58–66. [Google Scholar] [CrossRef]
- Peng, M.; Gao, Z.; Liao, Y.; Guo, J.; Shan, Y. Development of functional kiwifruit jelly with chenpi (FKJ) by 3D food printing technology and its anti-obesity and antioxidant potentials. Foods 2022, 11, 1894. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Russo, G.L.; Daglia, M.; Nabavi, S.M. Role of quercetin as an alternative for obesity treatment: You are what you eat! Food Chem. 2015, 179, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, L.; Fernández-Quintela, A.; Arias, N.; Portillo, M.P. Resveratrol: Anti-obesity mechanisms of action. Molecules 2014, 19, 18632–18655. [Google Scholar] [CrossRef]
- Zhao, L.; Cen, F.; Tian, F.; Li, M.J.; Zhang, Q.; Shen, H.Y.; Shen, X.C.; Du, J. Combination treatment with quercetin and resveratrol attenuates high fat diet-induced obesity and associated inflammation in rats via the AMPKα1/SIRT1 signaling pathway. Exp. Ther. Med. 2017, 14, 5942–5948. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, B.; Shen, J.; Wan, L.; Zhu, Y.; Yi, T.; Xiao, Z. The beneficial effects of quercetin, curcumin, and resveratrol in obesity. Oxidative Med. Cell. Longev. 2017, 2017, 1459497. [Google Scholar] [CrossRef] [PubMed]
- Konings, E.; Timmers, S.; Boekschoten, M.V.; Goossens, G.H.; Jocken, J.W.; Afman, L.A.; Müller, M.; Schrauwen, P.; Mariman, E.C.; Blaak, E. EThe effects of 30 days resveratrol supplementation on adipose tissue morphology and gene expression patterns in obese men. Int. J. Obes. 2014, 38, 470–473. [Google Scholar] [CrossRef]
- Mongioì, L.M.; Perelli, S.; Condorelli, R.A.; Barbagallo, F.; Crafa, A.; Cannarella, R.; Vignera, S.L.; Calogero, A.E. The role of resveratrol in human male fertility. Molecules 2021, 26, 2495. [Google Scholar] [CrossRef]
- Matsuura, H.N.; Fett-Neto, A.G. Plant alkaloids: Main features, toxicity, and mechanisms of action. Plant Toxins 2015, 2, 1–15. [Google Scholar] [CrossRef]
- Saad, B.; Zaid, H.; Shanak, S.; Kadan, S. Anti-diabetes and anti-obesity medicinal plants and phytochemicals. In Anti-diabetes and Anti-obesity Medicinal Plants and Phytochemicals; Springer: Cham, Switzerland, 2017; pp. 129–144. [Google Scholar] [CrossRef]
- Rupasinghe, H.V.; Sekhon-Loodu, S.; Mantso, T.; Panayiotidis, M.I. Phytochemicals in regulating fatty acid β-oxidation: Potential underlying mechanisms and their involvement in obesity and weight loss. Pharmacol. Ther. 2016, 165, 153–163. [Google Scholar] [CrossRef]
- Chow, Y.L.; Sogame, M.; Sato, F. 13-Methylberberine, a berberine analogue with stronger anti-adipogenic effects on mouse 3T3-L1 cells. Sci. Rep. 2016, 6, 38129. [Google Scholar] [CrossRef]
- Ma, C.; Wang, J.; Chu, H.; Zhang, X.; Wang, Z.; Wang, H.; Li, G. Purification and characterization of aporphine alkaloids from leaves of Nelumbo nucifera Gaertn and their effects on glucose consumption in 3T3-L1 adipocytes. Int. J. Mol. Sci. 2014, 15, 3481–3494. [Google Scholar] [CrossRef]
- Ahn, J.H.; Kim, E.S.; Lee, C.; Kim, S.; Cho, S.H.; Hwang, B.Y.; Lee, M.K. Chemical constituents from Nelumbo nucifera leaves and their anti-obesity effects. Bioorganic Med. Chem. Lett. 2013, 23, 3604–3608. [Google Scholar] [CrossRef] [PubMed]
- Gurung, P.; De, P. Spectrum of biological properties of cinchona alkaloids: A brief review. J. Pharmacogn. Phytochem. 2017, 6, 162–166. [Google Scholar]
- Huang, L.; Li, D.; Xu, Y.S.; Feng, Z.L.; Meng, F.C.; Zhang, Q.W.; Gan, L.S.; Lin, L.G. Clausoxamine, an alkaloid possessing a 1, 3-oxazine-4-one ring from the seeds of Clausena lansium and the anti-obesity effect of lansiumamide B. RSC Adv. 2017, 7, 46900–46905. [Google Scholar] [CrossRef]
- Jagtap, S.; Khare, P.; Mangal, P.; Kondepudi, K.K.; Bishnoi, M.; Bhutani, K.K. Effect of mahanimbine, an alkaloid from curry leaves, on high-fat diet-induced adiposity, insulin resistance, and inflammatory alterations. BioFactors 2017, 43, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S. Introductory Chapter: Terpenes and Terpenoids. In Terpenes and Terpenoids; IntechOpen: Benin City, Nigeria, 2018. [Google Scholar]
- Bonet, M.L.; Canas, J.A.; Ribot, J.; Palou, A. Carotenoids in adipose tissue biology and obesity. Carotenoids Nat. Biosynth. Regul. Funct. 2016, 79, 377–414. [Google Scholar] [CrossRef]
- Zhu, R.; Chen, B.; Bai, Y.; Miao, T.; Rui, L.; Zhang, H.; Xia, B.; Li, Y.; Gao, S.; Wang, X.D.; et al. Lycopene in protection against obesity and diabetes: A mechanistic review. Pharmacol. Res. 2020, 159, 104966. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qi, J.; Li, L. Phytochemicals as potential candidates to combat obesity via adipose non-shivering thermogenesis. Pharmacol. Res. 2019, 147, 104393. [Google Scholar] [CrossRef]
- Liu, M.; Liu, H.; Xie, J.; Xu, Q.; Pan, C.; Wang, J.; Wu, X.; Sanabil; Zheng, M.; Liu, J. Anti-obesity effects of zeaxanthin on 3T3-L1 preadipocyte and high fat induced obese mice. Food Funct. 2017, 8, 3327–3338. [Google Scholar] [CrossRef]
- Vinha, A.F.; Barreira, S.V.; Costa, A.S.; Alves, R.C.; Oliveira, M.B.P. Pre-meal tomato (Lycopersicon esculentum) intake can have anti-obesity effects in young women? Int. J. Food Sci. Nutr. 2014, 65, 1019–1026. [Google Scholar] [CrossRef]
- Jeepipalli, S.P.; Du, B.; Sabitaliyevich, U.Y.; Xu, B. New insights into potential nutritional effects of dietary saponins in protecting against the development of obesity. Food Chem. 2020, 318, 126474. [Google Scholar] [CrossRef]
- Leal-Díaz, A.M.; Noriega, L.G.; Torre-Villalvazo, I.; Torres, N.; Alemán-Escondrillas, G.; López-Romero, P.; Sánchez-Tapia, M.; Aguilar-López, M.; Furuzawa-Carballeda, J.; Velázquez-Villegas, L.A.; et al. Aguamiel concentrate from Agave salmiana and its extracted saponins attenuated obesity and hepatic steatosis and increased Akkermansia muciniphila in C57BL6 mice. Sci. Rep. 2016, 6, 34242. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Conforti, F.; Araniti, F.; Statti, G.A. Effects of saponins on lipid metabolism: A review of potential health benefits in the treatment of obesity. Molecules 2016, 21, 1404. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Kang, B.; Suh, H.J.; Choi, H.S. Red ginseng-derived saponin fraction suppresses the obesity-induced inflammatory responses via Nrf2-HO-1 pathway in adipocyte-macrophage co-culture system. Biomed. Pharmacother. 2018, 108, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, H.; Zhao, Y.; Zhu, H.; Cai, E.; Gao, Y.; Liu, S.; Yang, H.; Zhang, L. Saponins from stems and leaves of Panax ginseng prevent obesity via regulating thermogenesis, lipogenesis and lipolysis in high-fat diet-induced obese C57BL/6 mice. Food Chem. Toxicol. 2017, 106, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Hu, X.; Zhang, T.; Dong, P.; Li, Z.; Xue, C.; Chang, Y.; Wang, Y. Saponin from sea cucumber exhibited more significant effects than ginsenoside on ameliorating high fat diet-induced obesity in C57BL/6 mice. MedChemComm 2018, 9, 725–734. [Google Scholar] [CrossRef]
- Song, M.Y.; Kim, B.S.; Kim, H. Influence of Panax ginseng on obesity and gut microbiota in obese middle-aged Korean women. J. Ginseng Res. 2014, 38, 106–115. [Google Scholar] [CrossRef]
- Liu, R.; Zheng, Y.; Cai, Z.; Xu, B. Saponins and flavonoids from adzuki bean (Vigna angularis L.) ameliorate high-fat diet-induced obesity in ICR mice. Front. Pharmacol. 2017, 8, 687. [Google Scholar] [CrossRef]
- Abd Allah, A.L.; Abd-Elrahman, W.M. Hypocholesterolemic and anti-obesity effects of radish sprouts (Raphanus sativus) in adult females. Egypt. J. Food Sci. 2021, 49, 19–34. [Google Scholar] [CrossRef]
- Thakur, A.; Sharma, R. Health promoting phytochemicals in vegetables: A mini review. Int. J. Food Ferment. Technol. 2018, 8, 107–117. [Google Scholar] [CrossRef]
- Ngamsamer, C.; Sirivarasai, J.; Sutjarit, N. The benefits of anthocyanins against obesity-induced inflammation. Biomolecules 2022, 12, 852. [Google Scholar] [CrossRef]
- Badshah, H.; Ullah, I.; Kim, S.E.; Kim, T.H.; Lee, H.Y.; Kim, M.O. Anthocyanins attenuate body weight gain via modulating neuropeptide Y and GABAB1 receptor in rats hypothalamus. Neuropeptides 2013, 47, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Bakuradze, T.; Tausend, A.; Galan, J.; Groh, I.A.M.; Berry, D.; Tur, J.A.; Marko, D.; Richling, E. Antioxidative activity and health benefits of anthocyanin-rich fruit juice in healthy volunteers. Free. Radic. Res. 2019, 53, 1045–1055. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Xia, M.; Zou, T.; Ling, W.; Zhong, R.; Zhang, W. Cyanidin 3-glucoside attenuates obesity-associated insulin resistance and hepatic steatosis in high-fat diet-fed and db/db mice via the transcription factor FoxO1. J. Nutr. Biochem. 2012, 23, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Dey, P. Gut Microbiota in Phytopharmacology: A Comprehensive Overview of Concepts, Reciprocal Interactions, Biotransformations and Mode of Actions. Pharmacol. Res. 2019, 147, 104367. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Choi, M.; Lee, M. Effects of anthocyanin supplementation on reduction of obesity criteria: A systematic review and meta-analysis of randomized controlled trials. Nutrients 2021, 13, 2121. [Google Scholar] [CrossRef]
- Park, S.; Kang, S.; Jeong, D.Y.; Jeong, S.Y.; Park, J.J.; Yun, H.S. Cyanidin and malvidin in aqueous extracts of black carrots fermented with Aspergillus oryzae prevent the impairment of energy, lipid and glucose metabolism in estrogen-deficient rats by AMPK activation. Genes Nutr. 2015, 10, 6. [Google Scholar] [CrossRef]
- Liu, J.; Peng, Y.; Yue, Y.; Shen, P.; Park, Y. Epigallocatechin-3-gallate reduces fat accumulation in Caenorhabditis elegans. Prev. Nutr. Food Sci. 2018, 23, 214–219. [Google Scholar] [CrossRef]
- Peng, C.H.; Cheng, J.J.; Yu, M.H.; Chung, D.J.; Huang, C.N.; Wang, C.J. Solanum nigrum polyphenols reduce body weight and body fat by affecting adipocyte and lipid metabolism. Food Funct. 2020, 11, 483–492. [Google Scholar] [CrossRef]
- Roberts, A.T. Gallic Acid: Inhibiting Angiogenesis in Adipose Tissue; Louisiana State University and Agricultural & Mechanical College: Baton Rouge, LA, USA, 2006. [Google Scholar]
- Guo, X.; Zhang, T.; Shi, L.; Gong, M.; Jin, J.; Zhang, Y.; Liu, R.; Chang, M.; Jin, Q.; Wang, X. The relationship between lipid phytochemicals, obesity and its related chronic diseases. Food Funct. 2018, 9, 6048–6062. [Google Scholar] [CrossRef]
- Lai, C.S.; Wu, J.C.; Pan, M.H. Molecular mechanism on functional food bioactives for anti-obesity. Curr. Opin. Food Sci. 2015, 2, 9–13. [Google Scholar] [CrossRef]
- Trigueros, L.; Peña, S.; Ugidos, A.V.; Sayas-Barberá, E.; Pérez-Álvarez, J.A.; Sendra, E. Food ingredients as anti-obesity agents: A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Carnauba, R.A.; Chaves, D.F.; Baptistella, A.B.; Paschoal, V.; Naves, A.; Buehler, A.M. Association between high consumption of phytochemical-rich foods and anthropometric measures: A systematic review. Int. J. Food Sci. Nutr. 2017, 68, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Abshirini, M.; Mahaki, B.; Bagheri, F.; Siassi, F.; Koohdani, F.; Sotoudeh, G. Higher intake of phytochemical-rich foods is inversely related to prediabetes: A case-control study. Int. J. Prev. Med. 2018, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Mollica, A.; Zengin, G.; Stefanucci, A.; Ferrante, C.; Menghini, L.; Orlando, G.; Brunetti, L.; Locatelli, M.; Dimmita, M.P.; Novellino, E.; et al. Nutraceutical potential of Corylus avellana daily supplements for obesity and related dysmetabolism. J. Funct. Foods 2018, 47, 562–574. [Google Scholar] [CrossRef]
- Samad, A.; Azlan, A.; Ismail, A. Therapeutic effects of vinegar: A review. Curr. Opin. Food Sci. 2016, 8, 56–61. [Google Scholar] [CrossRef]
- Kim, A.Y.; Jeong, Y.J.; Park, Y.B.; Lee, M.K.; Jeon, S.M.; McGregor, R.A.; Choi, M.S. Dose dependent effects of lycopene enriched tomato-wine on liver and adipose tissue in high-fat diet fed rats. Food Chem. 2012, 130, 42–48. [Google Scholar] [CrossRef]
- Benbouguerra, N.; Hornedo-Ortega, R.; Garcia, F.; El Khawand, T.; Saucier, C.; Richard, T. Stilbenes in grape berries and wine and their potential role as anti-obesity agents: A review. Trends Food Sci. Technol. 2021, 112, 362–381. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Kim, S.H.; Kim, D.S.; Park, S.H.; Yoo, B.W.; Kim, H.K. Nutritional composition and anti-obesity effects of cereal bar containing Allium fistulosum (welsh onion) extract. J. Funct. Foods 2014, 6, 428–437. [Google Scholar] [CrossRef]
- Aljutaily, T.; Elbeltagy, A.; Ali, A.A.; Gadallah, M.G.; Khalil, N.A. Anti-Obesity Effects of Formulated Biscuits Supplemented with Date’s Fiber; Agro-Waste Products Used as a Potent Functional Food. Nutrients 2022, 14, 5315. [Google Scholar] [CrossRef]
- Gorjanović, S.; Micić, D.; Pastor, F.; Tosti, T.; Kalušević, A.; Ristić, S.; Zlatanović, S. Evaluation of apple pomace flour obtained industrially by dehydration as a source of biomolecules with antioxidant, antidiabetic and antiobesity effects. Antioxidants 2020, 9, 413. [Google Scholar] [CrossRef]
- Cho, Y.J.; Lee, H.G.; Seo, K.H.; Yokoyama, W.; Kim, H. Antiobesity effect of prebiotic polyphenol-rich grape seed flour supplemented with probiotic kefir-derived lactic acid bacteria. J. Agric. Food Chem. 2018, 66, 12498–12511. [Google Scholar] [CrossRef]
- Zhou, F.; Yin, M.; Liu, Y.; Han, X.; Guo, J.; Ren, C.; Wang, W.; Huang, W.; Zhan, J.; You, Y. Grape seed flour intake decreases adiposity gain in high-fat-diet induced obese mice by activating thermogenesis. J. Funct. Foods 2019, 62, 103509. [Google Scholar] [CrossRef]
- Lim, W.C.; Ho, J.N.; Lee, H.S.; Cho, H.Y. Germinated waxy black rice extract inhibits lipid accumulation with regulation of multiple gene expression in 3T3-L1 adipocytes. Food Sci. Biotechnol. 2016, 25, 821–827. [Google Scholar] [CrossRef]
- Koh, S.P.; Sew, Y.S.; Sabidi, S.; Maarof, S.; Sharifudin, S.A.; Abdullah, R. Anti-obesity Effects of SCOBY Jackfruit Beverages and Their Influence on Gut Microbiota. Explor. Res. Hypothesis Med. 2023, 8, 14–24. [Google Scholar] [CrossRef]
- Wu, T.; Jiang, Z.; Yin, J.; Long, H.; Zheng, X. Anti-obesity effects of artificial planting blueberry (Vaccinium ashei) anthocyanin in high-fat diet-treated mice. Int. J. Food Sci. Nutr. 2016, 67, 257–264. [Google Scholar] [CrossRef]
- Ali, Z.; Ma, H.; Rashid, M.T.; Ayim, I.; Wali, A. Reduction of body weight, body fat mass, and serum leptin levels by addition of new beverage in normal diet of obese subjects. J. Food Biochem. 2018, 42, e12554. [Google Scholar] [CrossRef]
- Sheng, Z.; Yu, L.; Li, X.; Zhao, Y.; Dai, W.; Chang, S.K.; Liu, J. The anti-obesity effect of fermented tremella/blueberry and its potential mechanisms in metabolically healthy obese rats. J. Funct. Foods 2021, 86, 104670. [Google Scholar] [CrossRef]
- Beh, B.K.; Mohamad, N.E.; Yeap, S.K.; Ky, H.; Boo, S.Y.; Chua, J.Y.H.; Tan, S.W.; Ho, W.Y.; Sharifuddin, S.A.; Long, K.; et al. Anti-obesity and anti-inflammatory effects of synthetic acetic acid vinegar and Nipa vinegar on high-fat-diet-induced obese mice. Sci. Rep. 2017, 7, 6664. [Google Scholar] [CrossRef]
- Saravanan, G.; Ponmurugan, P.; Deepa, M.A.; Senthilkumar, B. Anti-obesity action of gingerol: Effect on lipid profile, insulin, leptin, amylase and lipase in male obese rats induced by a high-fat diet. J. Sci. Food Agric. 2014, 94, 2972–2977. [Google Scholar] [CrossRef]
- Zhou, X.; Pak, S.; Li, D.; Dong, L.; Chen, F.; Hu, X.; Ma, L. Bamboo Shoots Modulate Gut Microbiota, Eliminate Obesity in High-Fat-Diet-Fed Mice and Improve Lipid Metabolism. Foods 2023, 12, 1380. [Google Scholar] [CrossRef]
- Li, X.; Tian, S.; Wang, Y.; Liu, J.; Wang, J.; Lu, Y. Broccoli microgreens juice reduces body weight by enhancing insulin sensitivity and modulating gut microbiota in high-fat diet-induced C57BL/6J obese mice. Eur. J. Nutr. 2021, 60, 3829–3839. [Google Scholar] [CrossRef]
- Kuesten, C.; Dang, J.; Nakagawa, M.; Bi, J.; Meiselman, H.L. Japanese consumer segmentation based on general self-efficacy psychographics data collected in a phytonutrient supplement study: Influence on health behaviors, well-being, product involvement and liking. Food Qual. Prefer. 2022, 99, 104545. [Google Scholar] [CrossRef]
- Liudvytska, O.; Kolodziejczyk-Czepas, J. A review on rhubarb-derived substances as modulators of cardiovascular risk factors—A special emphasis on anti-obesity action. Nutrients 2022, 14, 2053. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Hung, H.Y. Recent advances in natural anti-obesity compounds and derivatives based on in vivo evidence: A mini-review. Eur. J. Med. Chem. 2022, 237, 114405. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Singh, D.D.; Lakhawat, S.S.; Yasmeen, N.; Pandey, A.; Singla, R.K. Biogenic phytochemicals modulating obesity: From molecular mechanism to preventive and therapeutic approaches. Evid. Based Complement. Altern. Med. 2022, 2022, 68522. [Google Scholar] [CrossRef]
- Kazemipoor, M.; Cordell, G.A.; Sarker, M.M.R.; Radzi, C.W.J.B.W.M.; Hajifaraji, M.; En Kiat, P. Alternative treatments for weight loss: Safety/risks and effectiveness of anti-obesity medicinal plants. Int. J. Food Prop. 2015, 18, 1942–1963. [Google Scholar] [CrossRef]
- Sun, N.N.; Wu, T.Y.; Chau, C.F. Natural dietary and herbal products in anti-obesity treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef]
- Patil, B.S.; Uckoo, R.M.; Jayaprakasha, G.K.; Palma, M.A. Consumers’ changing perceptions of quality: Revisiting the science of fruit and vegetable cultivation for improved health benefits. In XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes (IHC2014); International Society for Horticultural Science: Leuven, Belgium, 2016; Volume 1120, pp. 459–468. [Google Scholar] [CrossRef]
- Paraschivu, M.; Cotuna, O.; Sărățeanu, V.; Durău, C.C.; Păunescu, R.A. Microgreens-current status, global market trends and forward statements. Sci. Pap. Ser. Manag. Econ. Eng. Agric. Rural. Dev. 2021, 21, 633–639. [Google Scholar]
- Viganó, J.; Zabot, G.L.; Martínez, J. Supercritical fluid and pressurized liquid extractions of phytonutrients from passion fruit by-products: Economic evaluation of sequential multi-stage and single-stage processes. J. Supercrit. Fluids 2017, 122, 88–98. [Google Scholar] [CrossRef]
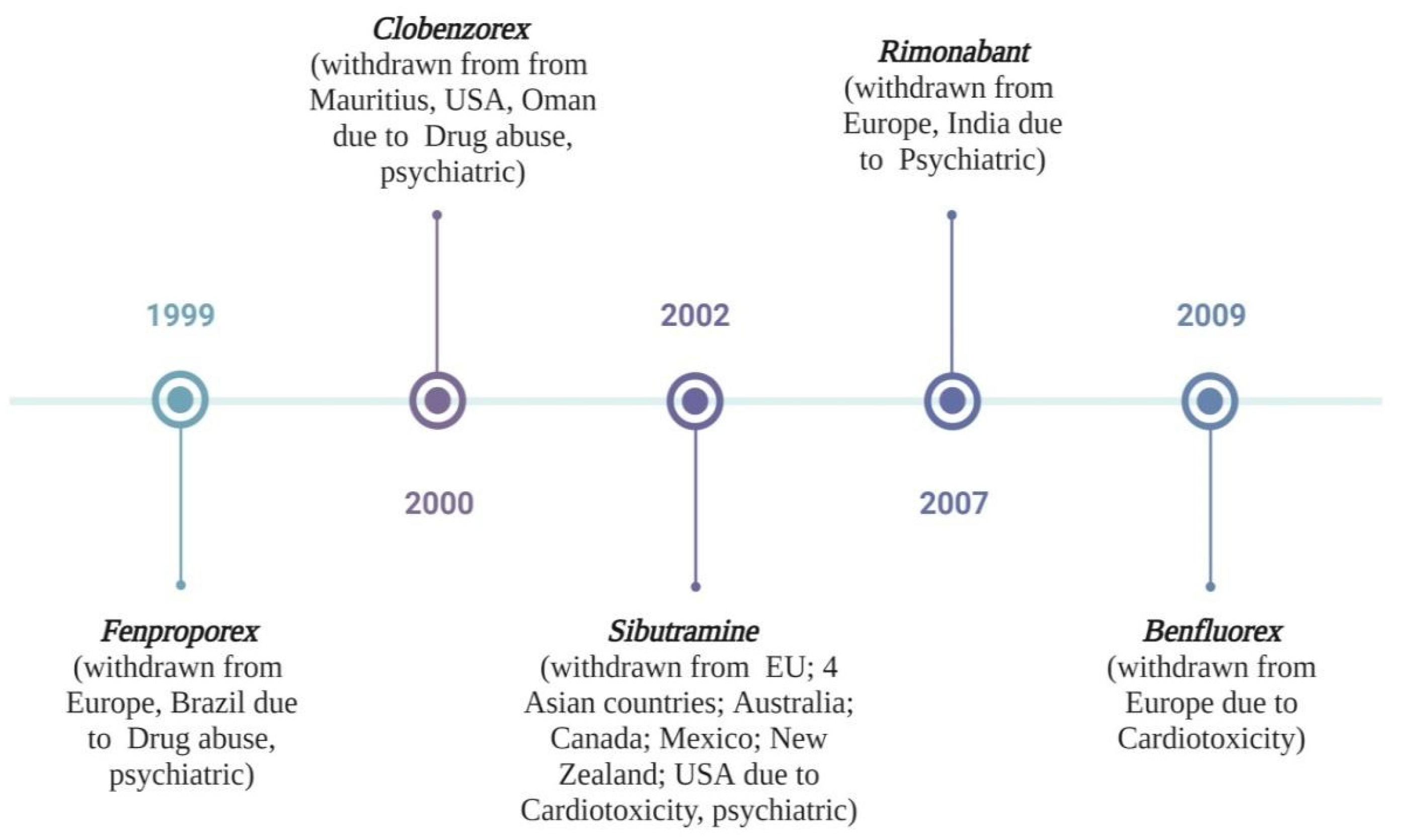
- Müller, T.D.; Blüher, M.; Tschöp, M.H.; DiMarchi, R.D. Anti-obesity drug discovery: Advances and challenges. Nat. Rev. Drug Discov. 2022, 21, 201–223. [Google Scholar] [CrossRef]
- Sawant, O.; Khan, T. Standardization of marketed anti-obesity nutraceuticals containing amla and ginseng. J. Food Process. Preserv. 2021, 45, e15693. [Google Scholar] [CrossRef]
- Bessesen, D.H.; Van Gaal, L.F. Progress and challenges in anti-obesity pharmacotherapy. Lancet Diabetes Endocrinol. 2018, 6, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Koncz, D.; Tóth, B.; Roza, O.; Csupor, D. A systematic review of the European rapid alert system for food and feed: Tendencies in illegal food supplements for weight loss. Front. Pharmacol. 2021, 11, 611361. [Google Scholar] [CrossRef] [PubMed]
- Dastjerdi, A.G.; Akhgari, M.; Kamali, A.; Mousavi, Z. Principal component analysis of synthetic adulterants in herbal supplements advertised as weight loss drugs. Complement. Ther. Clin. Pract. 2018, 31, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.K.; Kumar, B.K.; Sharma, P.K.; Murugesan, S.; Deepa, P.R. In silico and in vitro analysis of PPAR–α/γ dual agonists: Comparative evaluation of potential phytochemicals with anti-obesity drug orlistat. Comput. Biol. Med. 2022, 147, 105796. [Google Scholar] [CrossRef]
- Castrica, M.; Rebucci, R.; Giromini, C.; Tretola, M.; Cattaneo, D.; Baldi, A. Total phenolic content and antioxidant capacity of agri-food waste and by-products. Ital. J. Anim. Sci. 2019, 18, 336–341. [Google Scholar] [CrossRef]
- Ben-Othman, S.; Jõudu, I.; Bhat, R. Bioactives from agri-food wastes: Present insights and future challenges. Molecules 2020, 25, 510. [Google Scholar] [CrossRef]


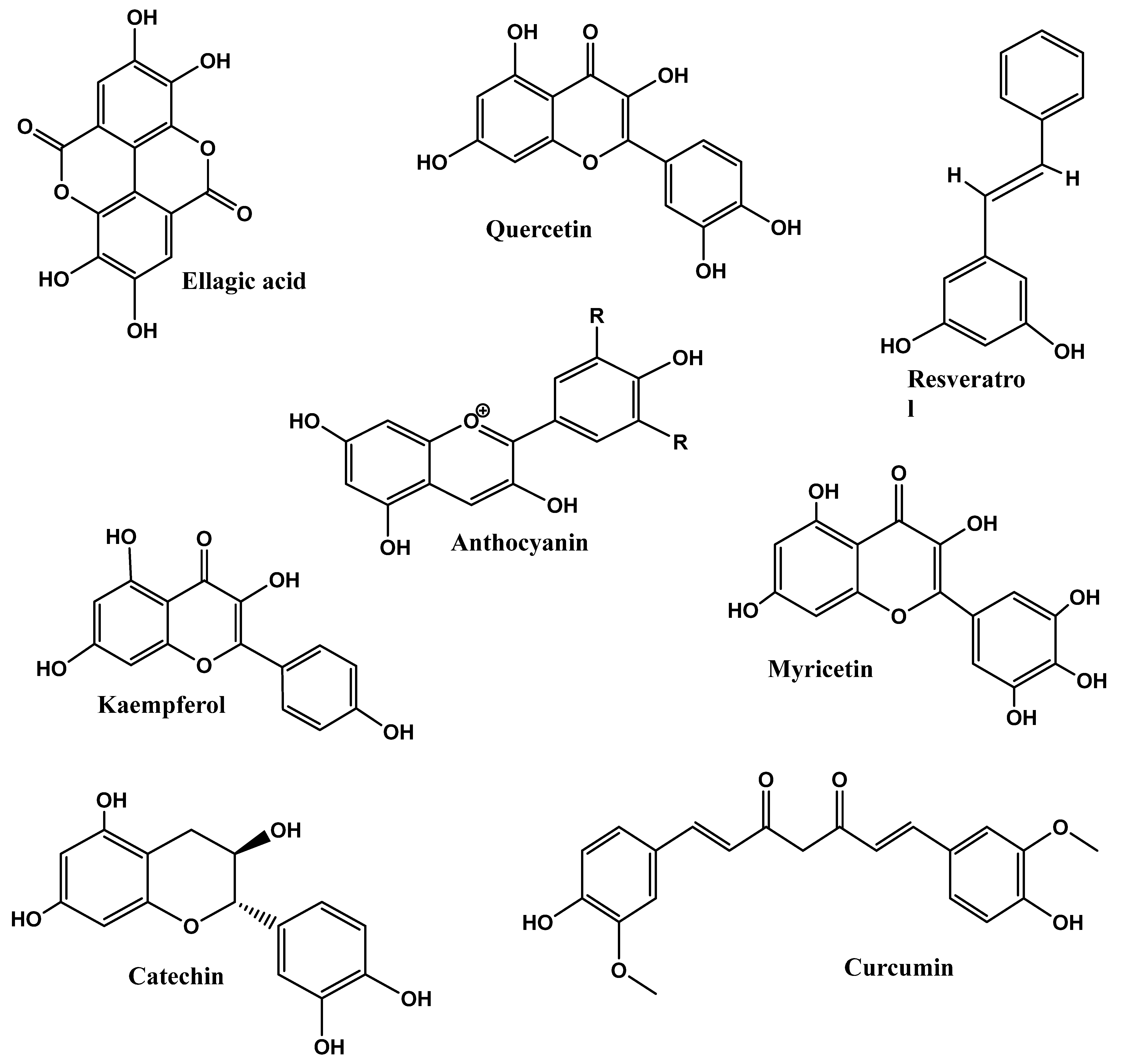
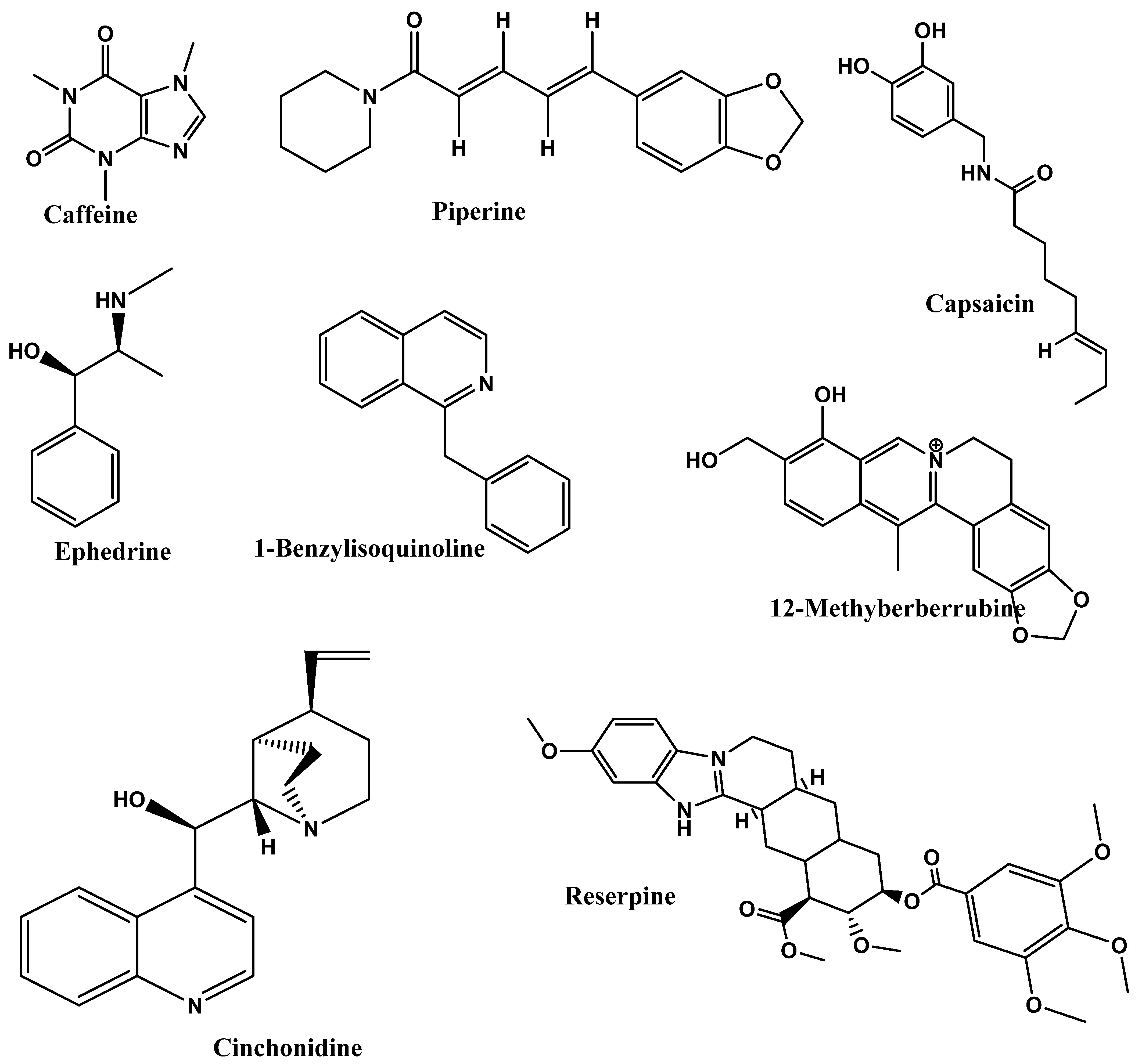
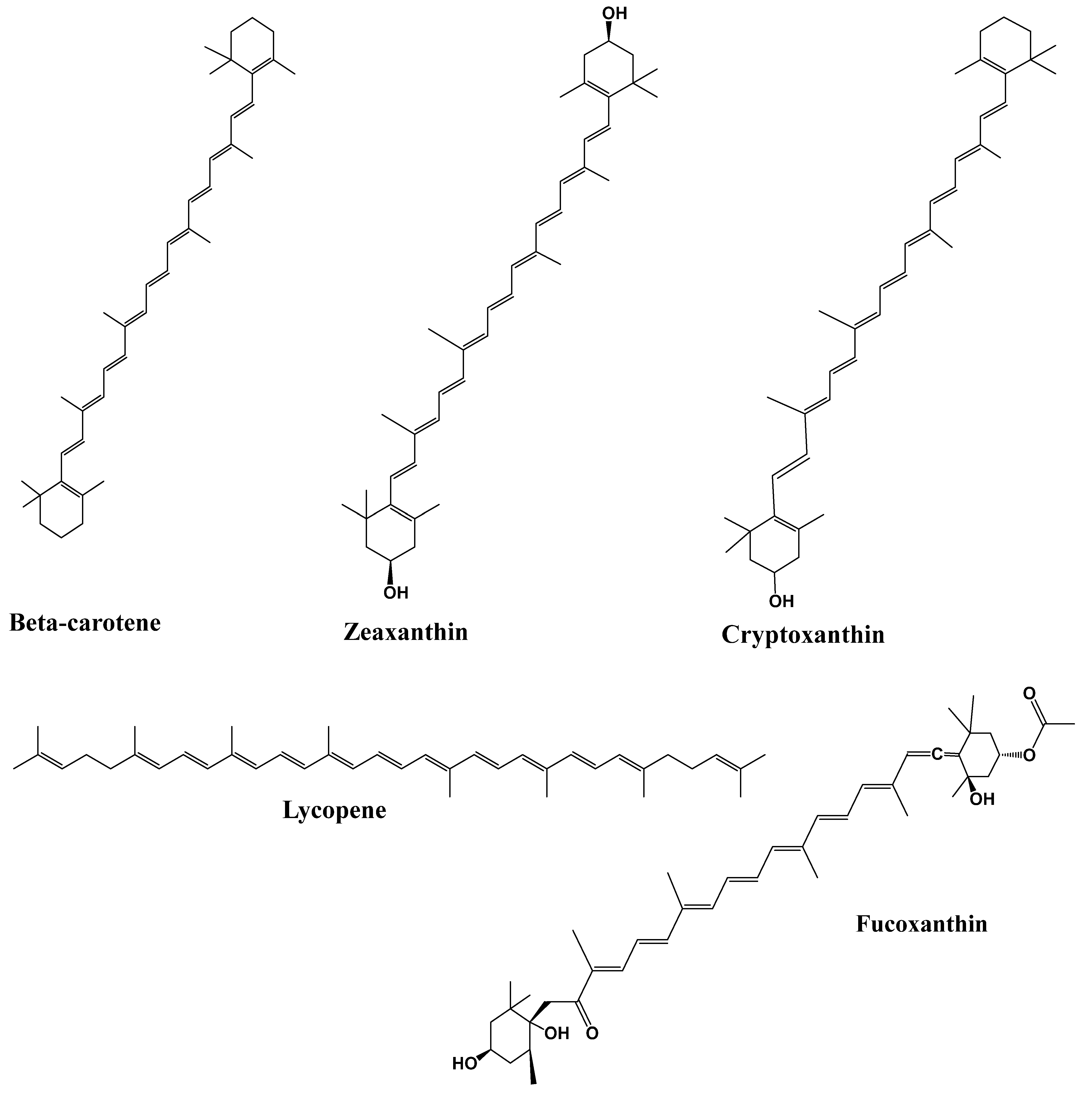
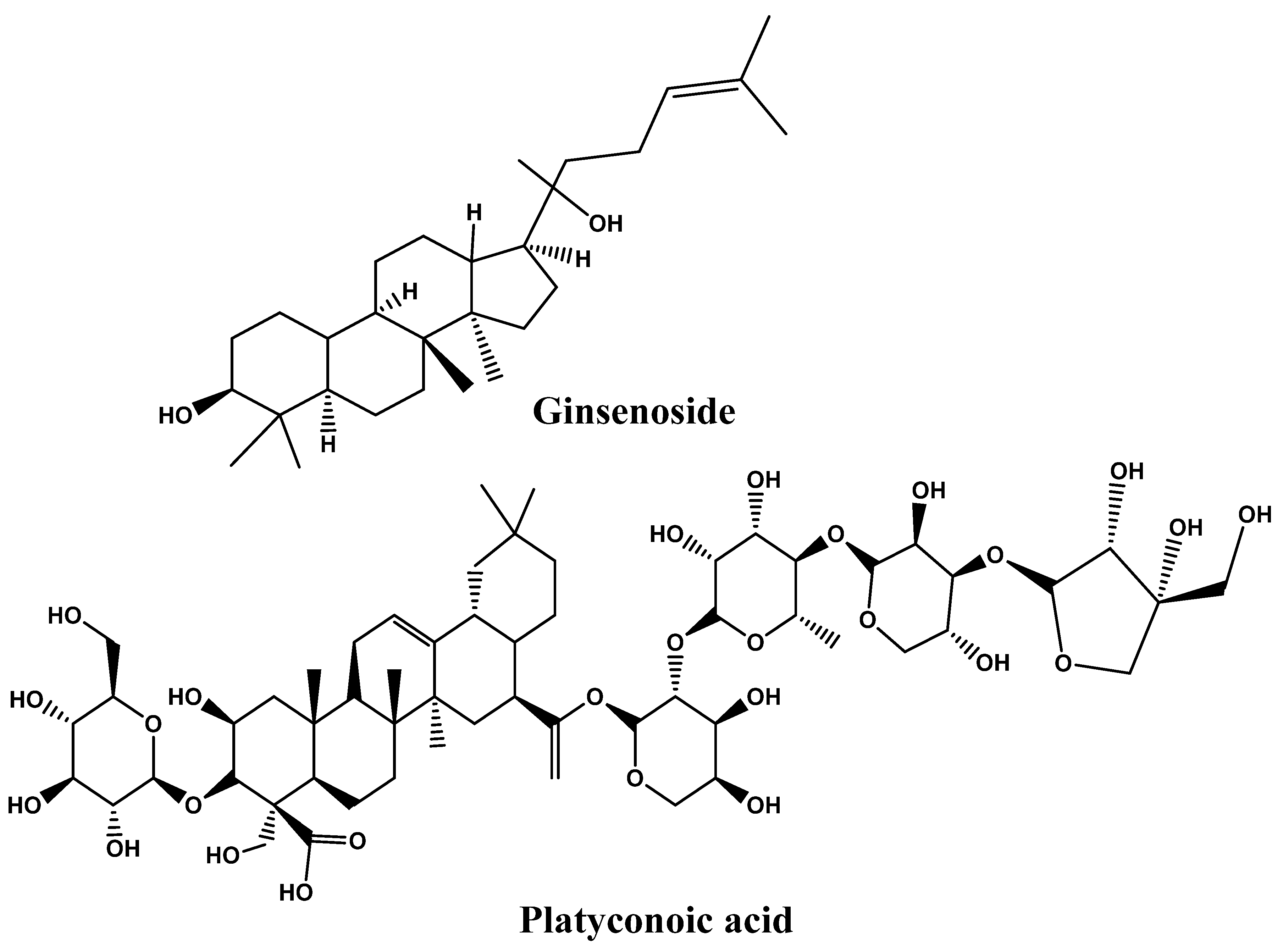
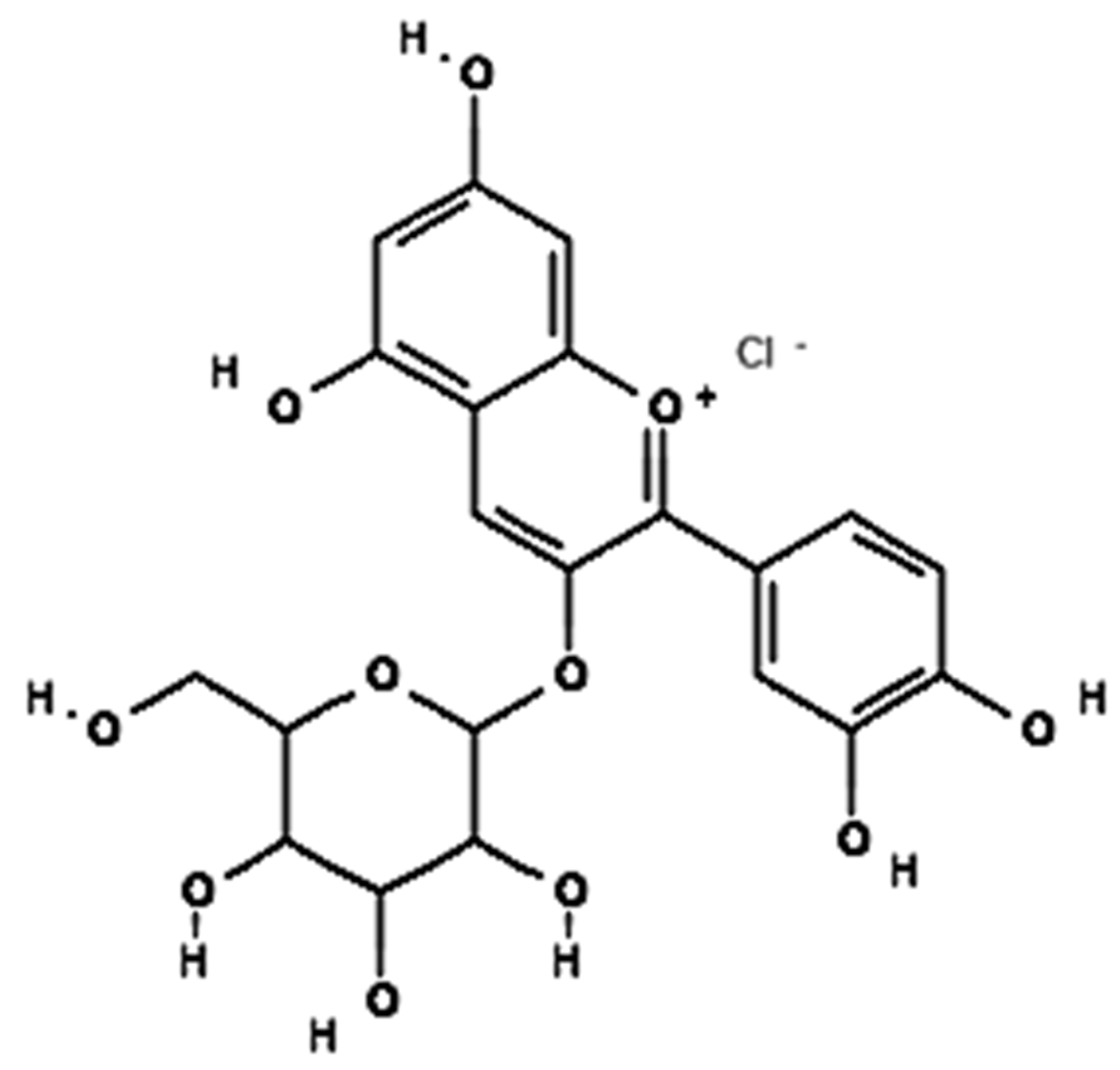

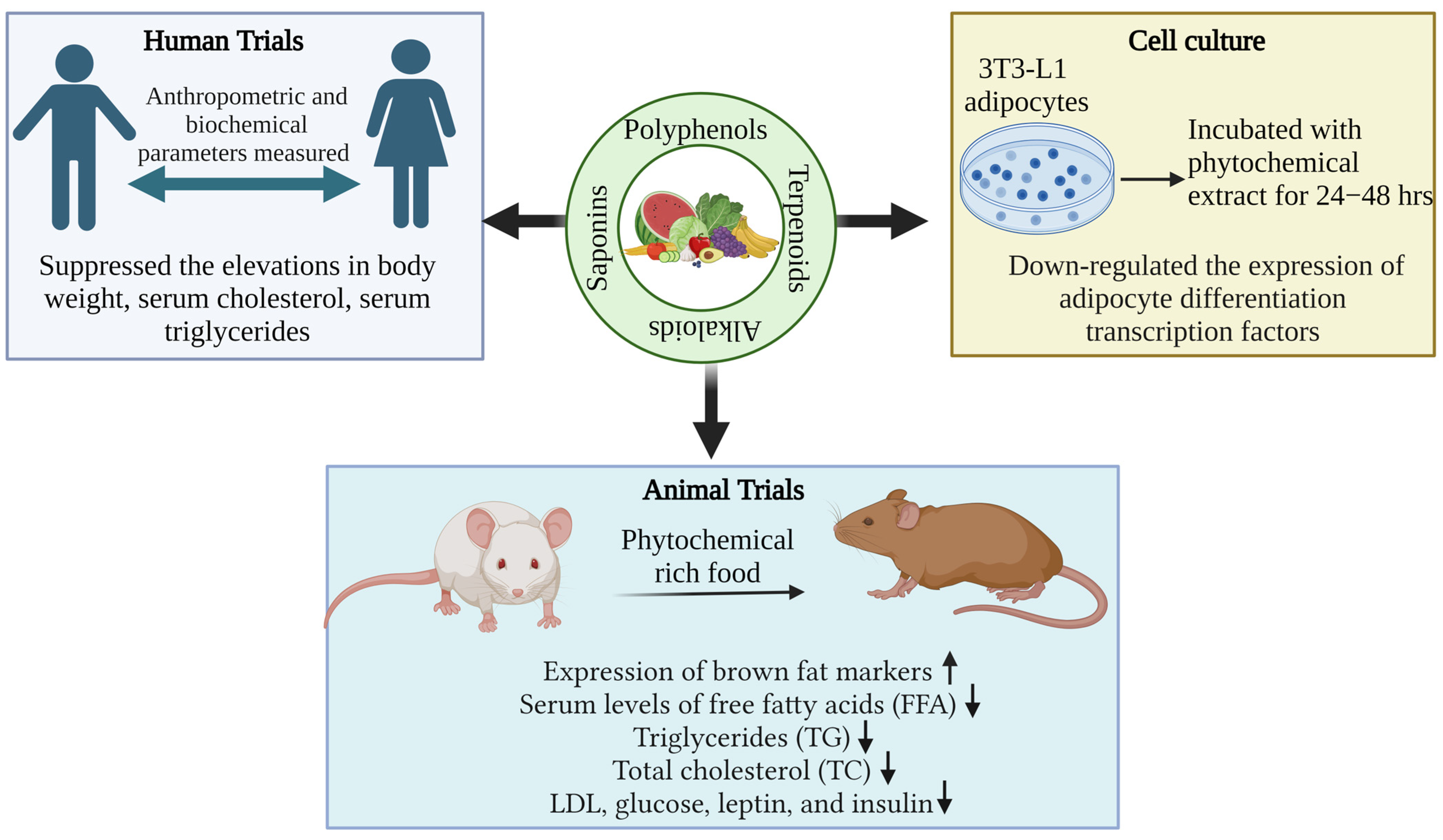
| Phytonutrients/ Source | Experimental Model/Condition | Experimental Design/Trial | Main Outcomes | References |
|---|---|---|---|---|
| Red ginseng (saponin fraction) | 3T3-L1 and RAW264.7 cells | Cells (1 × 104 or 1 × 105 cells/well) were incubated in 96-well plate for 24 h, sample (SF) (100 μg/mL) pretreated for 2 h and incubated for 24 h or 48 h; medium was removed and MTT reagent was added for 60 min at 37 °C followed by Dimethyl sulfoxide (DMSO) was added; and absorbance was recorded at 550 nm | SF suppressed tumor necrosis factor-alpha (TNF-α) (78%), monocyte chemo attractant protein-1 (MCP-1) (40%), and interleukin-6 (IL-6) (22%). Increases nuclear factor erythroid-derived 2(Nrf2) and target protein, hemoxygenase-1 (HO-1). | [195] |
| 11 protoberberine-type alkaloids (obtained from Berberis (Berberidaccae) and Coptis rhizomes) | 3T3-L1 adipocytes | Alkaloids (5µM) treated on adipocytes for 4 days, followed by adipocyte staining with Oil Red O on Day 12 | 13-Methylberberine down-regulated the expression of adipocyte differentiation transcription factors, (PPARγ) and CCAAT (C/EBPα). | [180] |
| Zeaxanthin | 3T3-L1 adipocytes | Cells were seeded in 96-well plates at 1 × 103 densityandwere cultured for 12 h in serum-free DMEM, various concentrations of zeaxanthin (0–60 µM) were incubated for 72 h;cell toxic effects of zeaxanthin were evaluated | Zeaxanthin significantly decreases the lipid content from intracellular in dose-dependent manner (5–15 µM) in adipocytes without causing cytotoxicity. | [190] |
| Green tea | High-fat diet-fed rats | 400 to 800 mg/kg dose was given for 6 weeks | Low serum leptin levels in rats; Inhibits fatty acid absorption; Suppresses expression of both IL-6 as well as TNF-α gene. | [156] |
| Adzuki beans (Vigna angularis L.) flavonoids and saponins | ABTE (Adzuki bean total extract), ABF (Adzuki bean flavonoid), and ABS (Adzuki bean saponin) orally administered in HFD mice | 60 and 300 mg/kg per day ABTE, ABF, and ABS for 4 weeks | Enhance lipolysis; reduce final body weight and adipose tissue accumulation. Reduce total cholesterol, serum triglyceride levels, LDL-cholesterol, and liver lipid. | [199] |
| Flavonol kaempferol | Male mice C57BL/6J | HFD+kaempferol @ 0.01 or 0.05% for 5 months | Increased lipolysis. Prevents high fatty acid-impaired glucose uptake, glycogen synthesis, AMPK activity, and Glut4 expression in skeletal muscle cells. Improving peripheral insulin sensitivity and protecting against pancreatic β-cell dysfunction. | [153] |
| Muscadine grape or wine | Male C57BL/6J mice |
| Decreased body weight by 12% compared to HF controls. Reduced plasma content of triglycerides, free fatty acids, and cholesterol in obese mice. | [154] |
| Sprouted Red Radish seed | Low-calorie diet with sprouted red radish seed (100 g per day) in adult female (25–40 years old) for 8 weeks | Anthropometric measurements and blood samples for analysis | Suppressed the elevations in body weight (~12%), serum cholesterol (~27.5% reduction), serum triglycerides (~33% reduction), and glucose (~7% reduction). | [200] |
| Tomato | 35 Caucasian women aged between 18 and 25 years consume tomato (cv. rama) @ average of 90 g/per day before lunch for 4 weeks | Anthropometric and biochemical parameters were measured | Reduced body weight, blood glucose, % fat, cholesterol, fasting triglycerides, and uric acid. | [191] |
| Phytonutrient Compounds | Mechanism System Target | Experimental Model | Major Findings | References |
|---|---|---|---|---|
| Phlorizin (PHZ) | In vitro study: Block the metabolic;LDL oxidation was prevented. | LDL-C isolated from human plasma | LDL-cholesterol oxidation was blocked | [158,159] |
| In vivo study: Improved manufacturing of the hormone glucagon-like peptide-2 (GLP-2) and healing of gut epithelial barriers disruption brought on by HFD. Clinical trial:Not yet | High-fat diet (HFD) induced gut microbiota alterations | PHZ’s anti-obesity action may also be mediated through the gut microbiota–barrier pathway. | ||
| Epigallocatechin gallate | In vitro study: Reduce adipogenesis to lower C. elegans fat content, as indicated by the reduced ATGL-1 gene expression level following EGCG therapy. | C. elegans strains and Escherichia coliOP50diet | Controls the body fat content | [163,165,209] |
| In vivo study: Increasing thermogenesis to improve utilization of energy. | Male C57BL/6J mice induced obesity | The blood sugar and triglyceride concentrations were significantly decreased, lowering lipid formation in adipose tissues and affecting weight growth. | ||
| Clinical trial: Controlled the plasma cholesterol and triglycerides | 15 women with central obesity were screened at our clinic. A total of 102 of them with a body mass index (BMI) ≥ 27 kg/m2 and a waist circumference (WC) ≥ 80 cm | Decrease body weight and BMI in obese women after a 12-week treatment, significantly decrease waist circumference. | ||
| Anthocyanins | In vitro study: Anthocyanins reduce the complete downstream cascade of pro-inflammatory mediators, including C-reactive protein (CRP), interleukin (IL)-6, and tumor necrosis factor (TNF), and they also ameliorate gut dysbiosis, therefore, reestablishing a healthy gut microbiota. | IL-6 gene in lipopolysaccharide (LPS)-induced adipose stem cells | Treat obesity-related inflammation and chronic diseases. | [202,203,204] |
| In vivo study: Regulating neuropeptide Y, and the -aminobutyric acid receptor in the hypothalamus decreases appetite | Male Sprague–Dawley rats induced obesity | In comparison to the control, there was a substantial decline in body weight growth (15.76%) and regular caloric consumption (19.10%) | ||
| Clinical trial: Lowered the incidence of metabolic disorders and decreased lipids, body composition, and inflammation in obese people | Healthy male volunteers | Reduce body mass index, low-density lipoprotein cholesterol | ||
| Resveratrol and Quercetin | In vitro study: Lower the gene expression of the essential adipogenic factors peroxisome proliferator-activated receptor (PPAR) and CCAAT/enhancer binding protein (C/EBP) to suppress adipogenesis. | Human SGBS adipocytes | Reduced concentrations of the adipokines ANGPTL4, adipsin, and PAI-1 as well as the glycolysis-related enzymes ENO2, PFKP, and PFKFB4; all arelinked to obesity and malfunction of adipose tissue. | [173,174] |
| In vivo study: Alteration of the intestinal flora | Male Wistar rats induced obesity | Reductions in adipocyte size, visceral adipose tissue weight, and body weight increase that are substantial | ||
| Clinical Trial: The inhibition of genes related to angiogenesis, Wnt signaling, intercellular connection, G protein-coupled receptors, and Notch signaling mechanisms involved in cell cycle regulation that have been up-regulated | 11 obese otherwise healthy men | Adipogenesis increased and adipocyte size was decreased. | ||
| Gallic acid | In vitro study: The size of adipocytes effected due adipose tissue inflammation and metabolic dysfunction, which are linked to adipocyte hypertrophy | Murine 3T3-L1 preadipocytes and RAW 264 macrophages | A typical fat cell content, the WAT group’s 18 size was noticeably smaller as compared to the GA group; with a control group | [169,210,211] |
| In vivo study: Lipolysis is induced and FAS is suppressed to prevent lipogenesis as a means of controlling the process of lipid metabolism. | Male C57BL/6 mice induced obesity | Weight loss and a decline in the accumulation of fat | ||
| Clinical trial: Suppressing adipogenesis and proliferation and reducing pancreatic lipase activity | Obese human subjects receiving capsules containing 200 mg of gallic acid and 50 mg of a Chinese herbal decoction, three times a day for 24 weeks | Serum levels in humans, which is what prevented it from causing weight reduction or a decrease in food intake. | ||
| Cyanidin | In vitro study: Lower the levels of pro-inflammatory substances correlated with fat, including tumor necrosis factor (TNF), interleukin-6, and C-reactive protein (CRP) | PC12 cells treated with H2O2 | In PC12 cells exposed to H2O2, oxidative stress-associated toxic effects were reduced. | [202,208] |
| In vivo study: Regulates the breakdown of lipids and enhancement of energy consumption | Ovariectomized Female Sprague–Dawley rats induced obesity | Significantly reducing the rate of obesity increase by 32.83%, triglycerides by 24.4%, and LDL by 29.58%, contrasted to control | ||
| Clinical trial: Alterations to lipid metabolism in conjunction with inflammatory indicators | Overweight and obesity (250 mL of blackberry juice) | Reduced levels of malondialdehyde, hydroxynonenal, and serum-oxidized LDL in the plasma and serum |
| Phytonutrients/ Source | Food Application and Purpose | Functional Effect Claim | Major Findings | References |
|---|---|---|---|---|
| Blueberry anthocyanin (BA) | Obese mice were supplemented with BA in daily food at doses of 50, 100, and 200 mg/kg | Anti-obesity effect |
| [228] |
| Allicin (vinyldithinins, sulfides, and ajoene) | Dates fruit and garlic-based beverage | Anti-obesity |
| [229] |
| Total polyphenols and monosaccharides | Tremella and blueberryfermented concentrate | Anti-obesity |
| [230] |
| Nipa vinegar | Nipa vinegar added in mice HFD-fed at 0.08 and 2 mL/kg body weight for 33 weeks | Anti-inflammatory and anti-obesity effect |
| [231] |
| Gingerol from ginger | Supplementation of Gingerol (25–75 mg/kg) suspended in 0.5% carboxy methylcellulose with high-fat diet of mice | Anti-obesity |
| [232] |
| Dietary fiber from bamboo shoot | Bamboo shoots lyophilized powder | Anti-obesity |
| [233] |
| Broccoli microgreens juice | 20 g/kg/body weight broccoli microgreen juice was given in high-fat and high-sugar mice diet for 2 to 10 weeks | Reduce obesity in mice |
| [234] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddiqui, S.A.; Azmy Harahap, I.; Suthar, P.; Wu, Y.S.; Ghosh, N.; Castro-Muñoz, R. A Comprehensive Review of Phytonutrients as a Dietary Therapy for Obesity. Foods 2023, 12, 3610. https://doi.org/10.3390/foods12193610
Siddiqui SA, Azmy Harahap I, Suthar P, Wu YS, Ghosh N, Castro-Muñoz R. A Comprehensive Review of Phytonutrients as a Dietary Therapy for Obesity. Foods. 2023; 12(19):3610. https://doi.org/10.3390/foods12193610
Chicago/Turabian StyleSiddiqui, Shahida Anusha, Iskandar Azmy Harahap, Priyanka Suthar, Yuan Seng Wu, Nibedita Ghosh, and Roberto Castro-Muñoz. 2023. "A Comprehensive Review of Phytonutrients as a Dietary Therapy for Obesity" Foods 12, no. 19: 3610. https://doi.org/10.3390/foods12193610
APA StyleSiddiqui, S. A., Azmy Harahap, I., Suthar, P., Wu, Y. S., Ghosh, N., & Castro-Muñoz, R. (2023). A Comprehensive Review of Phytonutrients as a Dietary Therapy for Obesity. Foods, 12(19), 3610. https://doi.org/10.3390/foods12193610