
Detection of the Microbial Composition of Some Commercial Fermented Liquid Products via Metagenomic Analysis

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. DNA Extraction
2.3. Metagenomic Sequencing
2.4. Statistical Analysis
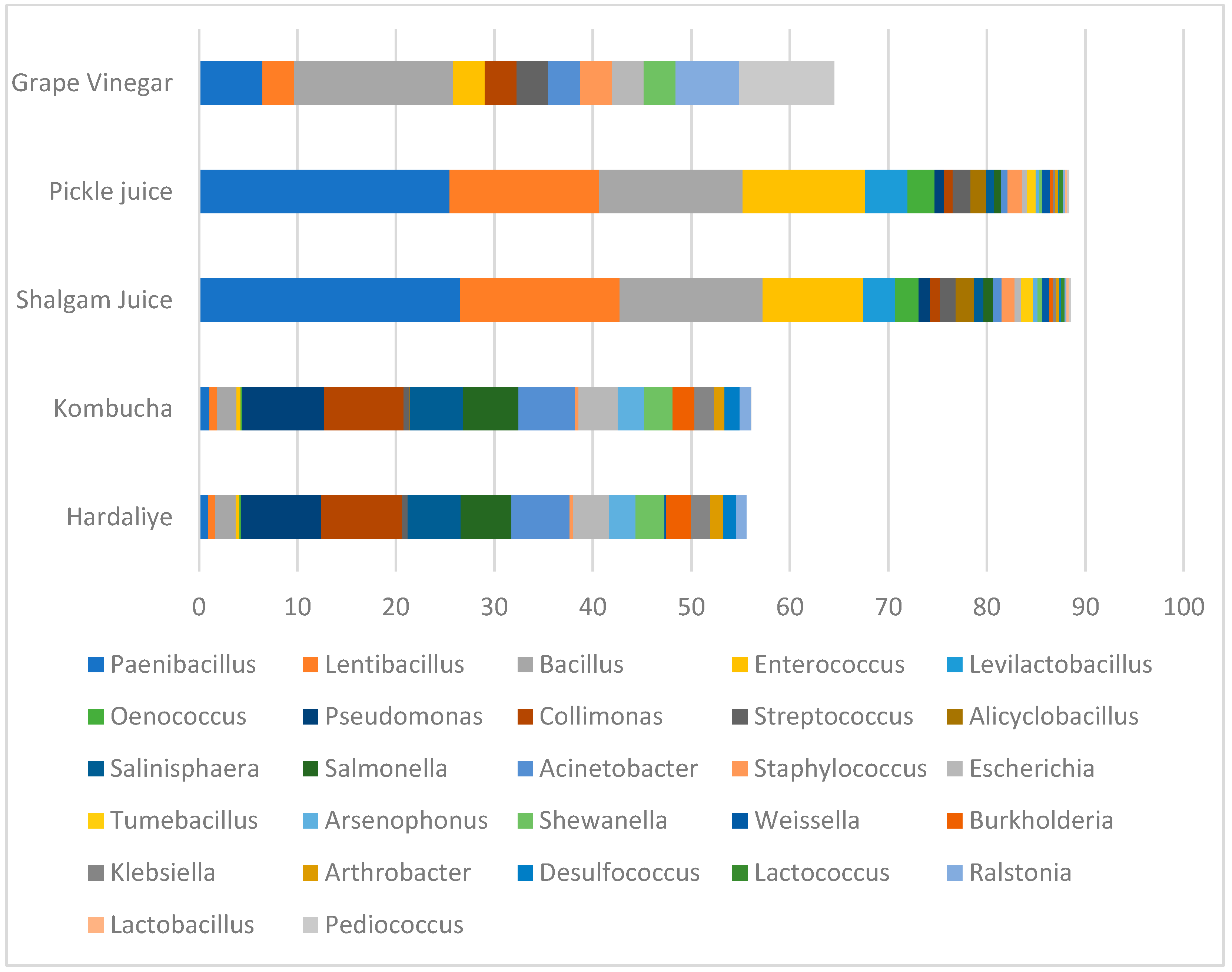
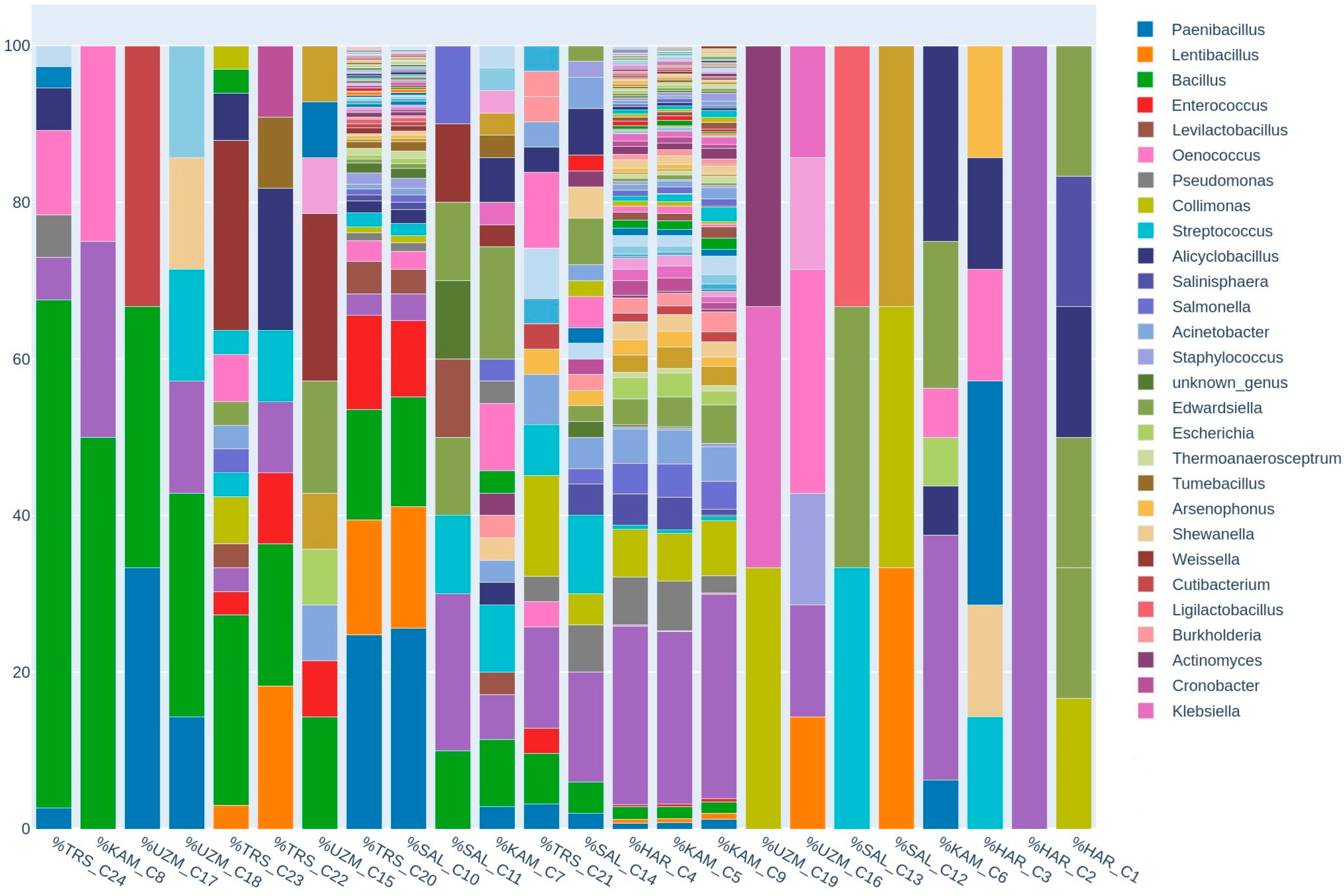
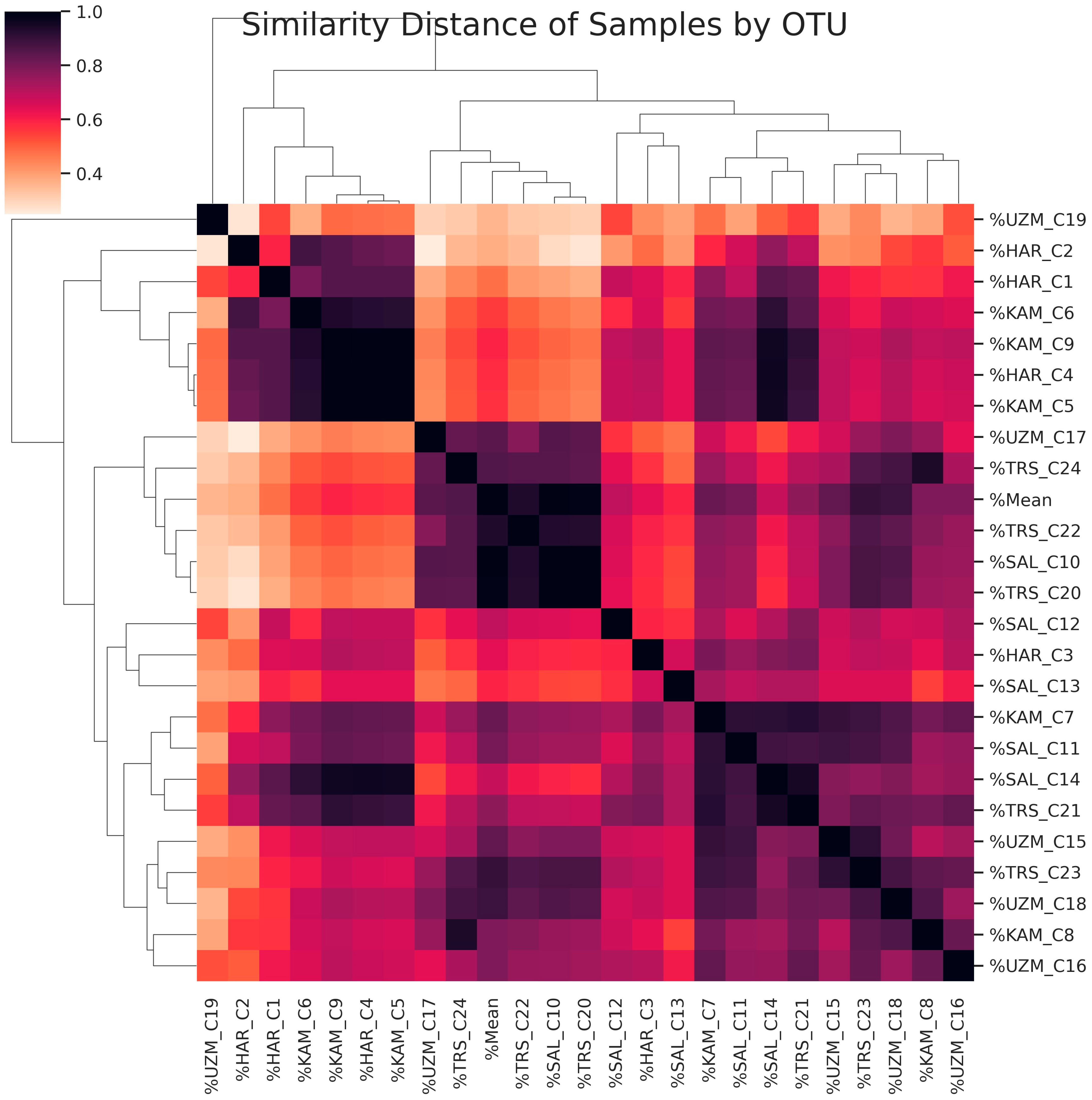
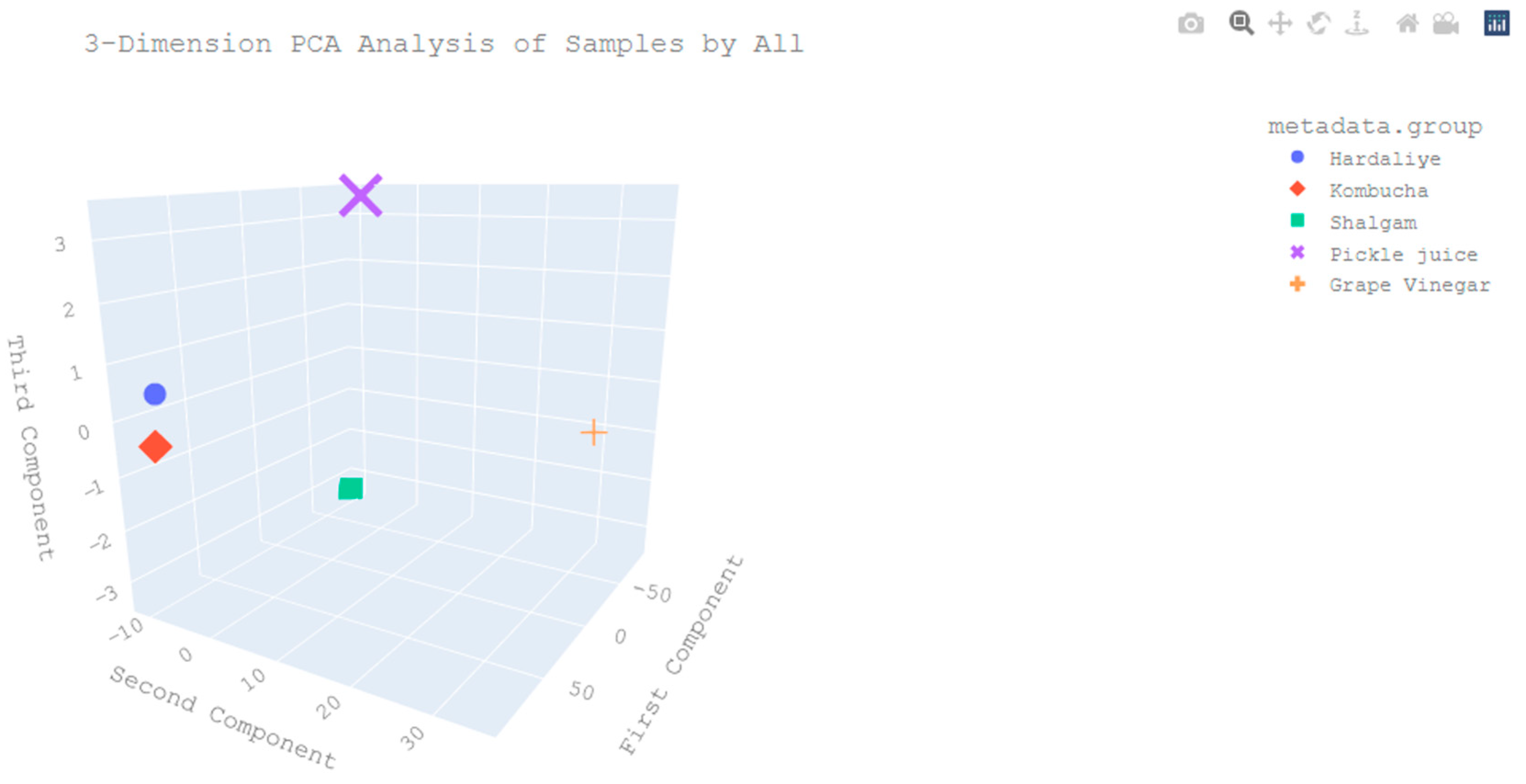
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Marsh, A.J.; O’Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-based analysis of the bacterial and fungal compositions of multiple kombucha (tea fungus) samples. Food Microbiol. 2014, 38, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Mercenier, A.; Pavan, S.; Pot, B. Probiotics as biotherapeutic agents: Present knowledge and future prospects. Curr. Pharm. Des. 2003, 9, 175–191. [Google Scholar] [CrossRef]
- Baschali, A.; Tsakalidou, E.; Kyriacou, A.; Karavasiloglou, N.; Matalas, A.L. Traditional low-alcoholic and non-alcoholic fermented beverages consumed in European countries: A neglected food group. Nutr. Res. Rev. 2017, 30, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Gu, C.T. Enterococcus pingfangensis sp. nov., Enterococcus dongliensis sp. nov., Enterococcus hulanensis sp. nov., Enterococcus nangangensis sp. nov. and Enterococcus songbeiensis sp. nov., isolated from Chinese traditional pickle juice. Int. J. Syst. Evol. 2019, 69, 3191–3201. [Google Scholar] [CrossRef]
- Song, J.; Zhang, J.H.; Kang, S.J.; Zhang, H.Y.; Yuan, J.; Zeng, C.Z.; Huang, Y.L. Analysis of microbial diversity in apple vinegar fermentation process through 16s rdna sequencing. Food Sci. Nutr. 2019, 7, 1230–1238. [Google Scholar] [CrossRef]
- Erten, H.; Tanguler, H.; Canbaş, A. A traditional Turkish lactic acid fermented beverage: Shalgam (Salgam). Food Rev. Int. 2008, 24, 352–359. [Google Scholar] [CrossRef]
- Tanguler, H.; Saris, P.E.; Erten, H. Microbial, chemical and sensory properties of shalgams made using different production methods. J. Sci. Food Agric. 2015, 95, 1008–1015. [Google Scholar] [CrossRef]
- Arici, M.; Coskun, F. Hardaliye: Fermented grape juice as a traditional Turkish beverage. Food Microbiol. 2001, 18, 417–421. [Google Scholar] [CrossRef]
- Neffe-Skocińska, K.; Sionek, B.; Ścibisz, I.; Kołożyn-Krajewska, D. Acid contents and the effect of fermentation condition of Kombucha tea beverages on physicochemical, microbiological and sensory properties. CYTA-J. Food. 2017, 15, 601–607. [Google Scholar] [CrossRef]
- Arıkan, M.; Mitchell, A.L.; Finn, R.D.; Gürel, F. Microbial composition of Kombucha determined using amplicon sequencing and shotgun metagenomics. J. Food Sci. 2020, 85, 455–464. [Google Scholar] [CrossRef]
- Fanning, S.; Proos, S.; Jordan, K.; Srikumar, S. A review on the applications of next generation sequencing technologies as applied to food-related microbiome studies. Front. Microbiol. 2017, 8, 1829. [Google Scholar]
- Li, Y.M.; Cao, W.W.; Liang, S.L.; Yamasaki, S.; Chen, X.; Shi, L.; Ye, L. Metagenomic characterization of bacterial community and antibiotic resistance genes in representative ready-to-eat food in southern China. Sci. Rep. 2020, 10, 15175. [Google Scholar] [CrossRef]
- Zabat, M.A.; Sano, W.H.; Cabral, D.J.; Wurster, J.I.; Belenky, P. The impact of vegan production on the kimchi microbiome. Food Microbiol. 2018, 74, 171–178. [Google Scholar] [CrossRef]
- Rausch, P.; Rühlemann, M.; Hermes, B.M.; Doms, S.; Dagan, T.; Dierking, K.; Baines, J.F. Comparative analysis of amplicon and metagenomic sequencing methods reveals key features in the evolution of animal metaorganisms. Microbiome 2019, 7, 33. [Google Scholar] [CrossRef]
- Winand, R.; Bogaerts, B.; Hoffman, S.; Lefevre, L.; Delvoye, M.; Van Braekel, J.; Vanneste, K. Targeting the 16s rRNA gene for bacterial identification in complex mixed samples: Comparative evaluation of second (illumina) and third (oxford nanopore technologies) generation sequencing technologies. Int. J. Mol. Sci. 2020, 21, 298. [Google Scholar] [CrossRef]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha tea fermentation: Microbial and biochemical dynamics. Int. J. Food Microbiol. 2016, 220, 63–72. [Google Scholar] [CrossRef]
- Erdem, B. Characterization of probiotic abilities of LAB from traditional pickle juice and shalgam. Res. Rev. Sci. Math. 2021, 1, 33–50. [Google Scholar]
- Çoban, E.P.; Touray, M.; Törün, B.; Biyik, H.H. Biodiversity of Bacteria Isolated from Home-Made Wine and Vinegar. Int. J. Second. Metab. 2018, 5, 42–48. [Google Scholar]
- Zhang, J.; Wang, X.; Huo, D.; Li, W.; Hu, Q.; Xu, C.; Li, C. Metagenomic approach reveals microbial diversity and predictive microbial metabolic pathways in Yucha, a traditional Li fermented food. Sci. Rep. 2016, 6, 32524. [Google Scholar] [CrossRef]
- Xie, M.; An, F.; Yue, X.; Liu, Y.; Shi, H.; Yang, M.; Wu, R. Characterization and comparison of metaproteomes in traditional and commercial dajiang, a fermented soybean paste in northeast China. Food Chem. 2019, 301, 125270. [Google Scholar] [CrossRef]
- Hussain, B.; Chen, J.S.; Hsu, B.M.; Chu, I.; Koner, S.; Chen, T.H.; Chan, M.W. Deciphering Bacterial Community Structure, Functional Prediction and Food Safety Assessment in Fermented Fruits Using Next-Generation 16S rRNA Amplicon Sequencing. Microorganisms 2021, 9, 1574. [Google Scholar] [CrossRef] [PubMed]
- Kok, C.R.; Hutkins, R. Yogurt and other fermented foods as sources of health-promoting bacteria. Nutr. Rev. 2018, 76 (Suppl. S1), 4–15. [Google Scholar] [CrossRef] [PubMed]
- Pittia, P.; Antonello, P. Safety by control of water activity: Drying, smoking, and salt or sugar addition. In Regulating Safety of Traditional and Ethnic Foods; Academic Press: Cambridge, MA, USA, 2016; pp. 7–28. [Google Scholar]
- Kunene, N.F.; Geornaras, I.; von Holy, A.; Hastings, J.W. Characterization and determination of origin of lactic acid bacteria from a sorghum-based fermented weaning food by analysis of soluble proteins and amplified fragment length polymorphism fingerprinting. Appl. Environ. Microbiol. 2000, 66, 1084–1092. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ryu, J.A.; Kim, E.; Kim, M.J.; Lee, S.; Yoon, S.R.; Ryu, J.G.; Kim, H.Y. Physicochemical characteristics and microbial communities in gochujang, a traditional korean fermented hot pepper paste. Front. Microbiol. 2021, 11, 620478. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Tang, X.; Yang, R.; Zhang, H.; Li, F.; Tao, F.; Wang, Z. Characteristics and application of a novel species of Bacillus: Bacillus velezensis. ACS Chem. Biol. 2018, 13, 500–505. [Google Scholar] [CrossRef]
- Çakmakçı, R. Bitki gelişimini teşvik eden rizobakterilerin tarımda kullanımı. Atatürk Üniv. Ziraat Fak. Derg. 2005, 36, 97–107. [Google Scholar]
- Saharan, B.S.; Nehra, V. Plant growth promoting rhizobacteria: A critical review. Life Sci. Med. Res. 2011, 21, 30. [Google Scholar]
- González-Arenzana, L.; Portu, J.; López, R.; Garijo, P.; Garde-Cerdán, T.; López-Alfaro, I. Phenylalanine and urea foliar application: Effect on grape and must microbiota. Int. J. Food Microbiol. 2017, 245, 88–97. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G. Arthrobacter. In Encyclopedia of Food Microbiology, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 69–76. [Google Scholar]
- Gumulya, Y.; Boxall, N.J.; Khaleque, H.N.; Santala, V.; Carlson, R.P.; Kaksonen, A.H. In a quest for engineering acidophiles for biomining applications: Challenges and opportunities. Genes 2018, 9, 116. [Google Scholar] [CrossRef]
- Phewpan, A.; Phuwaprisirisan, P.; Takahashi, H.; Ohshima, C.; Lopetcharat, K.; Techaruvichit, P.; Keeratipibul, S. Microbial diversity during processing of Thai traditional fermented shrimp paste, determined by next generation sequencing. LWT 2020, 122, 108989. [Google Scholar] [CrossRef]
- Pakdeeto, A.; Tanasupawat, S.; Thawai, C.; Moonmangmee, S.; Kudo, T.; Itoh, T. Lentibacillus kapialis sp. nov., from fermented shrimp paste in Thailand. Int. J. Syst. Evol. 2007, 57, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.Y.; Wang, D.Q.; Xu, J.; Yang, F.; Liu, G.; Jin, T. Dynamic changes in the bacterial community in Moutai liquor fermentation process characterized by deep sequencing. J. Inst. Brew. 2015, 121, 603–608. [Google Scholar] [CrossRef]
- Oh, Y.J.; Kim, J.Y.; Jo, H.E.; Park, H.K.; Lim, S.K.; Kwon, M.S.; Choi, H.J. Lentibacillus cibarius sp. nov., isolated from kimchi, a Korean fermented food. J. Microbiol. 2020, 58, 387–394. [Google Scholar] [CrossRef]
- Ma, M.; Wang, Z.; Li, L.; Jiang, X.; Guan, D.; Cao, F.; Li, J. Complete genome sequence of Paenibacillus mucilaginosus 3016, a bacterium functional as microbial fertilizer. J. Bacteriol. 2012, 194, 2777–2778. [Google Scholar] [CrossRef]
- Gallardo, O.; Pastor, F.J.; Polaina, J.; Diaz, P.; Łysek, R.; Vogel, P.; Sanz-Aparicio, J. Structural insights into the specificity of Xyn10B from Paenibacillus barcinonensis and its improved stability by forced protein evolution. J. Biol. Chem. 2010, 285, 2721–2733. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Solanki, M.K.; Yu, Z.X.; Anas, M.; Dong, D.F.; Xing, Y.X.; Li, Y.R. Genome characteristics reveal the biocontrol potential of actinobacteria isolated from sugarcane rhizosphere. Front. Microbiol. 2021, 12, 797889. [Google Scholar] [CrossRef]
- Sachdev, D.P.; Chaudhari, H.G.; Kasture, V.M.; Dhavale, D.D.; Chopade, B.A. Isolation and characterization of indole acetic acid (IAA) producing Klebsiella pneumoniae strains from rhizosphere of wheat (Triticum aestivum) and their effect on plant growth. Indian J. Exp. Biol. 2009, 47, 993–1000. [Google Scholar]
- Mello, L.D.P.S.; dos Santos, A.C.; dos Santos, R.M.; Kandasamy, S.; Lazarovits, G.; Rigobelo, E.C. Application of the bacterial strains’ Ruminobacter amylophilus, Fibrobacter succinogenes and Enterococcus faecium for growth promotion in maize and soybean plants. Aust. J. Crop Sci. 2020, 14, 2020–2027. [Google Scholar] [CrossRef]
- Valenzuela-Ruiz, V.; Robles-Montoya, R.I.; Parra-Cota, F.I.; Santoyo, G.; del Carmen Orozco-Mosqueda, M.; Rodríguez-Ramírez, R.; de Los Santos-Villalobos, S. Draft genome sequence of Bacillus paralicheniformis TRQ65, a biological control agent and plant growth-promoting bacterium isolated from wheat (Triticum turgidum subsp. durum) rhizosphere in the Yaqui Valley, Mexico. 3 Biotech 2019, 9, 436. [Google Scholar] [CrossRef]
- Sun, X.; Xu, Z.; Xie, J.; Hesselberg-Thomsen, V.; Tan, T.; Zheng, D.; Kovács, Á.T. Bacillus velezensis stimulates resident rhizosphere Pseudomonas stutzeri for plant health through metabolic interactions. ISME J. 2022, 16, 774–787. [Google Scholar] [CrossRef]
- Kamilova, F.; Leveau, J.H.; Lugtenberg, B. Collimonas fungivorans, an unpredicted in vitro but efficient in vivo biocontrol agent for the suppression of tomato foot and root rot. Environ. Microbiol. 2007, 9, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.T.; Rediers, H.; Ghequire, M.G.; Nguyen, H.H.; De Mot, R.; Vanderleyden, J.; Spaepen, S. The plant growth-promoting effect of the nitrogen-fixing endophyte Pseudomonas stutzeri A15. Arch. Microbiol. 2017, 199, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Rokhbakhsh-Zamin, F.; Sachdev, D.; Kazemi-Pour, N.; Engineer, A.; Pardesi, K.R.; Zinjarde, S.; Chopade, B.A. Characterization of plant-growth-promoting traits of Acinetobacter species isolated from rhizosphere of Pennisetum glaucum. J. Microbiol. Biotechnol. 2011, 21, 556–566. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef]
- Silva, H.S.; Tozzi, J.P.; Terrasan, C.R.; Bettiol, W. Endophytic microorganisms from coffee tissues as plant growth promoters and biocontrol agents of coffee leaf rust. Biol. Control 2012, 63, 62–67. [Google Scholar] [CrossRef]
- Li, S.; Niu, Y.; Chen, H.; He, P. Complete genome sequence of an Arctic Ocean bacterium Shewanella sp. Arc9-LZ with capacity of synthesizing silver nanoparticles in darkness. Mar. Genom. 2021, 56, 100808. [Google Scholar] [CrossRef]
- Farnworth, E.R.; Champagne, C. Production of probiotic cultures and their incorporation into foods. In Bioactive Foods in Promoting Health; Academic Press: Cambridge, MA, USA, 2010; pp. 3–17. [Google Scholar]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Edible table (bio) spread containing potentially probiotic Lactobacillus and Bifidobacterium species. Int. J. Dairy Technol. 2002, 55, 44–56. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Costa, G.A.; Regenhardt, S.; Reinheimer, J.A. Influence of compounds associated with fermented dairy products on the growth of lactic acid starter and probiotic bacteria. Int. Dairy J. 2002, 12, 579–589. [Google Scholar] [CrossRef]
- Ding, W.K.; Shah, N.P. Acid, bile, and heat tolerance of free and microencapsulated probiotic bacteria. J. Food Sci. 2007, 72, M446–M450. [Google Scholar] [CrossRef]
- Toscano, M.; Vecchi, E.D.; Rodighiero, V.; Drago, L. Microbiological and genetic identification of some probiotics proposed for medical use in 2011. J. Chemother. 2013, 25, 156–161. [Google Scholar] [CrossRef]
- İyiçınar, H. Kontrollü Şartlarda Şalgam Suyu Üretimi Üzerine Farklı Formulasyonların Etkisi. Master’s Thesis, Selcuk University Institute of Science and Technology, Konya, Turkey, 2007. [Google Scholar]
- Cueva, C.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bills, G.; Vicente, M.F.; Basilio, A.; Bartolomé, B. Antimicrobial activity of phenolic acids against commensal, probiotic and pathogenic bacteria. Res. Microbiol. 2010, 161, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Sabel, A.; Bredefeld, S.; Schlander, M.; Claus, H. Wine phenolic compounds: Antimicrobial properties against yeasts, lactic acid and acetic acid bacteria. Beverages 2017, 3, 29. [Google Scholar] [CrossRef]
- Irwin, S.V.; Fisher, P.; Graham, E.; Malek, A.; Robidoux, A. Sulfites inhibit the growth of four species of beneficial gut bacteria at concentrations regarded as safe for food. PLoS ONE 2017, 12, e0186629. [Google Scholar] [CrossRef] [PubMed]
- Hrncirova, L.; Hudcovic, T.; Sukova, E.; Machova, V.; Trckova, E.; Krejsek, J.; Hrncir, T. Human gut microbes are susceptible to antimicrobial food additives in vitro. Folia Microbiol. 2019, 64, 497–508. [Google Scholar] [CrossRef]
- Lin, W.H.; Hwang, C.F.; Chen, L.W.; Tsen, H.Y. Viable counts, characteristic evaluation for commercial lactic acid bacteria products. Food Microbiol. 2006, 23, 74–81. [Google Scholar] [CrossRef]
- Baliyan, N.; Kumari, M.; Kumari, P.; Dindhoria, K.; Mukhia, S.; Kumar, S.; Kumar, R. Probiotics in fermented products and supplements. In Current Developments in Bioengineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 73–107. [Google Scholar]
- Steele, M.; Odumeru, J. Irrigation water as source of foodborne pathogens on fruit and vegetables. J. Food Prot. 2004, 67, 2839–2849. [Google Scholar] [CrossRef]
- Snyder, A.; Breidt, F., Jr.; Andress, E.L.; Ingham, B.H. Manufacture of traditionally fermented vegetable products: Best practice for small businesses and retail food establishments. Food Prot. Trends. 2020, 40, 251–263. [Google Scholar]
- Bressan, A. Emergence and evolution of Arsenophonus bacteria as insect-vectored plant pathogens. Infect. Genet. Evol. 2014, 22, 81–90. [Google Scholar] [CrossRef]
- Shi, Y.; Lou, K.; Li, C. Growth promotion effects of the endophyte Acinetobacter johnsonii strain 3-1 on sugar beet. Symbiosis 2011, 54, 159–166. [Google Scholar] [CrossRef]
- Humblot, C.; Guyot, J.P. Pyrosequencing of tagged 16S rRNA gene amplicons for rapid deciphering of the microbiomes of fermented foods such as pearl millet slurries. Appl. Environ. Microbiol. 2009, 75, 4354–4361. [Google Scholar] [CrossRef]
- Xie, M.; An, F.; Zhao, Y.; Wu, R.; Wu, J. Metagenomic analysis of bacterial community structure and functions during the fermentation of da-jiang, a Chinese traditional fermented food. LWT 2020, 129, 109450. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermented Products | Sample Code | Number of Subsamples | Production Process Applicatıon Type * | Number of Total Subsamples | |
|---|---|---|---|---|---|
| Food Additive | Pasteurization | ||||
| Hardaliye | C1 | 4 | YES | NO | 16 |
| C2 | 4 | YES | YES | ||
| C3 | 4 | YES | YES | ||
| C4 | 4 | YES | NO | ||
| Kombucha | C5 | 4 | NO | NO | 20 |
| C6 | 4 | NO | NO | ||
| C7 | 4 | NO | NO | ||
| C8 | 4 | NO | NO | ||
| C9 | 4 | NO | NO | ||
| Shalgam Juice | C10 | 4 | YES | NO | 20 |
| C11 | 4 | NO | NO | ||
| C12 | 4 | NO | YES | ||
| C13 | 4 | YES | YES | ||
| C14 | 4 | YES | NO | ||
| Grape Vinegar | C15 | 4 | YES | NO | 20 |
| C16 | 4 | NO | NO | ||
| C17 | 4 | NO | NO | ||
| C18 | 4 | NO | YES | ||
| C19 | 4 | YES | YES | ||
| Pickle Juice | C20 | 4 | NO | NO | 20 |
| C21 | 4 | NO | YES | ||
| C22 | 4 | NO | NO | ||
| C23 | 4 | NO | NO | ||
| C24 | 4 | NO | NO | ||
| Number of Total Samples | 96 | ||||
| Plant Disease Prevention and Plant Growth Regulation Bacteria According to the Literature | The Relative Abundance of Plant Growth Regulator or Promoting Bacteria in Our Samples | ||||
|---|---|---|---|---|---|
| Hardaliye | Kombucha | Shalgam Juice | Pickle Juice | Grape Vinegar | |
| Actinobacteria [38] | 3 | 2.4 | 1.5 | 1.5 | - |
| Arthrobacter [27] | 1 | - | - | - | - |
| Klebsiella [39] | 1.4 | 1.5 | - | - | - |
| Paenibacillus mucilaginosus [36] | - | - | 15.5 | 14.2 | 3 |
| Paenibacillus barcinonensis [36] | - | - | 8.9 | 8.8 | 3 |
| Enterococcus faecium [40] | - | - | 9.7 | 12 | 3 |
| Bacillus spp. [27] | 1.5 | 1.5 | 14 | 14.1 | 14.7 |
| Bacillus paralicheniformis [41] | - | - | - | - | 11.76 |
| Bacillus velezensis [42] | - | - | 3 | 3 | - |
| Collimonas fungivorans [43] | 6.1 | 6.1 | 1 | - | 2.9 |
| Pseudomonas stutzeri [44] | 5 | 5 | 1.1 | - | - |
| Acinetobacter spp. [45] | 4.4 | 4.3 | - | - | 2.9 |
| Burkholderia thailandensis [46] | 1.9 | 1.6 | - | - | - |
| Salmonella enterica [47] | 3.8 | 4.2 | - | - | - |
| Shewanella sp. Arc9-LZ [48] | 1.1 | 1.3 | - | - | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çelik Doğan, C.; Yüksel Dolgun, H.T.; İkiz, S.; Kırkan, Ş.; Parın, U. Detection of the Microbial Composition of Some Commercial Fermented Liquid Products via Metagenomic Analysis. Foods 2023, 12, 3538. https://doi.org/10.3390/foods12193538
Çelik Doğan C, Yüksel Dolgun HT, İkiz S, Kırkan Ş, Parın U. Detection of the Microbial Composition of Some Commercial Fermented Liquid Products via Metagenomic Analysis. Foods. 2023; 12(19):3538. https://doi.org/10.3390/foods12193538
Chicago/Turabian StyleÇelik Doğan, Cansu, Hafize Tuğba Yüksel Dolgun, Serkan İkiz, Şükrü Kırkan, and Uğur Parın. 2023. "Detection of the Microbial Composition of Some Commercial Fermented Liquid Products via Metagenomic Analysis" Foods 12, no. 19: 3538. https://doi.org/10.3390/foods12193538
APA StyleÇelik Doğan, C., Yüksel Dolgun, H. T., İkiz, S., Kırkan, Ş., & Parın, U. (2023). Detection of the Microbial Composition of Some Commercial Fermented Liquid Products via Metagenomic Analysis. Foods, 12(19), 3538. https://doi.org/10.3390/foods12193538