

The Degradation of Intramuscular Connective Tissue In Vitro with Purified Cathepsin L from Bovine Pancreas

Abstract
1. Introduction
2. Materials and Methods
2.1. IMCT Preparation
2.2. The Preparation of Purified Cathepsin L
2.3. The Incubation of IMCT with Purified Cathepsin L, CNBr Digestion, and SDS-PAGE
2.4. A Western Blot Analysis of the Degradation of Decorin (DCN) Incubated with Cathepsin L In Vitro
2.5. Differential Scanning Calorimetry (DSC)
2.6. Statistical Analysis
3. Results and Discussion
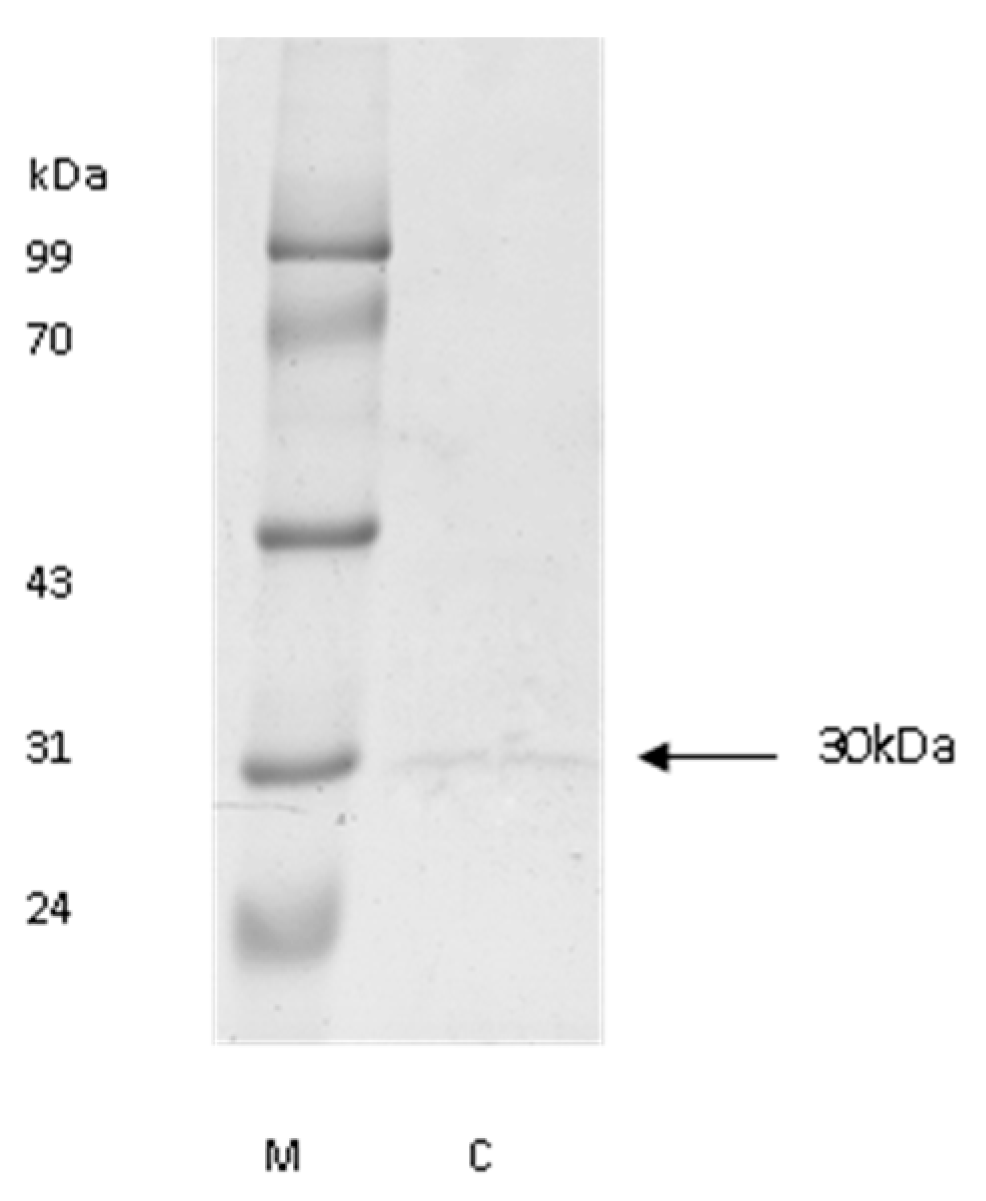
3.1. The Purification of Cathepsin L from Bovine Pancreas
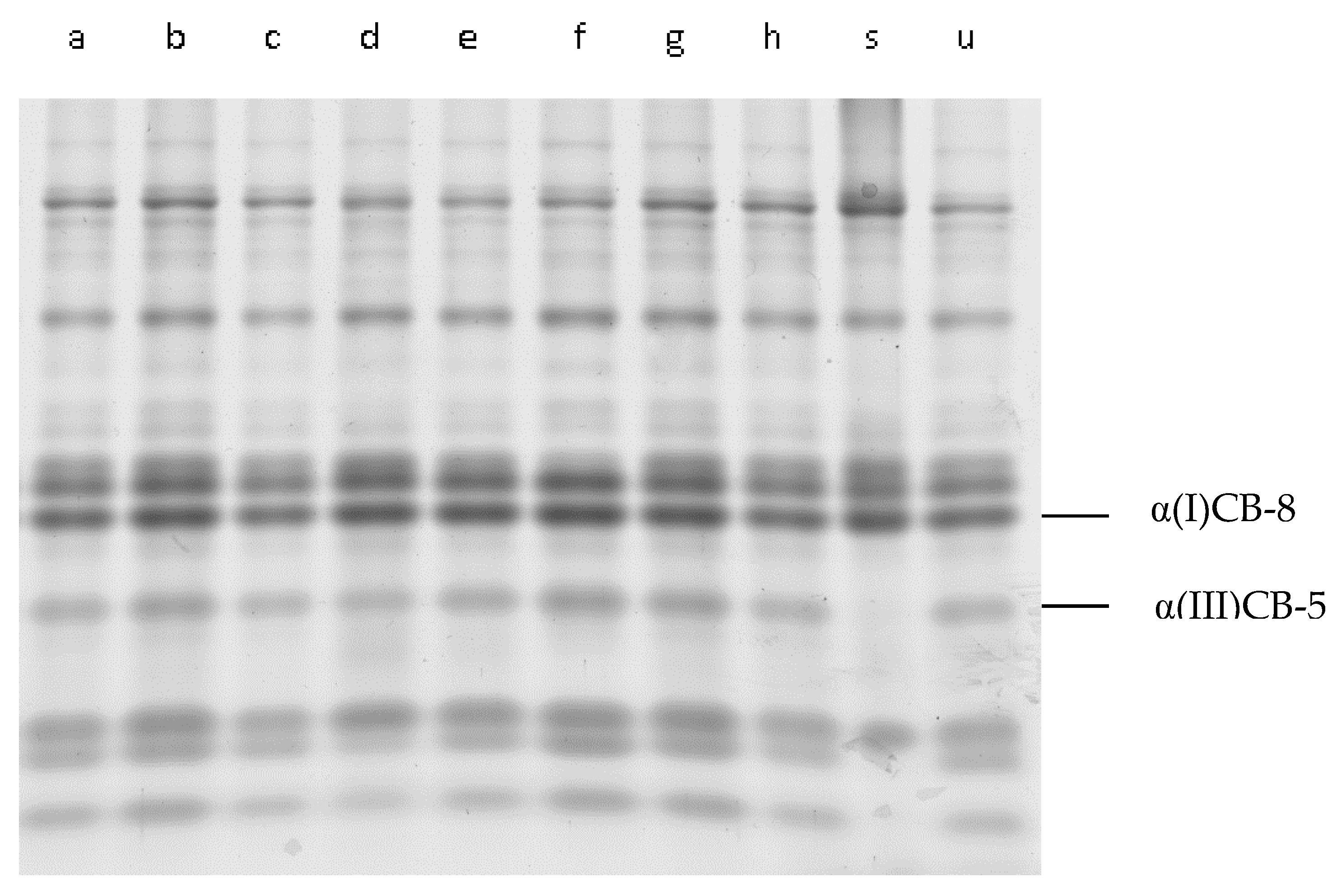
3.2. CNBr-Digested Peptides from Cathepsin L-Treated IMCT
3.3. The Degradation of DCN
3.4. The Analysis of IMCT by DSC
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Warner, R.D.; Wheeler, T.L.; Ha, M.; Li, X.; Bekhit, A.E.; Morton, J.; Vaskoska, R.; Dunshea, F.R.; Liu, R.; Purslow, P.; et al. Meat tenderness: Advances in biology, biochemistry, molecular mechanisms and new technologies. Meat Sci. 2021, 185, 108657. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T. The role of intramuscular connective tissue in meat texture. Anim. Sci. J. 2010, 81, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, D.E.; Latorre, M.E. Physicochemical, thermal and mechanical characterization study of perimysial collagen of two bovine muscles. Int. J. Biol. Macromol. 2019, 136, 404–409. [Google Scholar] [CrossRef]
- Christensen, M.; Ertbjerg, P.; Failla, S.; Sañudo, C.; Richardson, R.I.; Nute, G.R.; Olleta, J.L.; Panea, B.; Albertí, P.; Juárez, M.; et al. Relationship between collagen characteristics, lipid content and raw and cooked texture of meat from young bulls of fifteen European breeds. Meat Sci. 2010, 87, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.L.; Allingham, P.G.; Colgrave, M.; van de Ven, R.J. Interrelationship between measures of collagen, compression, shear force and tenderness. Meat Sci. 2013, 95, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Aktaş, N.; Kaya, M. Influence of weak organic acids and salts on the denaturation characteristics of intramuscular connective tissue. A differential scanning calorimetry study. Meat Sci. 2001, 58, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Latorre, M.E.; Velázquez, D.E.; Purslow, P.P. Differences in the energetics of collagen denaturation in connective tissue from two muscles. Int. J. Biol. Macromol. 2018, 113, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Lepetit, J. A theoretical approach of the relationships between collagen content, collagen cross-links and meat tenderness. Meat Sci. 2007, 76, 147–159. [Google Scholar] [CrossRef]
- Nishiumi, T.; Fukuda, T.; Nishimura, T. Isolation and characterization of a small proteoglycan associated with porcine intramuscular connective tissue. J. Agric. Food Chem. 1997, 45, 2978–2983. [Google Scholar] [CrossRef]
- Kaur, L.; Hui, S.X.; Boland, M. Changes in Cathepsin Activity during Low-Temperature Storage and Sous Vide Processing of Beef Brisket. Food Sci. Anim. Resour. 2020, 40, 415–425. [Google Scholar] [CrossRef]
- Kemp, C.M.; Sensky, P.L.; Bardsley, R.G.; Buttery, P.J.; Parr, T. Tenderness—An enzymatic view. Meat Sci. 2010, 84, 248–256. [Google Scholar] [CrossRef]
- Etherington, D.J. The purification of bovine cathepsin B1 and its mode of action on bovine collagens. Biochem. J. 1974, 137, 547–557. [Google Scholar] [CrossRef]
- Kafienah, W.; Bromme, D.; Buttle, D.; Croucher, L.; Hollander, A. Human cathepsin K cleaves native type I and II collagens at the N-terminal end of the triple helix. Biochem. J. 1998, 331, 727–732. [Google Scholar] [CrossRef]
- Ertbjerg, P.; Larsen, L.M.; Moøller, A.J. Effect of prerigor lactic acid treatment on lysosomal enzyme release in bovine muscle. J. Sci. Food Agric. 1999, 79, 95–100. [Google Scholar] [CrossRef]
- Beltrán, J.A.; Bonnet, M.; Ouali, A. Comparative action of cathepsins B and L on intramuscular collagen as assessed by differential scanning calorimetry. Meat Sci. 1992, 32, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Hattori, A.; Takahashi, K. Relationship between degradation of proteoglycans and weakening of the intramuscular connective tissue during post-mortem ageing of beef. Meat Sci. 1996, 42, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-J.; Wang, Q.; Zhou, G.-H.; Xu, X.-L.; Li, C.-B. Influence of Weak Organic Acids and Sodium Chloride Marination on Characteristics of Connective Tissue Collagen and Textural Properties of Beef Semitendinosus Muscle. J. Texture Stud. 2010, 41, 279–301. [Google Scholar] [CrossRef]
- Loveday, S.M.; Wang, X.L.; Rao, M.A.; Anema, S.G.; Creamer, L.K.; Singh, H. Tuning the properties of beta-lactoglobulin nanofibrils with pH, NaCl and CaCl2. Int. Dairy J. 2010, 20, 571–579. [Google Scholar] [CrossRef]
- Tang, Q.; Wang, W.; Zhang, L.; Liu, Y. Cloning, purification and biochemical characterization of recombinant Cathepsin L from Takifugu rubripes and its role in taste formation. J. Food Meas. Charact. 2020, 14, 485–491. [Google Scholar] [CrossRef]
- Li, S.; Zhou, X.; Zhang, N.; Liu, H.; Ma, C. Purification and characterisation of cathepsin L2 from dorsal muscle of silver carp (Hypophthalmichthys molitrix). Food Chem. 2008, 111, 879–886. [Google Scholar] [CrossRef]
- Gillies, A.; Lieber, R. Muscle fibrosis due to loss of desmin increases muscle stiffness due to an increased number of perimysial collagen cables (1102.8). FASEB J. 2014, 28, 1102.8. [Google Scholar] [CrossRef]
- Wang, R.; Peng, Z.; Hui, T.; Wang, F.; Yao, Y.; Zhang, Y.; Zhou, G. Potential use of crude extracts from Alaska Pollock muscle as meat tenderizer. CyTA-J. Food 2013, 11, 50–59. [Google Scholar] [CrossRef][Green Version]
- López De Padilla, C.M.; Coenen, M.J.; Tovar, A.; De la Vega, R.E.; Evans, C.H.; Müller, S.A. Picrosirius Red Staining: Revisiting Its Application to the Qualitative and Quantitative Assessment of Collagen Type I and Type III in Tendon. J. Histochem. Cytochem. 2021, 69, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Scarr, G. Fascial hierarchies and the relevance of crossed-helical arrangements of collagen to changes in the shape of muscles. J. Bodyw. Mov. Ther. 2016, 20, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Vidal, B.D.C. Fluorescence, aggregation properties and FT-IR microspectroscopy of elastin and collagen fibers. Acta Histochem. 2014, 116, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Kishioka, Y.; Wakamatsu, J.; Nishimura, T. Decorin activates Akt downstream of IGF-IR and promotes myoblast differentiation. Anim. Sci. J. 2013, 84, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Honardoust, D.; Varkey, M.; Marcoux, Y.; Shankowsky, H.A.; Tredget, E.E. Reduced decorin, fibromodulin, and transforming growth factor-β3 in deep dermis leads to hypertrophic scarring. J. Burn Care Res. 2012, 33, 218–227. [Google Scholar] [CrossRef]
- Huang, F.; Huang, M.; Zhou, G.; Xu, X.; Xue, M. In vitro proteolysis of myofibrillar proteins from beef skeletal muscle by caspase-3 and caspase-6. J. Agric. Food Chem. 2011, 59, 9658–9663. [Google Scholar] [CrossRef]
- Castrejón-Flores, J.L.; Reyna-Luna, J.; Flores-Martinez, Y.M.; García-Ventura, M.I.; Zamudio-Medina, A.; Franco-Pérez, M. Characterizing the thermal degradation mechanism of two bisphosphoramidates by TGA, DSC, mass spectrometry and first-principle theoretical protocols. J. Mol. Struct. 2020, 1221, 128781. [Google Scholar] [CrossRef]
- Hu, Y.; Ji, R.; Jiang, H.; Zhang, J.; Chen, J.; Ye, X. Participation of cathepsin L in modori phenomenon in carp (Cyprinus carpio) surimi gel. Food Chem. 2012, 134, 2014–2020. [Google Scholar] [CrossRef]
- Zhong, C.; Cai, Q.-F.; Liu, G.-M.; Sun, L.-C.; Hara, K.; Su, W.-J.; Cao, M.-J. Purification and characterisation of cathepsin L from the skeletal muscle of blue scad (Decapterus maruadsi) and comparison of its role with myofibril-bound serine proteinase in the degradation of myofibrillar proteins. Food Chem. 2012, 133, 1560–1568. [Google Scholar] [CrossRef]
- Sun, K.; Yang, F.; Kong, Y.; Kang, J.; Cao, W.; Yang, X.; Zha, S.; Zhang, G.; Wang, M. Quantitative Detection Method of Collagen Based on Biomass Spectrometry. Chin. J. Bio. Eng. 2015, 31, 1660–1668. [Google Scholar]
- Kuivaniemi, H.; Tromp, G. Type III collagen (COL3A1): Gene and protein structure, tissue distribution, and associated diseases. Gene 2019, 707, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.; Biswas, S. Structure-guided protein engineering of human cathepsin L for efficient collagenolytic activity. PEDS 2021, 34, gzab005. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, Y.; Li, J.; Guo, X.; Cui, B.; Peng, Z. Contribution of cross-links and proteoglycans in intramuscular connective tissue to shear force in bovine muscle with different marbling levels and maturities. LWT-Food Sci. Technol. 2016, 66, 413–419. [Google Scholar] [CrossRef]
- Dourte, L.M.; Pathmanathan, L.; Jawad, A.F.; Iozzo, R.V.; Mienaltowski, M.J.; Birk, D.E.; Soslowsky, L.J. Influence of decorin on the mechanical, compositional, and structural properties of the mouse patellar tendon. J. Biomech. Eng. 2012, 134, 031005. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.A.; Sun, M.; Barnum, C.E.; Weiss, S.N.; Huegel, J.; Shetye, S.S.; Lin, L.; Saez, D.; Adams, S.M.; Iozzo, R.V.; et al. Decorin and biglycan are necessary for maintaining collagen fibril structure, fiber realignment, and mechanical properties of mature tendons. Matrix Biol. 2017, 64, 81–93. [Google Scholar] [CrossRef]
- Voutila, L.; Ruusunen, M.; Puolanne, E. Comparison of the thermal characteristics of connective tissue in loose structured and normal structured porcine M. semimembranosus. Meat Sci. 2008, 80, 1024–1030. [Google Scholar] [CrossRef]
- Aktaş, N. The effects of pH, NaCl and CaCl2 on thermal denaturation characteristics of intramuscular connective tissue. Thermochim. Acta 2003, 407, 105–112. [Google Scholar] [CrossRef]
- Garnero, P.; Borel, O.; Byrjalsen, I.; Ferreras, M.; Drake, F.H.; McQueney, M.S.; Foged, N.T.; Delmas, P.D.; Delaissé, J.-M. The collagenolytic activity of cathepsin K is unique among mammalian proteinases. J. Biol. Chem. 2018, 273, 32347–32352. [Google Scholar] [CrossRef]
- Kirschke, H.; Kembhavi, A.A.; Bohley, P.; Barrett, A.J. Action of rat liver cathepsin L on collagen and other substrates. Biochem. J. 1982, 201, 367–372. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Untreated | Time | Untreated | ||||||
|---|---|---|---|---|---|---|---|---|---|
| C-36 h | 24 h | 36 h | C-36 h | 24 h | 36 h | ||||
| TO-5.0 (°C) | 57.87 ± 1.09 aX | 54.90 ± 0.82 bX | 41.41 ± 0.60 cX | 60.63 ± 0.72 d | TP-5.0 (°C) | 62.04 ± 1.02 aX | 60.76 ± 0.70 aX | 43.79 ± 0.66 bX | 65.38 ± 1.65 c |
| TO-5.5 (°C) | 59.73 ± 0.50 aX | 59.24 ± 0.93 aY | 56.57 ± 0.71 bY | 60.63 ± 0.72 a | TP-5.5 (°C) | 65.11 ± 0.30 aX | 64.80 ± 1.03 aY | 60.79 ± 1.01 aY | 65.38 ± 1.65 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Y.; He, W.; Teng, S.; Jamali, M.A. The Degradation of Intramuscular Connective Tissue In Vitro with Purified Cathepsin L from Bovine Pancreas. Foods 2023, 12, 3517. https://doi.org/10.3390/foods12183517
Peng Y, He W, Teng S, Jamali MA. The Degradation of Intramuscular Connective Tissue In Vitro with Purified Cathepsin L from Bovine Pancreas. Foods. 2023; 12(18):3517. https://doi.org/10.3390/foods12183517
Chicago/Turabian StylePeng, Yingbo, Wanhong He, Shuang Teng, and Muneer Ahmed Jamali. 2023. "The Degradation of Intramuscular Connective Tissue In Vitro with Purified Cathepsin L from Bovine Pancreas" Foods 12, no. 18: 3517. https://doi.org/10.3390/foods12183517
APA StylePeng, Y., He, W., Teng, S., & Jamali, M. A. (2023). The Degradation of Intramuscular Connective Tissue In Vitro with Purified Cathepsin L from Bovine Pancreas. Foods, 12(18), 3517. https://doi.org/10.3390/foods12183517