
Lachancea quebecensis a Novel Isolate for the Production of Craft Beer



Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Yeasts Isolation and Identification
2.2.1. Isolation
2.2.2. Identification
2.3. Fermentative Profile in Defined Medium
2.3.1. Glucose Defined Medium
2.3.2. Maltose Defined Medium
2.3.3. Resistance to Hop and Ethanol
2.4. Craft Beer Production at Laboratory Scale (10 L)
2.4.1. Propagation
2.4.2. Beer Making
2.5. Sensory Analysis
2.6. Statistical Analysis
3. Results and Discussion
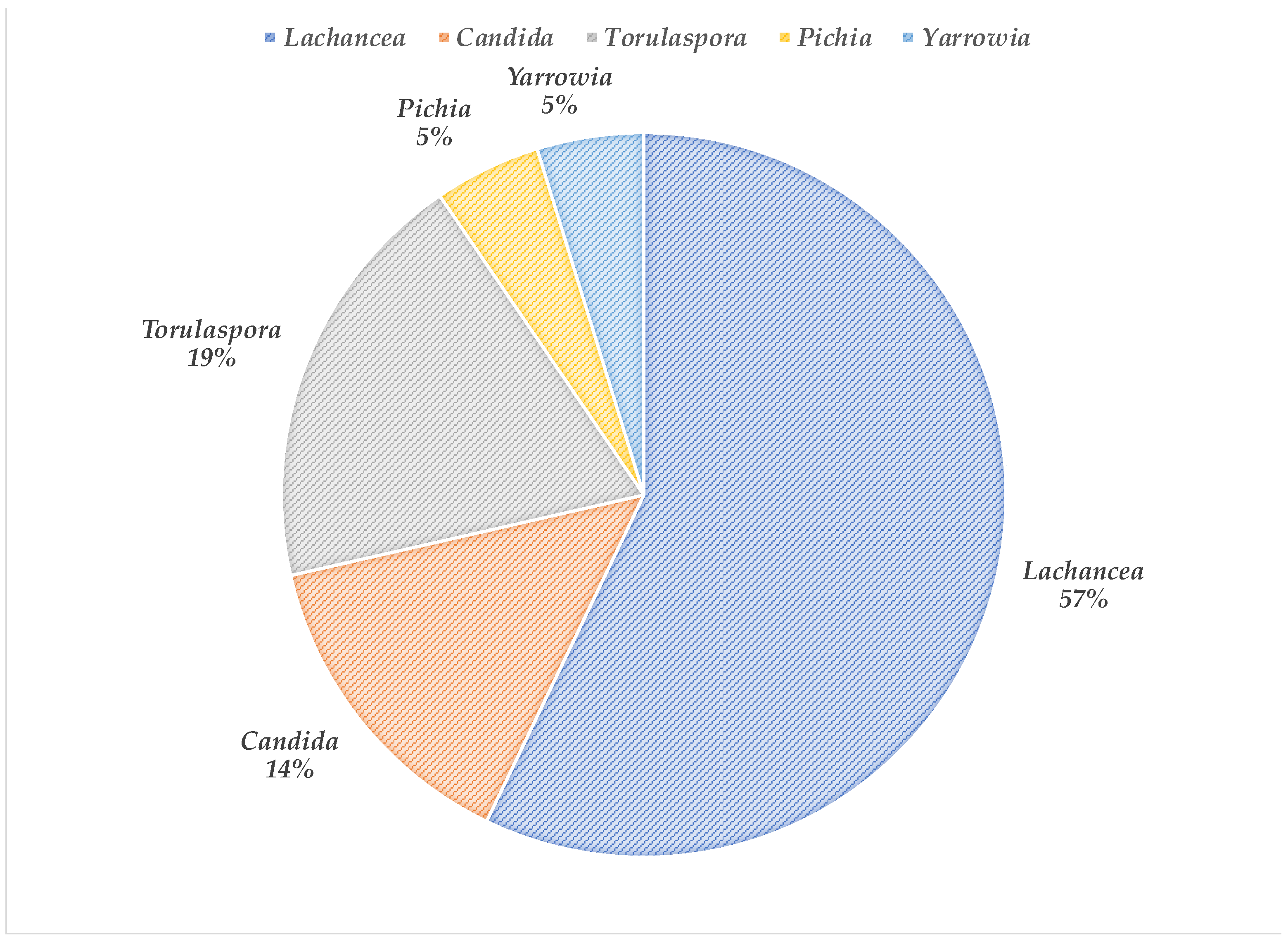
3.1. Yeasts Isolation and Identification
3.2. Screening for the Brewing Potential of Selected Lachance Isolates
Glucose and Maltose Defined Media
3.3. Resistance to Hop and Ethanol
3.4. Pale Ale Craft Beer Production
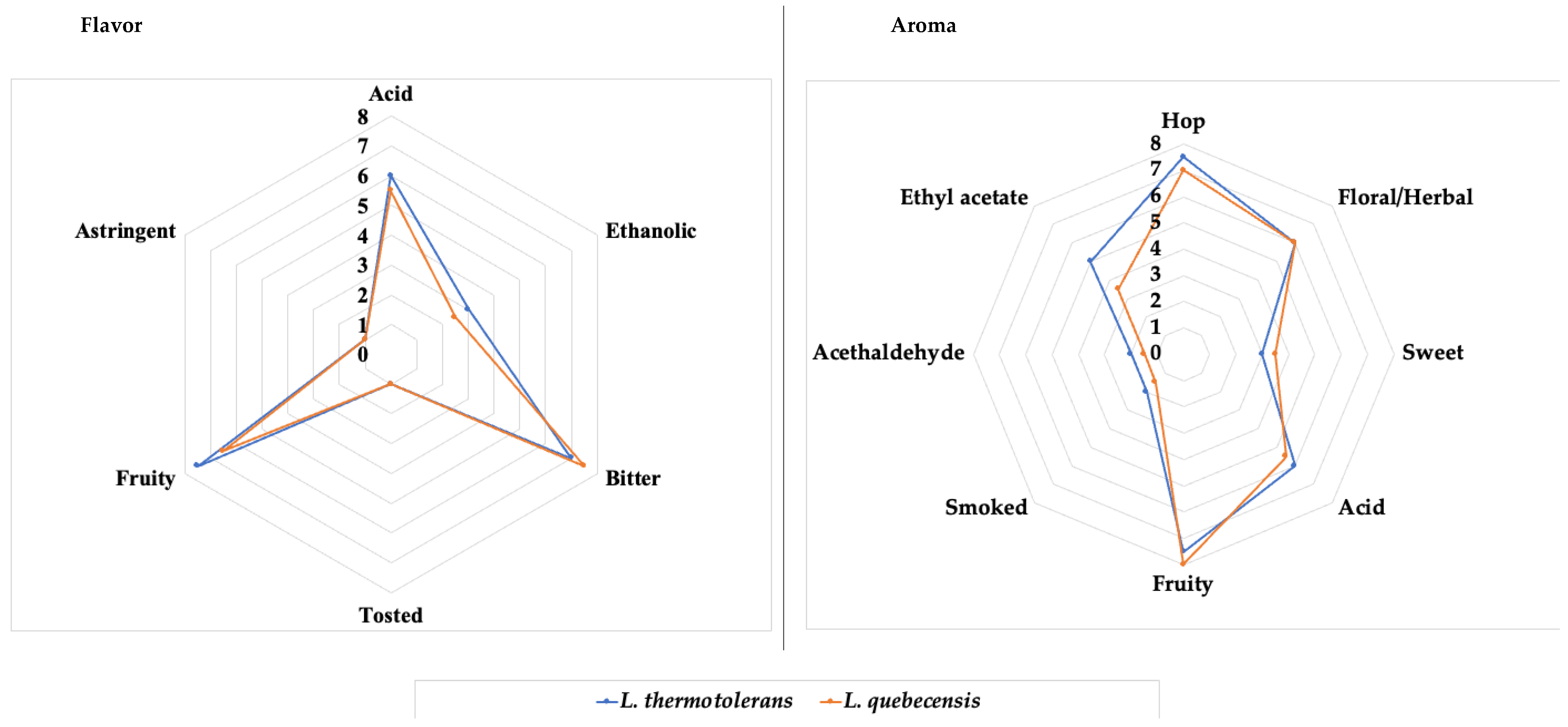
3.5. Beer Aroma and Sensory Attributes
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Borren, E.; Tian, B. The Important Contribution of Non-Saccharomyces Yeasts to the Aroma Complexity of Wine: A Review. Foods 2020, 10, 13. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Bellut, K.; Arendt, E.K. Chance and Challenge: Non-Saccharomyces Yeasts in Nonalcoholic and Low Alcohol Beer Brewing—A Review. J. Am. Soc. Brew. Chem. 2019, 77, 77–91. [Google Scholar] [CrossRef]
- Postigo, V.; Sánchez, A.; Cabellos, J.M.; Arroyo, T. New Approaches for the Fermentation of Beer: Non-Saccharomyces Yeasts from Wine. Fermentation 2022, 8, 280. [Google Scholar] [CrossRef]
- Larroque, M.; Carrau, F.; Fariña, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and non-Saccharomyces native yeasts on beer aroma compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef]
- Methner, Y.; Hutzler, M.; Zarnkow, M.; Prowald, A.; Endres, F.; Jacob, F. Investigation of Non-Saccharomyces Yeast Strains for Their Suitability for the Production of Non-Alcoholic Beers with Novel Flavor Profiles. J. Am. Soc. Brew. Chem. 2022, 80, 341–355. [Google Scholar] [CrossRef]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Esteve-Zarzoso, B.; Cabellos, J.M.; Gil-Díaz, M.; Arroyo, T. Biotechnological potential of non-Saccharomyces yeasts isolated during spontaneous fermentations of Malvar (Vitis vinifera cv. L.). Eur. Food Res. Technol. 2012, 236, 193–207. [Google Scholar] [CrossRef]
- Garofalo, C.; Russo, P.; Beneduce, L.; Massa, S.; Spano, G.; Capozzi, V. Non-Saccharomyces biodiversity in wine and the ‘microbial terroir’: A survey on Nero di Troia wine from the Apulian region, Italy. Ann. Microbiol. 2015, 66, 143–150. [Google Scholar] [CrossRef]
- Madden, A.A.; Lahue, C.; Gordy, C.L.; Little, J.L.; Nichols, L.M.; Calvert, M.D.; Dunn, R.R.; Heil, C.S. Sugar-seeking insects as a source of diverse bread-making yeasts with enhanced attributes. Yeast 2021, 39, 108–127. [Google Scholar] [CrossRef] [PubMed]
- Toh, D.W.K.; Chua, J.Y.; Lu, Y.; Liu, S.Q. Evaluation of the potential of commercial non-Saccharomyces yeast strains of Torulaspora delbrueckii and Lachancea thermotolerans in beer fermentation. Int. J. Food Sci. Technol. 2019, 55, 2049–2059. [Google Scholar] [CrossRef]
- Bellut, K.; Michel, M.; Hutzler, M.; Zarnkow, M.; Jacob, F.; De Schutter, D.P.; Daenen, L.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Investigation into the Potential of Lachancea fermentati Strain KBI 12.1 for Low Alcohol Beer Brewing. J. Am. Soc. Brew. Chem. 2019, 77, 157–169. [Google Scholar] [CrossRef]
- Gatto, V.; Binati, R.L.; Junior, W.J.L.; Basile, A.; Treu, L.; de Almeida, O.G.; Innocente, G.; Campanaro, S.; Torriani, S. New insights into the variability of lactic acid production in Lachancea thermotolerans at the phenotypic and genomic level. Microbiol. Res. 2020, 238, 126525. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018, 8, 14812. [Google Scholar] [CrossRef]
- Domizio, P.; House, J.F.; Joseph, C.M.L.; Bisson, L.F.; Bamforth, C.W. Lachancea thermotolerans as an alternative yeast for the production of beer. J. Inst. Brew. 2016, 122, 599–604. [Google Scholar] [CrossRef]
- Zdaniewicz, M.; Satora, P.; Pater, A.; Bogacz, S. Low Lactic Acid-Producing Strain of Lachancea thermotolerans as a New Starter for Beer Production. Biomolecules 2020, 10, 256. [Google Scholar] [CrossRef]
- Postigo, V.; Esteban, S.; Arroyo, T. Lachancea thermotolerans, an Innovative Alternative for Sour Beer Production. Beverages 2023, 9, 20. [Google Scholar] [CrossRef]
- Beer Drinkers Are Growing Sweet on Sour Beers. Available online: https://www.forbes.com/sites/jeanettehurt/2020/03/11/beer-drinkers-are-growing-sweet-on-sour-beers/?sh=757e84845176 (accessed on 21 May 2023).
- Dysvik, A.; La Rosa, S.L.; De Rouck, G.; Rukke, E.-O.; Westereng, B.; Wicklund, T. Microbial Dynamics in Traditional and Modern Sour Beer Production. Appl. Environ. Microbiol. 2020, 86, e00566-20. [Google Scholar] [CrossRef]
- Osburn, K.; Amaral, J.; Metcalf, S.R.; Nickens, D.M.; Rogers, C.M.; Sausen, C.; Caputo, R.; Miller, J.; Li, H.; Tennessen, J.M.; et al. Primary souring: A novel bacteria-free method for sour beer production. Food Microbiol. 2018, 70, 76–84. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Bellut, K.; Krogerus, K.; Arendt, E.K. Lachancea fermentati Strains Isolated from Kombucha: Fundamental Insights, and Practical Application in Low Alcohol Beer Brewing. Front. Microbiol. 2020, 11, 764. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Suh, S.-O.; Blackwell, M. Five novel Candida species in insect-associated yeast clades isolated from Neuroptera and other insects. Mycologia 2007, 99, 842–858. [Google Scholar] [CrossRef]
- Suh, S.-O.; Nguyen, N.H.; Blackwell, M. Yeasts isolated from plant-associated beetles and other insects: Seven novel Candida species near Candida albicans. FEMS Yeast Res. 2008, 8, 88–102. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification of Clinically Important Ascomycetous Yeasts Based on Nucleotide Divergence in the 5′ End of the Large-Subunit (26S) ribosomal DNA gene. J. Clin. Microbiol. 1997, 35, 1216. [Google Scholar] [CrossRef]
- Altschul, D.J.; Gish, S.F.; Miller, W.; Myers, W.; Lipman, E.W. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- McFeeters, R.F.; Barish, A.O. Sulfite analysis of fruits and vegetables by high-performance liquid chromatography (HPLC) with ultraviolet spectrophotometric detection. J. Agric. Food Chem. 2003, 51, 1513–1517. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dörnberg, T.; Jacob, F.; Methner, F.; Wagner, R.S.; Hutzler, M. Review: Pure non-Saccharomyces starter cultures for beer fermentation with a focus on secondary metabolites and practical applications. J. Inst. Brew. 2016, 122, 569–587. [Google Scholar] [CrossRef]
- Lie, S. The ebc-ninhydrin method for determination of free alpha amino nitrogen. J. Inst. Brew. 1973, 79, 37–41. [Google Scholar] [CrossRef]
- Meier-Dörnberg, T.; Hutzler, M.; Michel, M.; Methner, F.-J.; Jacob, F. The Importance of a Comparative Characterization of Saccharomyces cerevisiae and Saccharomyces Pastorianus Strains for Brewing. Fermentation 2017, 3, 41. [Google Scholar] [CrossRef]
- Lachance, M.A.; Bowles, J.M.; Díaz, M.M.C.; Janzen, D.H. Candida cleridarum, Candida tilneyi and Candida powellii, three new yeast species isolated from insects associated with flowers. Int. J. Syst. Evol. Microbiol. 2001, 51, 1201–1207. [Google Scholar] [CrossRef]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Vitanović, E.; Lopez, J.M.; Aldrich, J.R.; Špika, M.J.; Boundy-Mills, K.; Zalom, F.G. Yeasts Associated with the Olive Fruit Fly Bactrocera oleae (Rossi) (Diptera: Tephritidae) Lead to New Attractants. Agronomy 2020, 10, 1501. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Palomero, F.; Benito, S. Combine Use of Selected Schizosaccharomyces pombe and Lachancea thermotolerans Yeast Strains as an Alternative to the Traditional Malolactic Fermentation in Red Wine Production. Molecules 2015, 20, 9510–9523. [Google Scholar] [CrossRef]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- Freel, K.C.; Charron, G.; Leducq, J.-B.; Landry, C.R.; Schacherer, J. Lachancea quebecensis sp. nov., a yeast species consistently isolated from tree bark in the Canadian province of Québec. Int. J. Syst. Evol. Microbiol. 2015, 65, 3392–3399. [Google Scholar] [CrossRef]
- Mestre, M.C.; Ulloa, J.R.; Rosa, C.A.; Lachance, M.A.; Fontenla, S. Lachancea nothofagi sp. nov., a yeast associated with Nothofagus species in Patagonia, Argentina. Int. J. Syst. Evol. Microbiol. 2010, 60, 2247–2250. [Google Scholar] [CrossRef]
- Hranilovic, A.; Bely, M.; Masneuf-Pomarede, I.; Jiranek, V.; Albertin, W. The evolution of Lachancea thermotolerans is driven by geographical determination, anthropisation and flux between different ecosystems. PLoS ONE 2017, 12, e0184652. [Google Scholar] [CrossRef]
- Bamforth, C. Progress in Brewing Science and Beer Production. Annu. Rev. Chem. Biomol. Eng. 2017, 8, 161–176. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Conventional and Non-Conventional Yeasts in Beer Production. Fermentation 2018, 4, 38. [Google Scholar] [CrossRef]
- Gallone, B.; Mertens, S.; Gordon, J.L.; Maere, S.; Verstrepen, K.J.; Steensels, J. Origins, evolution, domestication and diversity of Saccharomyces beer yeasts. Curr. Opin. Biotechnol. 2018, 49, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of Non-Saccharomyces Yeasts for the Reduction of Alcohol Content in Wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Van Rijswijck, I.M.H.; Wolkers-Rooijackers, J.C.M.; Abee, T.; Smid, E.J. Performance of non-conventional yeasts in co-culture with brewers’ yeast for steering ethanol and aroma production. Microb. Biotechnol. 2017, 10, 1591–1602. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Galli, E.; Ciani, E.; Comitini, F.; Ciani, M. Exploitation of Three Non-Conventional Yeast Species in the Brewing Process. Microorganisms 2019, 7, 11. [Google Scholar] [CrossRef]
- Svendsen, C.I. Investigation of Lachancea thermotolerans as a Novel Single Culture Yeast, North Carolina State University. 2015. Available online: https://www.ptonline.com/articles/how-to-get-better-mfi-results (accessed on 17 December 2022).
- Del Carmen Portillo, M.; Mas, A. Analysis of microbial diversity and dynamics during wine fermentation of Grenache grape variety by high-throughput barcoding sequencing. LWT 2016, 72, 317–321. [Google Scholar] [CrossRef]
- Franco, W.; Benavides, S.; Valencia, P.; Ramírez, C.; Urtubia, A. Native Yeasts and Lactic Acid Bacteria Isolated from Spontaneous Fermentation of Seven Grape Cultivars from the Maule Region (Chile). Foods 2021, 10, 1737. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Insect | Yeast Identification (Number of Colonies) |
|---|---|
| Ladybug | L. thermotolerans (4) |
| Corn weevil | C. boidinii (1) |
| L. quebecensis (1) | |
| L. thermotolerans (2) | |
| Yarrowia sp. (1) | |
| T. delbrueckii (1) | |
| Wheat flour weevil | T. delbrueckii (1) |
| C. boidinii (1) | |
| P. guilliermondii (1) | |
| L. thermotolerans (3) | |
| Cabbage worm | L. thermotolerans (2) |
| Yeast | CFU/mL | Ethanol | pH | Yield (g Etanol/g Glucosa) | Classification According to Ethanol Yield * |
|---|---|---|---|---|---|
| L. thermotolerans ** | 3.82 × 108 | 3.25 ± 1.77 | 3.67 ± 0.01 | 0.17 | Very low |
| L. quebecensis ** | 1.25 × 109 | 4.25 ± 1.06 | 3.72 ± 0.00 | 0.22 | Low |
| L. thermotolerans | 1.04 × 109 | 4.50 ± 0.00 | 3.69 ± 0.01 | 0.24 | |
| L. thermotolerans | 3.46 × 109 | 5.25 ± 1.06 | 3.76 ± 0.04 | 0.27 | |
| L. thermotolerans | 8.93 × 107 | 5.50 ± 0.71 | 3.65 ± 0.02 | 0.29 | |
| L. thermotolerans | 2.08 × 108 | 5.50 ± 0.71 | 3.95 ± 0.08 | 0.29 | |
| L. thermotolerans ** | 2.00 × 107 | 6.88 ± 2.25 | 4.51 ± 0.23 | 0.36 | Moderate |
| L. thermotolerans | 4.00 × 107 | 7.00 ± 0.82 | 4.45 ± 0.25 | 0.37 | |
| L. thermotolerans | 3.56 × 108 | 7.50 ± 0.71 | 3.66 ± 0.01 | 0.39 | |
| L. thermotolerans | 3.00 × 106 | 7.50 ± 1.29 | 3.86 ± 0.01 | 0.39 | |
| L. thermotolerans ** | 2.07 × 109 | 9.25 ±0.35 | 3.80 ± 0.00 | 0.48 | Similar to control |
| L. thermotolerans | 2.00 × 107 | 9.63± 2.36 | 3.87 ± 0.06 | 0.5 | |
| S. cerevisiae (control) | 9.00 × 107 | 9.52 ± 3.20 | 4.35 ± 0.39 | 0.5 | Control |
| Yeast | log CFU/mL | Ethanol% (v/v) | pH |
|---|---|---|---|
| S. cerevisiae (control) | 9.41 | 5.50 ± 0.32 | 5.02 ± 0.29 |
| L. thermotolerans | 9.36 | 5.67 ± 0.82 | 3.16 ± 0.03 |
| L. thermotolerans | 8.15 | 4.92 ± 0.36 | 4.12 ± 0.18 |
| L. quebecensis (OP923903) | 9.17 | 4.25 ± 0.45 | 3.07 ± 0.14 |
| L. thermotolerans (OP923897) | 9.00 | 4.50 ± 0.55 | 5.14 ± 0.49 |
| Parameters | Initial Values | L. thermotolerans | L. quebecencis |
|---|---|---|---|
| Color | 9.71 ± 1.01 | 9.83 ± 1.545 a | 9.73 ± 1.35 a |
| Bitterness | 58.7 ± 4.85 | 30.2 ± 4.72 a | 32.0 ± 3.06 a |
| Final Extract (%w/w) | 13.0 ± 0.78 | 6.3 ± 0.17 a | 7.19 ± 1.31 b |
| Original extract (%w/w) | 12.9 ± 0.76 | 12.5 ± 0.738 a | 12.1 ± 0.604 a |
| pH | 5.55 ± 0.01 | 4.18 ± 0.11 a | 4.06 ± 0.20 b |
| Ethanol (%v/v) | ND | 3.78 ± 0.49 a | 3.50 ± 0.46 b |
| Lactic acid (g/L) | ND | 1.25 ± 0.03 a | 1.21 ± 0.05 a |
| Glucose (g/L) | 11.9 ± 1.56 | ND | ND |
| Maltose (g/L) | 75.3 ± 1.23 | 2.75 ± 0.35 a | 2.81 ± 0.50 a |
| FAN (ppm) | 185.8 ± 5.27 | - | - |
| Volatile Composition (ppm) | L. thermotolerans | L. quebecensis | Odor Threshold (ppm) |
|---|---|---|---|
| Acetaldehyde | 16.9 ± 3.69 a | 14.9 ± 8.49 a | 20–10 |
| Dimethyl sulfur (DMS) | 65.3 ± 18.99 a | 47.2 ± 24.71 b | 25–50 |
| Ethyl acetate | 7.87 ± 1.06 a | 34.9 ± 11.87 b | 20–40 |
| 1-propanol | 26.5 ± 5.15 a | 21.6 ± 1.70 b | >700 |
| Isobutanol | 67.9 ± 10.91 a | 26.4 ± 2.04 b | >200 |
| Isoamyl acetate | 0.29 ± 0.05 a | 3.18 ± 1.37 b | NA |
| Isoamyl alcohol | 74.3 ± 7.55 a | 88.8 ± 6.78 b | >70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galaz, V.; Franco, W. Lachancea quebecensis a Novel Isolate for the Production of Craft Beer. Foods 2023, 12, 3347. https://doi.org/10.3390/foods12183347
Galaz V, Franco W. Lachancea quebecensis a Novel Isolate for the Production of Craft Beer. Foods. 2023; 12(18):3347. https://doi.org/10.3390/foods12183347
Chicago/Turabian StyleGalaz, Valeria, and Wendy Franco. 2023. "Lachancea quebecensis a Novel Isolate for the Production of Craft Beer" Foods 12, no. 18: 3347. https://doi.org/10.3390/foods12183347
APA StyleGalaz, V., & Franco, W. (2023). Lachancea quebecensis a Novel Isolate for the Production of Craft Beer. Foods, 12(18), 3347. https://doi.org/10.3390/foods12183347