
Fermentation of Betaphycus gelatinum Using Lactobacillus brevis: Growth of Probiotics, Total Polyphenol Content, Polyphenol Profile, and Antioxidant Capacity
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemicals and Reagents
2.3. Preparation of the Betaphycus Gelatinum Substrate for Fermentation
2.4. Preparation of the L. brevis Starter
2.5. Fermentation
2.6. Determination of the Viable Colony Number, pH, and Reducing Sugar Content
2.7. Extraction of Free and Bound Polyphenols
2.8. Determination of Total Phenolic Content (TPC)
2.9. Polyphenol Profile
2.10. Antioxidant Ability
2.11. Statistical Analyses
3. Results and Discussion
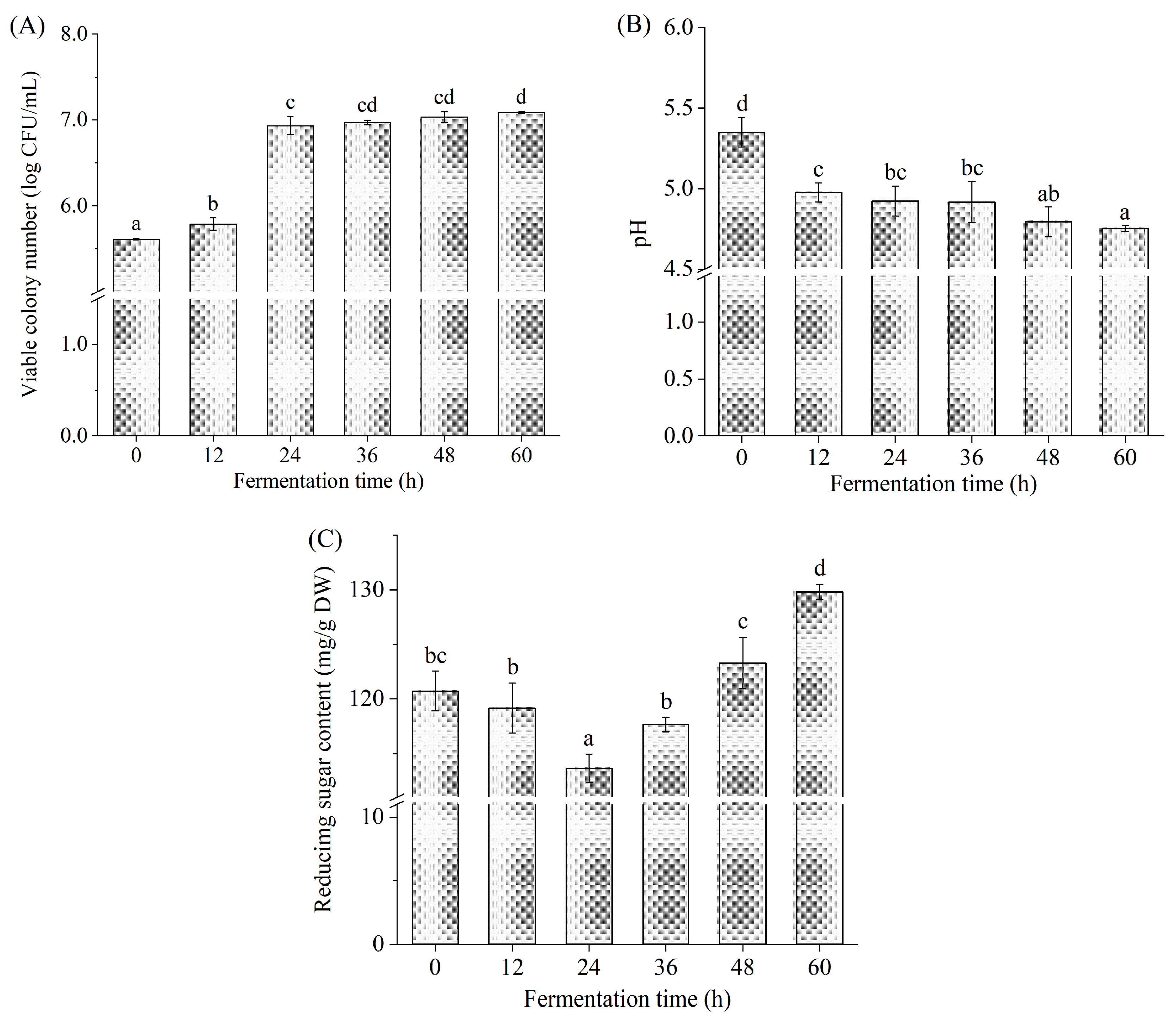
3.1. Viable Colony Number, pH, and Reducing Sugar Content
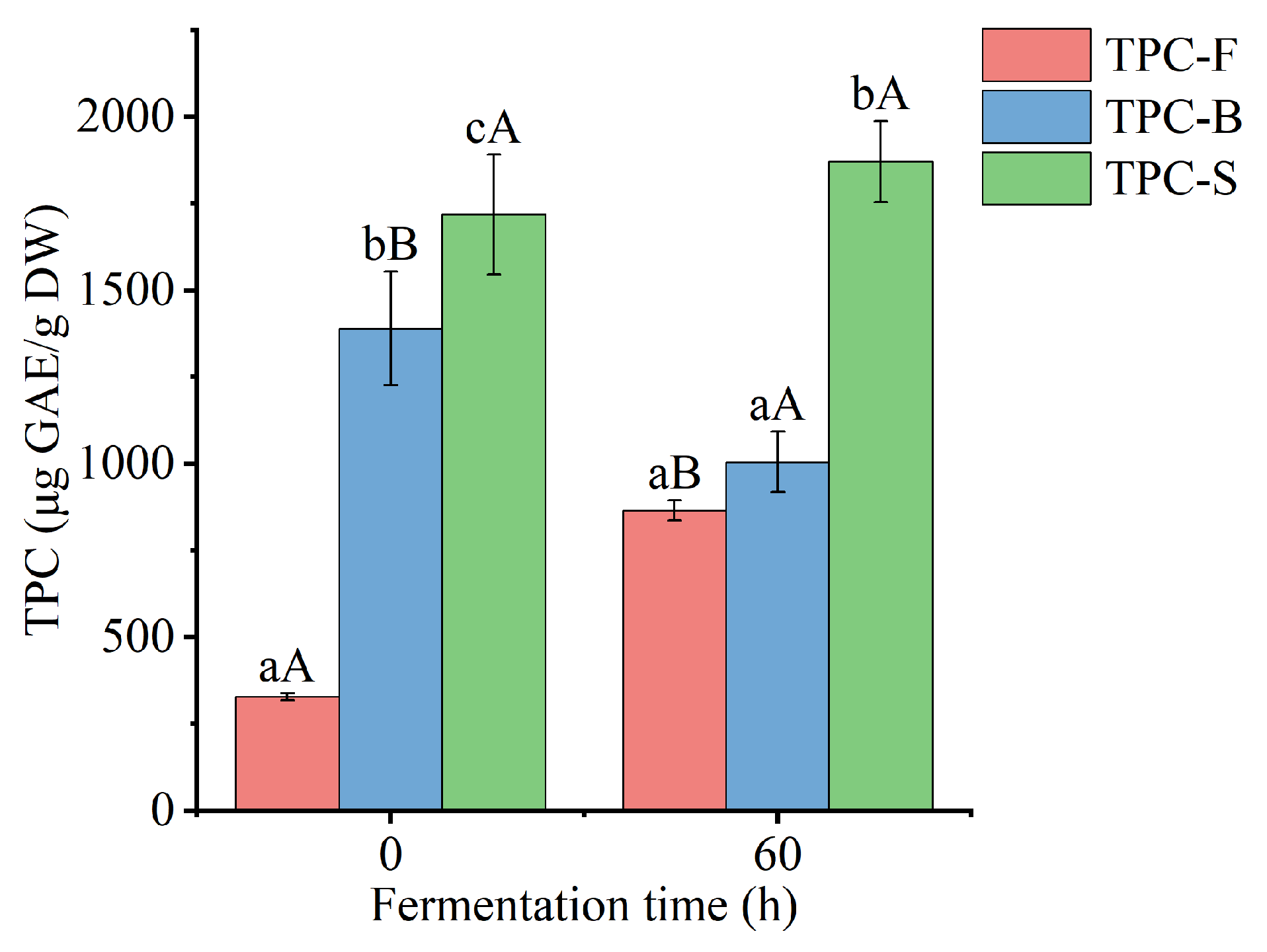
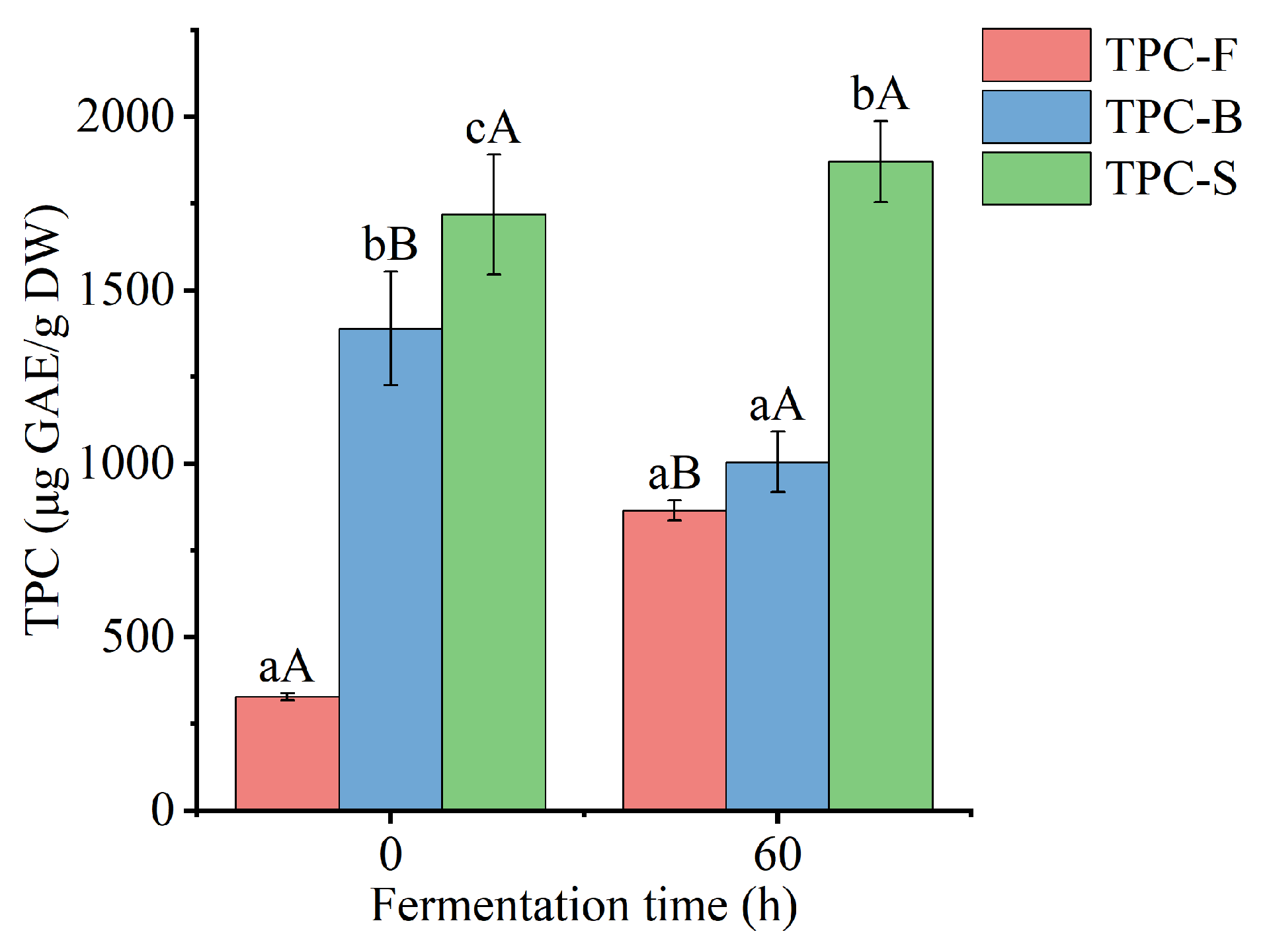
3.2. Free and Bound TPC
3.3. Polyphenol Compound Identification
3.4. Polyphenol Compound Quantification
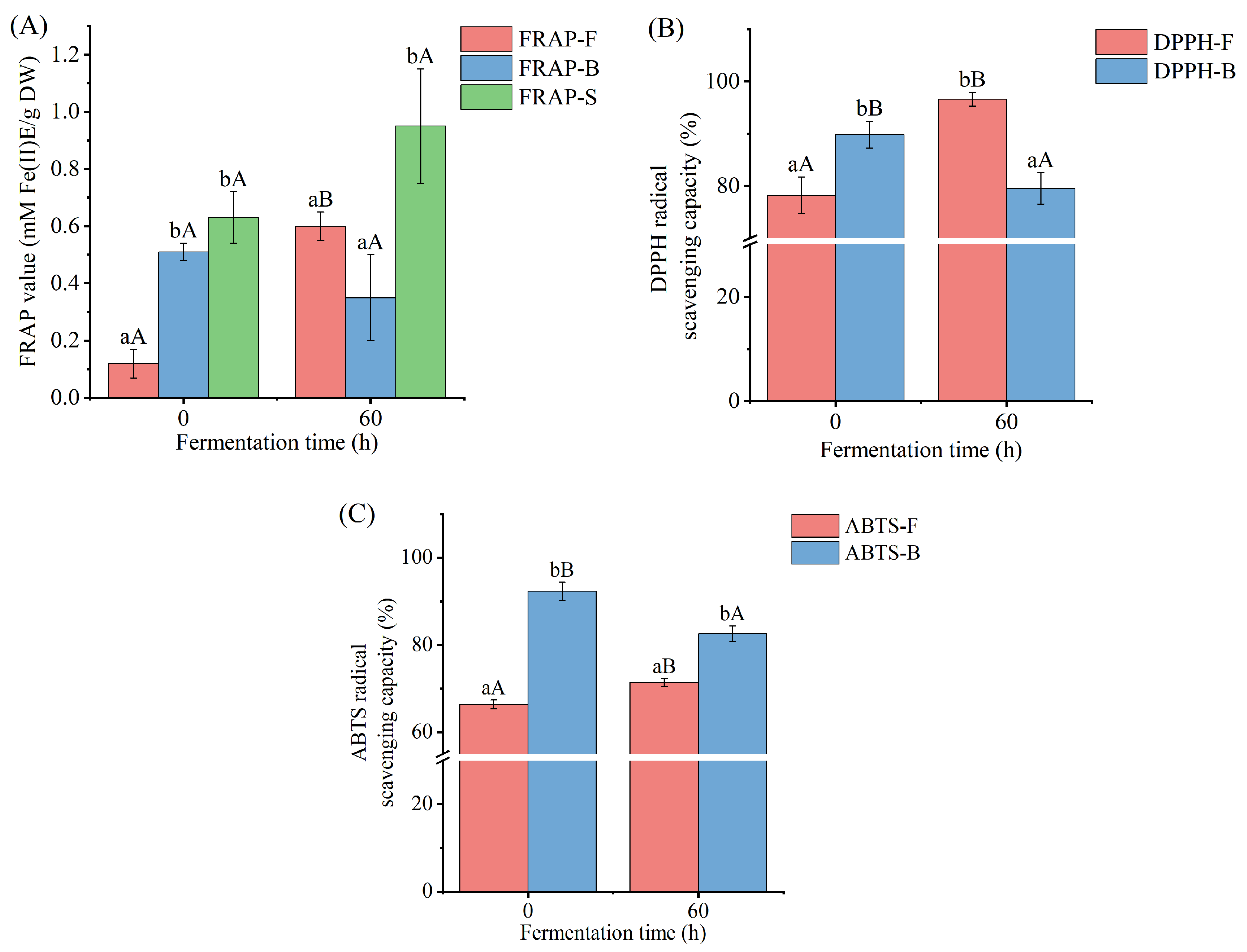
3.5. Antioxidant Activity
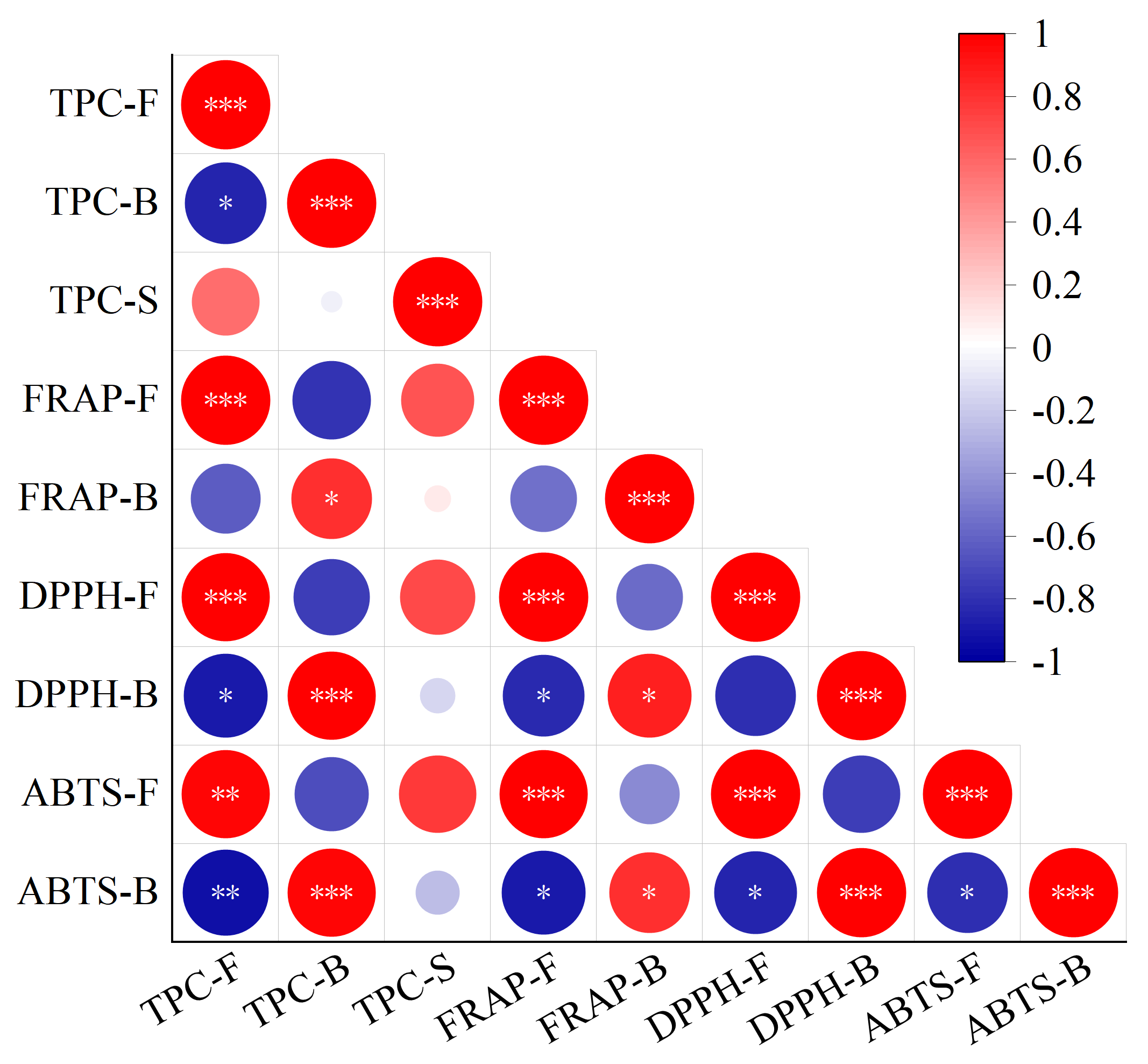
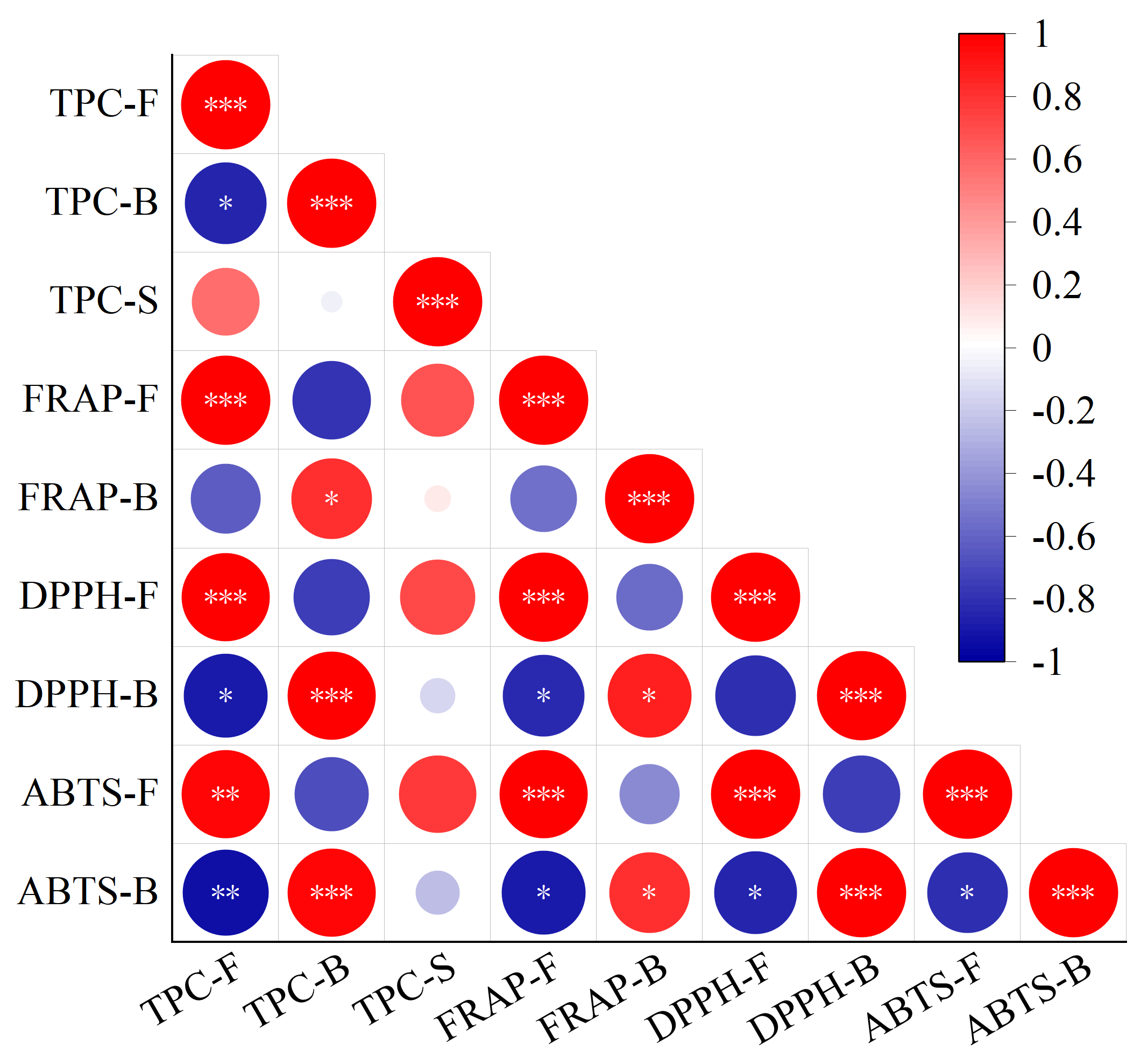
3.6. Correlation Analysis between TPC and Antioxidant Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Laboratory of Marine Biology of Institute of South China Sea Oceanography of Chinese Academy of Sciences. Marine Medicinal organisms in the South China Sea, 1st ed.; Science Press: Beijing, China, 1978; pp. 15–23. [Google Scholar]
- Pereira, L.; Critchley, A.T.; Amado, A.M.; Ribeiro-Claro, P.J.A. A comparative analysis of phycocolloids produced by underutilized versus industrially utilized carrageenophytes (Gigartinales, Rhodophyta). J. Appl. Phycol. 2009, 21, 599–605. [Google Scholar] [CrossRef]
- Zhu, J.; Fang, Z.; Liu, M.; Liang, L.; Huang, H.; Zou, X.; Lin, X.; Bao, S. Effects of temperature, salinity and light intensity on the growth, pigment and carrageenan of Betaphycus gelatinum. Algol. Stud. 2014, 145–146, 135–144. [Google Scholar] [CrossRef]
- Rudke, A.R.; da Silva, M.; Andrade, C.J.d.; Vitali, L.; Ferreira, S.R.S. Green extraction of phenolic compounds and carrageenan from the red alga Kappaphycus alvarezii. Algal Res. 2022, 67, 102866. [Google Scholar] [CrossRef]
- De Corato, U.; Salimbeni, R.; De Pretis, A.; Avella, N.; Patruno, G. Antifungal activity of crude extracts from brown and red seaweeds by a supercritical carbon dioxide technique against fruit postharvest fungal diseases. Postharvest Biol. Technol. 2017, 131, 16–30. [Google Scholar] [CrossRef]
- Namvar, F.; Mohamed, S.; Fard, S.G.; Behravan, J.; Mustapha, N.M.; Alitheen, N.B.M.; Othman, F. Polyphenol-rich seaweed (Eucheuma cottonii) extract suppresses breast tumour via hormone modulation and apoptosis induction. Food Chem. 2012, 130, 376–382. [Google Scholar] [CrossRef]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Wozniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Goncalves, A.M.M.; da Silva, G.J.; Pereira, L. Seaweed Phenolics: From extraction to applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, H.; Wang, Y.; Peng, Z.; Guo, Z.; Ma, Y.; Zhang, R.; Zhang, M.; Wu, Q.; Xiao, J.; et al. Effects of different extraction methods on contents, profiles, and antioxidant abilities of free and bound phenolics of Sargassum polycystum from the South China Sea. J. Food Sci. 2022, 87, 968–981. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, Y.; Li, H.; Deng, Z.; Tsao, R. A review on insoluble-bound phenolics in plant-based food matrix and their contribution to human health with future perspectives. Trends Food Sci. Technol. 2020, 105, 347–362. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.; Zhou, M.; Wu, Y.; Guan, X. Study on the interaction between grain polyphenols and intestinal microorganisms: A review. Food Biosci. 2023, 53, 102536. [Google Scholar] [CrossRef]
- Qiu, L.; Zhang, M.; Chang, L. Effects of lactic acid bacteria fermentation on the phytochemicals content, taste and aroma of blended edible rose and shiitake beverage. Food Chem. 2023, 405, 134722. [Google Scholar] [CrossRef]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of blueberry and blackberry juices using Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium bifidum: Growth of probiotics, metabolism of phenolics, antioxidant capacity in vitro and sensory evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, J.; Wang, Y.; Wang, X.; Ren, Y.; Yue, T.; Wang, Z.; Gao, Z. Study on the nutritional characteristics and antioxidant activity of dealcoholized sequentially fermented apple juice with Saccharomyces cerevisiae and Lactobacillus plantarum fermentation. Food Chem. 2021, 363, 130351. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Wang, L.; Li, S. Enhancement in the physicochemical properties, antioxidant activity, volatile compounds, and non-volatile compounds of watermelon juices through Lactobacillus plantarum JHT78 fermentation. Food Chem. 2023, 420, 136146. [Google Scholar] [CrossRef]
- Othman, N.B.; Roblain, D.; Chammen, N.; Thonart, P.; Hamdi, M. Antioxidant phenolic compounds loss during the fermentation of Chétoui olives. Food Chem. 2009, 116, 662–669. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N.; Scannell, A.G.M. Growth and kinetics of Lactobacillus plantarum in the fermentation of edible Irish brown seaweeds. Food Bioprod. Process. 2011, 89, 346–355. [Google Scholar] [CrossRef]
- Shobharani, P.; Halami, P.M.; Sachindra, N.M. Potential of marine lactic acid bacteria to ferment Sargassum sp. for enhanced anticoagulant and antioxidant properties. J. Appl. Microbiol. 2013, 114, 96–107. [Google Scholar] [CrossRef]
- Takei, M.; Kuda, T.; Eda, M.; Shikano, A.; Takahashi, H.; Kimura, B. Antioxidant and fermentation properties of aqueous solutions of dried algal products from the Boso Peninsula, Japan. Food Biosci. 2017, 19, 85–91. [Google Scholar] [CrossRef]
- Lee, S.-J.; Lee, D.-G.; Park, S.-H.; Kim, M.; Kong, C.-S.; Kim, Y.-Y.; Lee, S.-H. Comparison of biological activities in Sargassum siliquanstrum fermented by isolated lactic acid bacteria. Biotechnol. Bioprocess Eng. 2015, 20, 341–348. [Google Scholar] [CrossRef]
- Niccolai, A.; Shannon, E.; Abu-Ghannam, N.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Lactic acid fermentation of Arthrospira platensis (spirulina) biomass for probiotic-based products. J. Appl. Phycol. 2018, 31, 1077–1083. [Google Scholar] [CrossRef]
- Shin, N.R.; Wang, J.-H.; Lim, D.; Lee, M.-J.; Kim, H. Microbial change and fermentation characteristics during Samjung-Hwan natural fermentation. J. Korean Med. Obes. Res. 2015, 15, 123–130. [Google Scholar] [CrossRef]
- Chibuike, O.A.; Agwaranze, D.I.; Aliba, N.V.; Chukwuma, K.A.; Blessing, N.C. Fermentation by lactic acid bacteria consortium and its effect on anti-nutritional factors in maize flour. J. Biol. Sci. 2018, 19, 17–23. [Google Scholar] [CrossRef]
- Jeon, C.-P.; Lee, J.-G.; Lee, J.-B.; Park, S.-C.; Choi, C.-S.; Kim, J.-E.; Kwon, G.-S. Biological activities of fermented Dioscorea batatas dence by two stage fermentation. Korean J. Microbiol. 2012, 48, 29–36. [Google Scholar] [CrossRef]
- Yu, D.; Kwon, G.; Kim, M.; Kim, Y.M.; Choi, S.I.; Kim, G.H.; Chung, D. Moisture sorption characteristics of probiotic-fermented sea tangle powder and its thermodynamic properties. J. Food Process. Pres. 2019, 43, e13991. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, C.; Xiao, J. Study on Ultrasound-assisted mild acid hydrolysis of Betaphycus Gelatinum. Nat. Sci. J. Hainan Univ. 2023. accepted. [Google Scholar]
- Wang, D.; Wang, J.; Zeng, R.; Wu, J.; Michael, S.V.; Qu, W. The degradation activities for three seaweed polysaccharides of Shewanella sp. WPAGA9 isolated from deep-sea sediments. J. Basic. Microbiol. 2021, 61, 406–418. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, R.; Deng, Y.; Zhang, Y.; Xiao, J.; Huang, F.; Wen, W.; Zhang, M. Fermentation and complex enzyme hydrolysis enhance total phenolics and antioxidant activity of aqueous solution from rice bran pretreated by steaming with alpha-amylase. Food Chem. 2017, 221, 636–643. [Google Scholar] [CrossRef]
- Guo, W.; Chen, M.; Cui, S.; Tang, X.; Zhang, Q.; Zhao, J.; Mao, B.; Zhang, H. Dynamics changes in physicochemical properties, volatile metabolites, non-volatile metabolites, and physiological functions of barley juice during Bifidobacterium infantis fermentation. Food Chem. 2023, 407, 135201. [Google Scholar] [CrossRef]
- Aragon-Alegro, L.C.; Alarcon Alegro, J.H.; Roberta Cardarelli, H.; Chih Chiu, M.; Isay Saad, S.M. Potentially probiotic and synbiotic chocolate mousse. LWT-Food Sci. Technol. 2007, 40, 669–675. [Google Scholar] [CrossRef]
- Wang, W.; Yuan, Y.; Cao, J.; Shen, X.; Li, C. Beneficial effects of Holothuria leucospilota polysaccharides on fermentability in vivo and in vitro. Foods 2021, 10, 1884. [Google Scholar] [CrossRef]
- Zheng, X.; Yu, Y.; Xiao, G.; Xu, Y.; Wu, J.; Tang, D.; Cheng, Y.; Zhang, Y. Changes of anti-glucosidase content and some other characteristics in mulberry juice during fermentation with Leuconostoc mesenteroides. Acta Aliment. 2014, 43, 668–675. [Google Scholar] [CrossRef]
- Gao, H.; Wen, J.-J.; Hu, J.-L.; Nie, Q.-X.; Chen, H.-H.; Nie, S.-P.; Xiong, T.; Xie, M.-Y. Momordica charantia juice with Lactobacillus plantarum fermentation: Chemical composition, antioxidant properties and aroma profile. Food Biosci. 2019, 29, 62–72. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Li, X.; Wang, C.; Li, Q.; Xu, M.; Guan, X.; Lan, Z.; Ni, Y.; Zhang, Y. Widely targeted metabolomics analysis of enriched secondary metabolites and determination of their corresponding antioxidant activities in Elaeagnus angustifolia var. orientalis (L.) Kuntze fruit juice enhanced by Bifidobacterium animalis subsp. Lactis HN-3 fermentation. Food Chem. 2022, 374, 131568. [Google Scholar] [CrossRef] [PubMed]
- Arivalagan, M.; Roy, T.K.; Yasmeen, A.M.; Pavithra, K.C.; Jwala, P.N.; Shivasankara, K.S.; Manikantan, M.R.; Hebbar, K.B.; Kanade, S.R. Extraction of phenolic compounds with antioxidant potential from coconut (Cocos nucifera L.) testa and identification of phenolic acids and flavonoids using UPLC coupled with TQD-MS/MS. LWT 2018, 92, 116–126. [Google Scholar] [CrossRef]
- de Jager, L.S.; Perfetti, G.A.; Diachenko, G.W. Determination of coumarin, vanillin, and ethyl vanillin in vanilla extract products: Liquid chromatography mass spectrometry method development and validation studies. J. Chromatogr. A 2007, 1145, 83–88. [Google Scholar] [CrossRef]
- Lambert, M.; Meudec, E.; Verbaere, A.; Mazerolles, G.; Wirth, J.; Masson, G.; Cheynier, V.; Sommerer, N. A high-throughput UHPLC-QqQ-MS method for polyphenol profiling in rose wines. Molecules 2015, 20, 7890–7914. [Google Scholar] [CrossRef]
- Ren, M.; Xu, W.; Zhang, Y.; Ni, L.; Lin, Y.; Zhang, X.; Huang, M. Qualitative and quantitative analysis of phenolic compounds by UPLC-MS/MS and biological activities of Pholidota chinensis Lindl. J. Pharm. Biomed. Anal. 2020, 187, 113350. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Thadhani, V.M.; Ul Haq, F.; Khan, M.N.; Ali, S.; Musharraf, S.G. Rapid identification and quantification of bioactive metabolites in processed Camellia sinensis samples by UHPLC-ESI-MS/MS and evaluation of their antioxidant activity. J. Ind. Eng. Chem. 2020, 90, 419–426. [Google Scholar] [CrossRef]
- Fischer, U.A.; Carle, R.; Kammerer, D.R. Identification and quantification of phenolic compounds from pomegranate (Punica granatum L.) peel, mesocarp, aril and differently produced juices by HPLC-DAD-ESI/MS(n). Food Chem. 2011, 127, 807–821. [Google Scholar] [CrossRef]
- Justesen, U. Negative atmospheric pressure chemical ionisation low-energy collision activation mass spectrometry for the characterisation of flavonoids in extracts of fresh herbs. J. Chromatogr. A 2000, 902, 369–379. [Google Scholar] [CrossRef]
- Wang, X.; Xia, H.; Xing, F.; Deng, G.; Shen, Q.; Zeng, S. A highly sensitive and robust UPLC–MS with electrospray ionization method for quantitation of taxifolin in rat plasma. J. Chromatogr. B 2009, 877, 1778–1786. [Google Scholar] [CrossRef]
- Olate-Gallegos, C.; Barriga, A.; Vergara, C.; Fredes, C.; García, P.; Giménez, B.; Robert, P. Identification of polyphenols from Chilean brown seaweeds extracts by LC-DAD-ESI-MS/MS. J. Aquat. Food Prod. Technol. 2019, 28, 375–391. [Google Scholar] [CrossRef]
- Xin, Z.; Ma, S.; Ren, D.; Liu, W.; Han, B.; Zhang, Y.; Xiao, J.; Yi, L.; Deng, B. UPLC-Orbitrap-MS/MS combined with chemometrics establishes variations in chemical components in green tea from Yunnan and Hunan origins. Food Chem. 2018, 266, 534–544. [Google Scholar] [CrossRef]
- Chen, P.X.; Tang, Y.; Marcone, M.F.; Pauls, P.K.; Zhang, B.; Liu, R.; Tsao, R. Characterization of free, conjugated and bound phenolics and lipophilic antioxidants in regular- and non-darkening cranberry beans (Phaseolus vulgaris L.). Food Chem. 2015, 185, 298–308. [Google Scholar] [CrossRef]
- Gao, H.; Wang, Y.; Guo, Z.; Liu, Y.; Wu, Q.; Xiao, J. Optimization of ultrasound-assisted extraction of phenolics from Asparagopsis taxiformis with deep eutectic solvent and their characterization by ultra-high-performance liquid chromatography-mass spectrometry. Front. Nutr. 2022, 9, 1036436. [Google Scholar] [CrossRef]
- Chung, H.Y.; Ma, W.C.J.; Ang, P.O.; Kim, J.-S.; Chen, F. Seasonal variations of bromophenols in brown algae (Padina arborescens, Sargassum siliquastrum, and Lobophora variegata) collected in Hong Kong. J. Agric. Food Chem. 2003, 51, 2619–2624. [Google Scholar] [CrossRef]
- Generalic Mekinic, I.; Skroza, D.; Simat, V.; Hamed, I.; Cagalj, M.; Popovic Perkovic, Z. Phenolic content of brown algae (Pheophyceae) species: Extraction, identification, and quantification. Biomolecules 2019, 9, 244. [Google Scholar] [CrossRef]
- Yoshie-Stark, Y.; Hsieh, Y.P.; Suzuki, T. Distribution of flavonoids and related compounds from seaweeds in Japan. J. Tokyo Univ. Fish. 2003, 89, 1–6. [Google Scholar]
- Zhang, J.; Huang, X.; Cheng, J.; Wang, C. Effect of Lactobacillus (L. acidophilus NCIB1899, L. casei CRL 431, L. paracasei LP33) fermentation on free and bound polyphenolic, antioxidant activities in three Chenopodium quinoa cultivars. J. Food Sci. 2023, 88, 2679–2692. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Cirlini, M.; Calani, L.; Bernini, V.; Neviani, E.; Del Rio, D.; Galaverna, G.; Lazzi, C. In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria. Food Chem. 2019, 276, 692–699. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Devlamynck, T.; Te Poele, E.M.; Meng, X.; van Leeuwen, S.S.; Dijkhuizen, L. Glucansucrase Gtf180-ΔN of Lactobacillus reuteri 180: Enzyme and reaction engineering for improved glycosylation of non-carbohydrate molecules. Appl. Microbiol. Biotechnol. 2016, 100, 7529–7539. [Google Scholar] [CrossRef]
- Li, X.; Ouyang, X.; Cai, R.; Chen, D. 3′,8″-Dimerization enhances the antioxidant capacity of flavonoids: Evidence from acacetin and isoginkgetin. Molecules 2019, 24, 2039. [Google Scholar] [CrossRef] [PubMed]
- Scherer, R.; Godoy, H.T. Antioxidant activity index (AAI) by the 2,2-diphenyl-1-picrylhydrazyl method. Food Chem. 2009, 112, 654–658. [Google Scholar] [CrossRef]
- Jones, A.; Pravadali-Cekic, S.; Dennis, G.R.; Bashir, R.; Mahon, P.J.; Shalliker, R.A. Ferric reducing antioxidant potential (FRAP) of antioxidants using reaction flow chromatography. Anal. Chim. Acta 2017, 967, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Hu, C.Y.; Chen, Y.H.; Li, Y.T.; Chung, Y.C. Submerged fermentation with Lactobacillus brevis significantly improved the physiological activities of Citrus aurantium flower extract. Heliyon 2022, 8, e10498. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D.; Waśko, A.; Targoński, Z.; Niedźwiadek, M.; Bancarzewska, M. The use of a novel ferulic acid esterase from Lactobacillus acidophilus K1 for the release of phenolic acids from brewer’s spent grain. J. Inst. Brew. 2010, 116, 293–303. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Tentative Assignment | ESI Model | Parent Ions | Fragment Ions | Reference |
|---|---|---|---|---|---|
| 1 | 4-hydroxybenzoic acid | − | 137.00 | 93.00 | [35], standard |
| 2 | ethyl vanillin | − | 164.95 | 136.21, 92.05 | [36], standard |
| 3 | trans-cinnamic acid | + | 146.95 | 118.94, 77.01, 40.07 | [35], standard |
| 4 | caffeic acid | − | 179.00 | 135.00, 79.00 | [37], standard |
| 5 | ferulic acid | − | 193.00 | 178.00, 149.00, 134.00 | [37], standard |
| 6 | rosmarinic acid | + | 358.96 | 196.96, 161.00 | [4], standard |
| 7 | 1,3-O-dicaffeoylquinic acid | − | 514.91 | 352.94, 191.01 | [38], standard |
| 8 | gallocatechin | − | 304.98 | 179.01, 124.98 | [39], standard |
| 9 | epigallocatechin | − | 304.98 | 179.01, 124.98 | [39], standard |
| 10 | epicatechin gallate | − | 440.89 | 288.99, 168.98, 124.97 | [39], standard |
| 11 | ellagic acid | − | 301.08 | 258.07, 229.10 | [40], standard |
| 12 | quercetin | − | 301.01 | 178.99, 150.99 | [35], standard |
| 13 | morin | − | 300.83 | 150.99, 124.98 106.97 | [41], standard |
| 14 | myricetin | − | 317.00 | 178.95, 150.99 | [37], standard |
| 15 | taxifolin | − | 302.83 | 284.98, 176.91, 124.98 | [42], standard |
| 16 | hinokiflavone | + | 537.00 | 417.00, 284.00 | [9], standard |
| 17 | hyperoside | − | 463.00 | 300.90, 270.90, 254.90 | [43], standard |
| 18 | isoquercitrin | − | 447.00 | 301.00, 179.00, 151.00 | [43], standard |
| 19 | kaempferol-3-O-glucoside | − | 446.90 | 285.06, 254.94, 226.98 | [43], standard |
| 20 | cosemetin | − | 430.90 | 268.90 | [43], standard |
| 21 | vitexin | − | 431.00 | 310.90, 340.80, 282.90 | [44], standard |
| 22 | myricitrin | − | 462.90 | 316.99 | [43], standard |
| 23 | taxifolin-7-O-rhamnoside | − | 448.85 | 302.92, 284.91, 124.98 | [43], standard |
| 24 | isorhamnetin-3-O-glucoside | − | 476.86 | 313.96, 242.84 | [43], standard |
| 25 | cyanidin | + | 286.91 | 109.05, 137.01 | [45], standard |
| 26 | 2,4-dibromophenol | − | 250.74 | 168.70, 81.37, 78.51 | [46], standard |
| 27 | 2,4,6-tribromophenol | − | 330.60 | 81.35 | [47], standard |
| Free Polyphenol Content (μg/g DW) | Bound Polyphenol Content (μg/g DW) | |||
|---|---|---|---|---|
| 0 h | 60 h | 0 h | 60 h | |
| 4-hydroxybenzoic acid | 1.64 ± 0.02 a | 1.61 ± 0.02 a | 1.84 ± 0.04 A | 1.79 ± 0.03 A |
| ethyl vanillin | 24.05 ± 0.70 a | 48.91 ± 1.51 b | 11.65 ± 0.31 B | 2.23 ± 0.03 A |
| trans-cinnamic acid | 84.49 ± 3.47 b | 77.16 ± 0.47 a | 1244.25 ± 2.66 B | 195.74 ± 5.51 A |
| caffeic acid | nd | 1.60 ± 0.01 a | 1.63 ± 0.02 A | 1.67 ± 0.03 A |
| ferulic acid | 19.94 ± 0.56 a | nd | 16.74 ± 0.67 B | 12.81 ± 0.32 A |
| rosmarinic acid | nd | 560.94 ± 21.56 a | 583.75 ± 1.25 B | 171.77 ± 4.90 A |
| 1,3-O-dicaffeoylquinic acid | 2.39 ± 0.002 a | 3.17 ± 0.16 b | 2.49 ± 0.05 A | 2.38 ± 0.08 A |
| number of phenolic acids | 5 | 6 | 7 | 7 |
| sum of individul phenolic acid content | 132.51 ± 4.75 a | 693.40 ± 23.72 b | 1862.34 ± 4.99 B | 388.38 ± 10.89 A |
| gallocatechin | 4.25 ± 0.04 a | 8.76 ± 0.26 b | 9.44 ± 0.27 B | 8.96 ± 0.08 A |
| epigallocatechin | nd | 4.73 ± 0.19 a | 4.23 ± 0.06 A | 4.38 ± 0.20 A |
| epicatechin gallate | 3.51 ± 0.15 b | 2.39 ± 0.08 a | 3.71 ± 0.01 B | 2.42 ± 0.08 A |
| ellagic acid | 2.71 ± 0.05 a | 4.39 ± 0.12 b | 4.63 ± 0.15 A | 4.53 ± 0.03 A |
| quercetin | 1.62 ± 0.05 a | 1.60 ± 0.05 a | 3.09 ± 0.01 B | 1.65 ± 0.004 A |
| morin | 1.19 ± 0.03 a | 1.15 ± 0.002 a | 1.17 ± 0.004 A | nd |
| myricetin | 2.28 ± 0.07 a | nd | nd | nd |
| taxifolin | 1.13 ± 0.01 a | 1.14 ± 0.002 a | 1.13 ± 0.01 A | 1.13 ± 0.004 A |
| hinokiflavone | 42.50 ± 0.42 b | 39.33 ± 0.54 a | 49.03 ± 1.40 B | 43.02 ± 1.35 A |
| hyperoside | nd | 0.56 ± 0.002 a | 0.58 ± 0.01 A | 0.60 ± 0.03 A |
| isoquercitrin | nd | 0.67 ± 0.01 a | 0.68 ± 0.02 A | nd |
| kaempferol-3-O-glucoside | 1.18 ± 0.01 a | nd | nd | nd |
| cosemetin | nd | 0.81 ± 0.002 a | nd | nd |
| vitexin | 3.06 ± 0.10 a | nd | 3.46 ± 0.10 A | 3.31 ± 0.15 A |
| myricitrin | 3.24 ± 0.09 a | 3.22 ± 0.09 a | 3.40 ± 0.04 B | 3.21 ± 0.08 A |
| taxifolin-7-O-rhamnoside | nd | 1.50 ± 0.002 a | 1.50 ± 0.002 A | nd |
| isorhamnetin-3-O-glucoside | 3.44 ± 0.004 b | 1.97 ± 0.08 a | 3.35 ± 0.09 B | 1.97 ± 0.09 A |
| cyanidin | 2.47 ± 0.09 b | 2.01 ± 0.06 a | 7.28 ± 0.22 B | 1.83 ± 0.004 A |
| number of flavonoids | 13 | 15 | 15 | 12 |
| sum of individual flavonoid content | 72.58 ± 1.11 a | 74.22 ± 1.48 a | 96.68 ± 2.39 B | 77.00 ± 2.09 A |
| 2,4-dibromophenol | nd | 32.91 ± 0.89 a | 24.83 ± 0.88 A | nd |
| 2,4,6-tribromophenol | 1.40 ± 0.06 a | 21.67 ± 0.62 b | 2.27 ± 0.06 B | 1.13 ± 0.04 A |
| number of bromophenols | 1 | 2 | 2 | 1 |
| sum of individual bromophenol content | 1.40 ± 0.06 a | 54.57 ± 1.50 b | 27.11 ± 0.94 B | 1.13 ± 0.04 A |
| number of polyphenols | 19 | 23 | 24 | 20 |
| sum of individual polyphenol content | 206.48 ± 5.92 a | 822.19 ± 26.70 b | 1986.13 ± 8.32 B | 466.51 ± 13.02 A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhao, C.; Guo, Z.; Li, S.; Zhu, Z.; Grimi, N.; Xiao, J. Fermentation of Betaphycus gelatinum Using Lactobacillus brevis: Growth of Probiotics, Total Polyphenol Content, Polyphenol Profile, and Antioxidant Capacity. Foods 2023, 12, 3334. https://doi.org/10.3390/foods12183334
Wang Z, Zhao C, Guo Z, Li S, Zhu Z, Grimi N, Xiao J. Fermentation of Betaphycus gelatinum Using Lactobacillus brevis: Growth of Probiotics, Total Polyphenol Content, Polyphenol Profile, and Antioxidant Capacity. Foods. 2023; 12(18):3334. https://doi.org/10.3390/foods12183334
Chicago/Turabian StyleWang, Zhe, Caibo Zhao, Zhiqiang Guo, Shuyi Li, Zhenzhou Zhu, Nabil Grimi, and Juan Xiao. 2023. "Fermentation of Betaphycus gelatinum Using Lactobacillus brevis: Growth of Probiotics, Total Polyphenol Content, Polyphenol Profile, and Antioxidant Capacity" Foods 12, no. 18: 3334. https://doi.org/10.3390/foods12183334
APA StyleWang, Z., Zhao, C., Guo, Z., Li, S., Zhu, Z., Grimi, N., & Xiao, J. (2023). Fermentation of Betaphycus gelatinum Using Lactobacillus brevis: Growth of Probiotics, Total Polyphenol Content, Polyphenol Profile, and Antioxidant Capacity. Foods, 12(18), 3334. https://doi.org/10.3390/foods12183334