
CodY: An Essential Transcriptional Regulator Involved in Environmental Stress Tolerance in Foodborne Staphylococcus aureus RMSA24


Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Growth Condition
2.2. Construction of codY Complementary Mutant
2.3. Growth Curve Determination
2.4. Heat Stress Assay
2.5. Cold Stress Assay
2.6. Desiccation Stress Assay
2.7. H2O2 Stress Assay
2.8. Salt Stress Assay
2.9. Reverse Transcriptase Quantitative PCR Experiment
2.10. Statistical Analysis
3. Results
3.1. Mutation of codY Had No Significant Effect on RMSA24 Growth
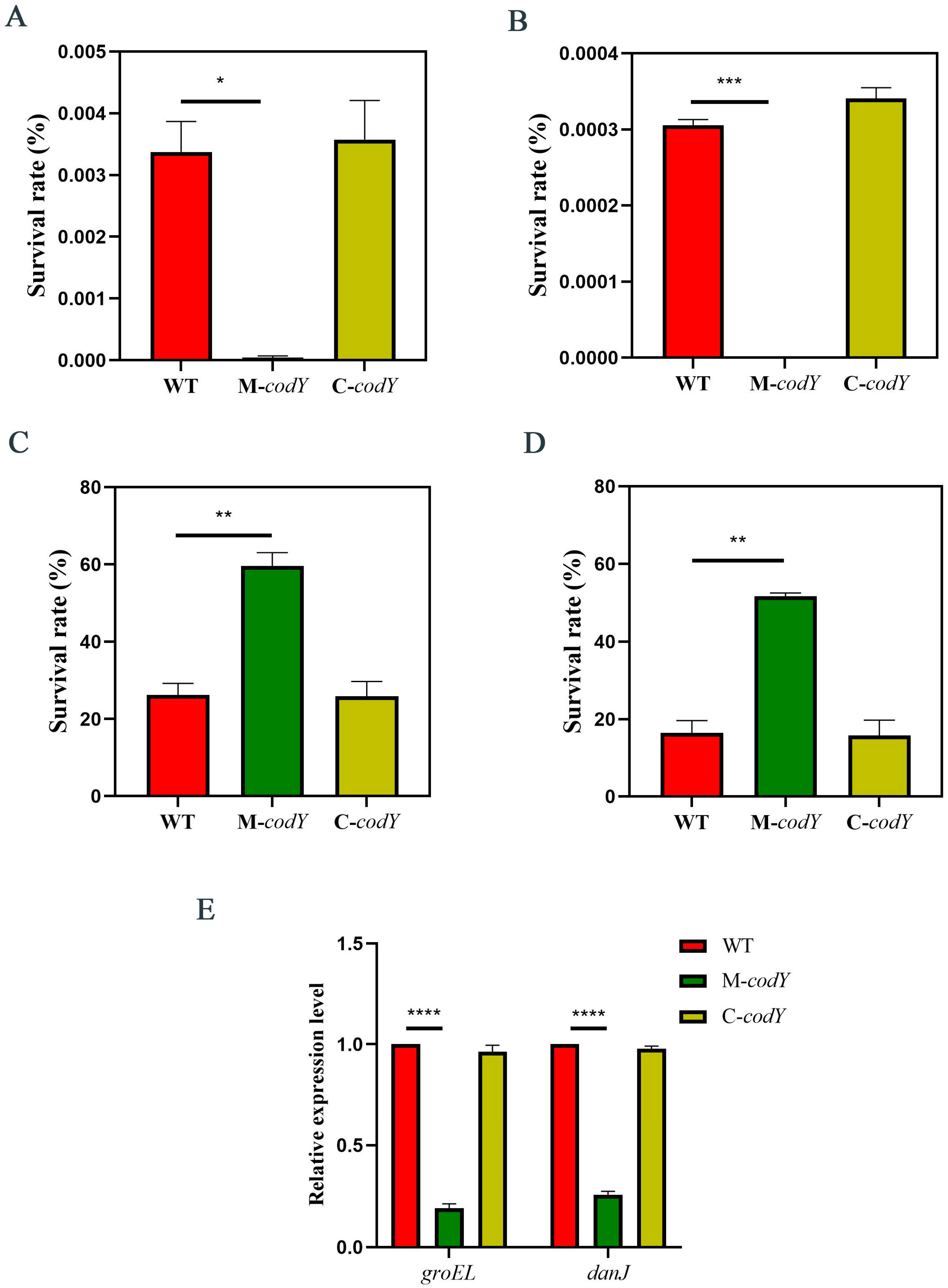
3.2. CodY Positively Regulates the Desiccation Tolerance of RMSA24
3.3. CodY Is Involved in Regulating the Temperature Stress Response of RMSA24
3.4. CodY Is Involved in Regulating the Oxidative Stress Response of RMSA24
3.5. CodY Mutation Increases the Sensitivity of RMSA24 to Salt
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dayan, G.H.; Mohamed, N.; Scully, I.L.; Cooper, D.; Begier, E.; Eiden, J.; Jansen, K.U.; Gurtman, A.; Anderson, A.S. Staphylococcus aureus: The current state of disease, pathophysiology and strategies for prevention. Expert Rev. Vaccines 2016, 15, 1373–1392. [Google Scholar] [CrossRef]
- Peacock, S.J.; de Silva, I.; Lowy, F.D. What determines nasal carriage of Staphylococcus aureus? Trends Microbiol. 2001, 9, 605–610. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus Infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal Enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef]
- Visioli, F.; Strata, A. Milk, Dairy Products, and Their Functional Effects in Humans: A Narrative Review of Recent Evidence. Adv. Nutr. Int. Rev. J. 2014, 5, 131–143. [Google Scholar] [CrossRef]
- Kim, N.H.; Cho, T.J.; Rhee, M.S. Chapter One-Sodium Chloride Does Not Ensure Microbiological Safety of Foods: Cases and Solutions. In Advances in Applied Microbiology; Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, UK, 2017; Volume 101, pp. 1–47. [Google Scholar]
- Lupín, H.M.; Boeri, R.L.; Moschiar, S.M. Water activity and salt content relationship in moist salted fish products. Int. J. Food Sci. Technol. 1981, 16, 31–38. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Wijtzes, T.; Rombouts, F.M.; Riet, K.V. A decision support system for prediction of microbial spoilage in foods. J. Ind. Microbiol. Biotechnol. 1993, 12, 324–329. [Google Scholar] [CrossRef]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and Staphylococcal Food-Borne Disease: An Ongoing Challenge in Public Health. BioMed Res. Int. 2014, 2014, 827965. [Google Scholar] [CrossRef]
- Chaibenjawong, P.; Foster, S.J. Desiccation tolerance in Staphylococcus aureus. Arch. Microbiol. 2011, 193, 125–135. [Google Scholar] [CrossRef]
- Neely, A.N.; Maley, M.P. Survival of Enterococci and Staphylococci on Hospital Fabrics and Plastic. J. Clin. Microbiol. 2000, 38, 724–726. [Google Scholar] [CrossRef] [PubMed]
- Abadias, M.; Benabarre, A.; Teixidó, N.; Usall, J.; Viñas, I. Effect of freeze drying and protectants on viability of the biocontrol yeast Candida sake. Int. J. Food Microbiol. 2001, 65, 173–182. [Google Scholar] [CrossRef]
- Cosgrove, K.; Coutts, G.; Jonsson, I.-M.; Tarkowski, A.; Kokai-Kun, J.F.; Mond, J.J.; Foster, S.J. Catalase (KatA) and Alkyl Hydroperoxide Reductase (AhpC) Have Compensatory Roles in Peroxide Stress Resistance and Are Required for Survival, Persistence, and Nasal Colonization in Staphylococcus aureus. J. Bacteriol. 2007, 189, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Linzner, N.; Fritsch, V.N.; Busche, T.; Tung, Q.N.; Van Loi, V.; Bernhardt, J.; Kalinowski, J.; Antelmann, H. The plant-derived naphthoquinone lapachol causes an oxidative stress response in Staphylococcus aureus. Free. Radic. Biol. Med. 2020, 158, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Chandrangsu, P.; Van Loi, V.; Antelmann, H.; Helmann, J.D. The Role of Bacillithiol in Gram-Positive Firmicutes. Antioxid. Redox Signal. 2017, 28, 445–462. [Google Scholar] [CrossRef]
- Christian, J.H.B.; Waltho, J.A. The Sodium and Potassium Content of Non-Halophilic Bacteria in Relation to Salt Tolerance. J. Gen. Microbiol. 1961, 25, 97–102. [Google Scholar] [CrossRef]
- Freeman, Z.N.; Dorus, S.; Waterfield, N.R. The KdpD/KdpE Two-Component System: Integrating K+ Homeostasis and Virulence. PLoS Pathog. 2013, 9, e1003201. [Google Scholar] [CrossRef] [PubMed]
- Price-Whelan, A.; Poon Chun, K.; Benson Meredith, A.; Eidem Tess, T.; Roux Christelle, M.; Boyd Jeffrey, M.; Dunman Paul, M.; Torres Victor, J.; Krulwich Terry, A. Transcriptional Profiling of Staphylococcus aureus During Growth in 2 M NaCl Leads to Clarification of Physiological Roles for Kdp and Ktr K+ Uptake Systems. mBio 2013, 4, e00407-13. [Google Scholar] [CrossRef]
- Brandon, L.; Dorus, S.; Epstein, W.; Altendorf, K.; Jung, K. Modulation of KdpD phosphatase implicated in the physiological expression of the Kdp ATPase of Escherichia coli. Mol. Microbiol. 2000, 38, 1086–1092. [Google Scholar] [CrossRef]
- Sugiura, A.; Nakashima, K.; Mizuno, T. Sequence-directed DNA Curvature in Activator-binding Sequence in the Escherichia coli kdpABC Promoter. Biosci. Biotechnol. Biochem. 1993, 57, 356–357. [Google Scholar] [CrossRef]
- Voelkner, P.; Puppe, W.; Altendorf, K. Characterization of the KdpD protein, the sensor kinase of the K+-translocating Kdp system of Escherichia coli. JBIC J. Biol. Inorg. Chem. 1993, 217, 1019–1026. [Google Scholar] [CrossRef]
- Tran, H.T.; Bonilla, C.Y. SigB-regulated antioxidant functions in gram-positive bacteria. World J. Microbiol. Biotechnol. 2021, 37, 38. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pförtner, H.; Niemann, S.; Geraci, J.; Van De Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma Factor SigB Is Crucial to Mediate Staphylococcus aureus Adaptation during Chronic Infections. PLoS Pathog. 2015, 11, e1004870. [Google Scholar] [CrossRef] [PubMed]
- Wemekamp-Kamphuis, H.H.; Wouters, J.A.; de Leeuw, P.P.L.A.; Hain, T.; Chakraborty, T.; Abee, T. Identification of Sigma Factor σ B -Controlled Genes and Their Impact on Acid Stress, High Hydrostatic Pressure, and Freeze Survival in Listeria monocytogenes EGD-e. Appl. Environ. Microbiol. 2004, 70, 3457–3466. [Google Scholar] [CrossRef] [PubMed]
- Hecker, M.; Pané-Farré, J.; Uwe, V. SigB-Dependent General Stress Response in Bacillus subtilis and Related Gram-Positive Bacteria. Annu. Rev. Microbiol. 2007, 61, 215–236. [Google Scholar] [CrossRef]
- Reeves, A.; Gerth, U.; Völker, U.; Haldenwang, W.G. ClpP Modulates the Activity of the Bacillus subtilis Stress Response Transcription Factor, σ B. J. Bacteriol. 2007, 189, 6168–6175. [Google Scholar] [CrossRef]
- Schulthess, B.; Bloes, D.A.; François, P.; Girard, M.; Schrenzel, J.; Bischoff, M.; Berger-Bächi, B. The σB-Dependent yabJ-spoVG Operon Is Involved in the Regulation of Extracellular Nuclease, Lipase, and Protease Expression in Staphylococcus aureus. J. Bacteriol. 2011, 193, 4954–4962. [Google Scholar] [CrossRef]
- Wang, H.; Hou, X.; Shen, J.; Wang, W.; Ye, Y.; Yu, J.; Xue, T. Alternative sigma factor B reduces biofilm formation and stress response in milk-derived Staphylococcus aureus. LWT 2022, 162, 113515. [Google Scholar] [CrossRef]
- Blevins, J.S.; Gillaspy, A.F.; Rechtin, T.M.; Hurlburt, B.K.; Smeltzer, M.S. The staphylococcal accessory regulator (sar) represses transcription of the Staphylococcus aureus collagen adhesin gene (cna ) in an agr-independent manner. Mol. Microbiol. 1999, 33, 317–326. [Google Scholar] [CrossRef]
- Cheung, A.L.; Koomey, J.M.; Butler, C.A.; Projan, S.J.; Fischetti, V.A. Regulation of exoprotein expression in Staphylococcus aureus by a locus (sar) distinct from agr. Proc. Natl. Acad. Sci. USA 1992, 89, 6462–6466. [Google Scholar] [CrossRef]
- Ballal, A.; Manna, A.C. Regulation of Superoxide Dismutase (sod) Genes by SarA in Staphylococcus aureus. J. Bacteriol. 2009, 191, 3301–3310. [Google Scholar] [CrossRef]
- Alonzo, F., 3rd; Torres, V.J. The Bicomponent Pore-Forming Leucocidins of Staphylococcus aureus. Microbiol. Mol. Biol. Rev. 2014, 78, 199–230. [Google Scholar] [CrossRef] [PubMed]
- Brinsmade, S.R. CodY, a master integrator of metabolism and virulence in Gram-positive bacteria. Curr. Genet. 2016, 63, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Belitsky, B.R. Indirect Repression by Bacillussubtilis CodY via Displacement of the Activator of the Proline Utilization Operon. J. Mol. Biol. 2011, 413, 321–336. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Belitsky, B.R.; Sonenshein, A.L. Roadblock Repression of Transcription by Bacillus subtilis CodY. J. Mol. Biol. 2011, 411, 729–743. [Google Scholar] [CrossRef]
- Chapeton-Montes, D.; Plourde, L.; Deneve, C.; Garnier, D.; Barbirato, F.; Colombié, V.; Demay, S.; Haustant, G.; Gorgette, O.; Schmitt, C.; et al. Tetanus Toxin Synthesis is Under the Control of A Complex Network of Regulatory Genes in Clostridium tetani. Toxins 2020, 12, 328. [Google Scholar] [CrossRef]
- Ababneh, Q.O.; Herman, J.K. CodY Regulates SigD Levels and Activity by Binding to Three Sites in the fla/che Operon. J. Bacteriol. 2015, 197, 2999–3006. [Google Scholar] [CrossRef]
- Brinsmade, S.R.; Alexander, E.L.; Livny, J.; Stettner, A.I.; Segrè, D.; Rhee, K.Y.; Sonenshein, A.L. Hierarchical expression of genes controlled by the Bacillus subtilis global regulatory protein CodY. Proc. Natl. Acad. Sci. USA 2014, 111, 8227–8232. [Google Scholar] [CrossRef] [PubMed]
- Pohl, K.; Francois, P.; Stenz, L.; Schlink, F.; Geiger, T.; Herbert, S.; Goerke, C.; Schrenzel, J.; Wolz, C. CodY in Staphylococcus aureus: A Regulatory Link between Metabolism and Virulence Gene Expression. J. Bacteriol. 2009, 191, 2953–2963. [Google Scholar] [CrossRef]
- Petranovic, D.; Guédon, E.; Sperandio, B.; Delorme, C.; Ehrlich, D.; Renault, P. Intracellular effectors regulating the activity of the Lactococcus lactis CodY pleiotropic transcription regulator. Mol. Microbiol. 2004, 53, 613–621. [Google Scholar] [CrossRef]
- Shivers, R.P.; Sonenshein, A.L. Activation of the Bacillus subtilis global regulator CodY by direct interaction with branched-chain amino acids. Mol. Microbiol. 2004, 53, 599–611. [Google Scholar] [CrossRef]
- Wang, Y.; He, H.; Li, H.; Lu, W.; Guo, T.; Kong, J. The global regulator CodY responds to oxidative stress by the regulation of glutathione biosynthesis in Streptococcus thermophilus. J. Dairy Sci. 2017, 100, 8768–8775. [Google Scholar] [CrossRef]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef]
- Richter, K.; Haslbeck, M.; Buchner, J. The Heat Shock Response: Life on the Verge of Death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Arsène, F.; Tomoyasu, T.; Bukau, B. The heat shock response of Escherichia coli. Int. J. Food Microbiol. 2000, 55, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.-B.; Shao, Y.-M.; Miao, S.; Wang, L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell. Mol. Life Sci. 2006, 63, 2560–2570. [Google Scholar] [CrossRef]
- Wang, H.; Ma, K.; Shen, J.; Fang, M.; Pei, H.; Li, Y.; Zhu, C.; Shu, F.; Li, B.; Xue, T. Genes associated with desiccation stress in foodborne Staphylococcus aureus as revealed by transposon insertion mutagenesis. Food Res. Int. 2023, 163, 112271. [Google Scholar] [CrossRef]
- Shen, J.; Wang, H.; Zhu, C.; Zhang, M.; Shang, F.; Xue, T. Effect of biofilm on the survival of Staphylococcus aureus isolated from raw milk in high temperature and drying environment. Food Res. Int. 2021, 149, 110672. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Buranen, S.L.; Zhi-Hai, Y. Construction of single-copy integration vectors for Staphylococcus aureus. Gene 1991, 103, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.; Wang, J.; Zhu, C.; Wang, H.; Fang, M.; Shu, F.; Wang, H.; Hu, Y.; Li, B.; Xue, T. A novel gdmH-related gene, ghl, involved in environmental stress tolerance and vancomycin susceptibility in milk-derived Staphylococcus aureus. Food Res. Int. 2023, 167, 112720. [Google Scholar] [CrossRef]
- Peralta, G.H.; Bergamini, C.V.; Audero, G.; Páez, R.; Wolf, I.V.; Perotti, M.C.; Hynes, E.R. Spray-dried adjunct cultures of autochthonous non-starter lactic acid bacteria. Int. J. Food Microbiol. 2017, 255, 17–24. [Google Scholar] [CrossRef]
- Fan, X.; Sokorai, K.J.; Gurtler, J.B. Advanced oxidation process for the inactivation of Salmonella typhimurium on tomatoes by combination of gaseous ozone and aerosolized hydrogen peroxide. Int. J. Food Microbiol. 2020, 312, 108387. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ming, T.; Zhou, J.; Lu, C.; Wang, R.; Su, X. The Response and Survival Mechanisms of Staphylococcus aureus under High Salinity Stress in Salted Foods. Foods 2022, 11, 1503. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, C.P.; Boyle-Vavra, S.; Roux, A.; Ebine, K.; Sonenshein, A.L.; Daum, R.S. CodY Deletion Enhances In Vivo Virulence of Community-Associated Methicillin-Resistant Staphylococcus aureus Clone USA300. Infect. Immun. 2012, 80, 2382–2389. [Google Scholar] [CrossRef] [PubMed]
- Sonenshein, A.L. CodY, a global regulator of stationary phase and virulence in Gram-positive bacteria. Curr. Opin. Microbiol. 2005, 8, 203–207. [Google Scholar] [CrossRef]
- Lang, E.; Iaconelli, C.; Zoz, F.; Guyot, S.; Alvarez-Martin, P.; Beney, L.; Perrier-Cornet, J.-M.; Gervais, P. Drying parameters greatly affect the destruction of Cronobacter sakazakii and Salmonella Typhimurium in standard buffer and milk. Food Microbiol. 2017, 62, 82–91. [Google Scholar] [CrossRef]
- Wang, X.; Meng, J.; Zhang, J.; Zhou, T.; Zhang, Y.; Yang, B.; Xi, M.; Xia, X. Characterization of Staphylococcus aureus isolated from powdered infant formula milk and infant rice cereal in China. Int. J. Food Microbiol. 2012, 153, 142–147. [Google Scholar] [CrossRef]
- Singh, V.K.; Syring, M.; Singh, A.; Singhal, K.; Dalecki, A.; Johansson, T. An insight into the significance of the DnaK heat shock system in Staphylococcus aureus. Int. J. Med Microbiol. 2012, 302, 242–252. [Google Scholar] [CrossRef]
- Maudsdotter, L.; Imai, S.; Ohniwa, R.L.; Saito, S.; Morikawa, K. Staphylococcus aureus dry stress survivors have a heritable fitness advantage in subsequent dry exposure. Microbes Infect. 2015, 17, 456–461. [Google Scholar] [CrossRef]
- Beavers, W.N.; Skaar, E.P. Neutrophil-generated oxidative stress and protein damage in Staphylococcus aureus. Pathog. Dis. 2016, 74, ftw060. [Google Scholar] [CrossRef]
- Cebrián, G.; Sagarzazu, N.; Aertsen, A.; Pagán, R.; Condón, S.; Mañas, P. Role of the alternative sigma factor sigma on Staphylococcus aureus resistance to stresses of relevance to food preservation. J. Appl. Microbiol. 2009, 107, 187–196. [Google Scholar] [CrossRef]
- Cheung, A.L.; Nishina, K.A.; Trotonda, M.P.; Tamber, S. The SarA protein family of Staphylococcus aureus. Int. J. Biochem. Cell Biol. 2008, 40, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Tong, Y.; Cheng, J.; Abbas, Z.; Li, Z.; Wang, J.; Zhou, Y.; Si, D.; Zhang, R. Biofilm and Small Colony Variants—An Update on Staphylococcus aureus Strategies toward Drug Resistance. Int. J. Mol. Sci. 2022, 23, 1241. [Google Scholar] [CrossRef] [PubMed]
- Painter, K.L.; Strange, E.; Parkhill, J.; Bamford, K.B.; Armstrong-James, D.; Edwards, A.M. Staphylococcus aureus Adapts to Oxidative Stress by Producing H2O2-Resistant Small-Colony Variants via the SOS Response. Infect. Immun. 2015, 83, 1830–1844. [Google Scholar] [CrossRef]
- Belitsky, B.R.; Sonenshein, A.L. Genome-wide identification of Bacillus subtilis CodY-binding sites at single-nucleotide resolution. Proc. Natl. Acad. Sci. USA 2013, 110, 7026–7031. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Chen, X.; Sant’Ana, A.S.; Feng, J.; Ding, T. Pre-Exposure of Foodborne Staphylococcus aureus Isolates to Organic Acids Induces Cross-Adaptation to Mild Heat. Microbiol. Spectr. 2023, 11, e0383222. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Gupta, R.S. Conserved inserts in the Hsp60 (GroEL) and Hsp70 (DnaK) proteins are essential for cellular growth. Mol. Genet. Genom. 2009, 281, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Tomoyasu, T.; Tsuruno, K.; Tanatsugu, R.; Miyazaki, A.; Kondo, H.; Tabata, A.; Whiley, R.A.; Sonomoto, K.; Nagamune, H. Recognizability of heterologous co-chaperones with Streptococcus intermedius DnaK and Escherichia coli DnaK. Microbiol. Immunol. 2018, 62, 681–693. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Relevant Genotype | Reference or Source |
|---|---|---|
| Strains | ||
| WT | RMSA24 with pLI50 | Laboratory stock |
| S. aureus RN4220 | 8325-4, restriction-negative strain | NARSA |
| M-codY | RMSA24 codY-inserted mutant | This work |
| C-codY | RMSA24 codY-inserted mutant with pLI50-cody | This work |
| DH5α | Clone host strain, supE44 ΔlacU169(φ80 lacZΔM15) hsdR17 recA1 endA1 gyrA96 thi-1 relA1 | Invitrogen |
| Plasmids | ||
| pLI50 | Shuttle vector, Apr, Cmr | [49] |
| pLI50-codY | pLI50 with codY | This work |
| Primers Name | Oligonucleotide (5′–3′) | Source |
|---|---|---|
| codY-F | TCTAGAGTCGACCTGCAGGCATGCAAGTGGTCAAGATGTCTCAAGAC | This work |
| codY-R | TTATGCCTAAAAACCTACAGAAGCTTGTCCCAGACTCATCGACTTA | This work |
| check-pLI50-f | CCTGACGTCTAAGAAACCAT | This work |
| check-pLI50-r | CGATAACCACATAACAGTCA | This work |
| hu-F | AAAAAGAAGCTGGTTCAGCAGTAG | This work |
| hu-R | TTTACGTGCAGCACGTTCAC | This work |
| dnaJ-F | AGGATTCAATGGCTCTG | This work |
| dnaJ-R | TGTACCAAATACCGCTTC | This work |
| groEL-F | GTAGGTGCGATTTCAGC | This work |
| groEL-R | AAATGTATGGGCGTTCT | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, H.; Zhu, C.; Shu, F.; Lu, Z.; Wang, H.; Ma, K.; Wang, J.; Lan, R.; Shang, F.; Xue, T. CodY: An Essential Transcriptional Regulator Involved in Environmental Stress Tolerance in Foodborne Staphylococcus aureus RMSA24. Foods 2023, 12, 3166. https://doi.org/10.3390/foods12173166
Pei H, Zhu C, Shu F, Lu Z, Wang H, Ma K, Wang J, Lan R, Shang F, Xue T. CodY: An Essential Transcriptional Regulator Involved in Environmental Stress Tolerance in Foodborne Staphylococcus aureus RMSA24. Foods. 2023; 12(17):3166. https://doi.org/10.3390/foods12173166
Chicago/Turabian StylePei, Hao, Chengfeng Zhu, Fang Shu, Zhengfei Lu, Hui Wang, Kai Ma, Jun Wang, Ranxiang Lan, Fei Shang, and Ting Xue. 2023. "CodY: An Essential Transcriptional Regulator Involved in Environmental Stress Tolerance in Foodborne Staphylococcus aureus RMSA24" Foods 12, no. 17: 3166. https://doi.org/10.3390/foods12173166
APA StylePei, H., Zhu, C., Shu, F., Lu, Z., Wang, H., Ma, K., Wang, J., Lan, R., Shang, F., & Xue, T. (2023). CodY: An Essential Transcriptional Regulator Involved in Environmental Stress Tolerance in Foodborne Staphylococcus aureus RMSA24. Foods, 12(17), 3166. https://doi.org/10.3390/foods12173166