
Use of X-Irradiations in Reducing the Waste of Aflatoxin-Contaminated Pistachios and Evaluation of the Physicochemical Properties of the Irradiated Product



Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Fungal Inoculation and AFB1 Contamination of Pistachio Nuts
2.3. Electron Beam Irradiation
2.4. Fungal Load Determination
2.5. Aflatoxin B1 Extraction and Detection by HPLC
2.6. LC-MS Analysis of Degraded AFB1
2.7. Determination of Total Phenolic Content (TPC) and Antioxidant Activity (AOA)
2.8. Determination of Chlorophyll and Carotenoid Pigments
2.9. Color Analysis
2.10. Total Soluble Protein (TSP)
2.11. Native-PAGE and SDS-PAGE Experiments
2.12. Determination of Malondialdehyde (MDA)
2.13. Analysis of Fatty Acid Composition
2.14. Sensory Evaluation
2.15. Statistics and Data Analysis
3. Results and Discussion
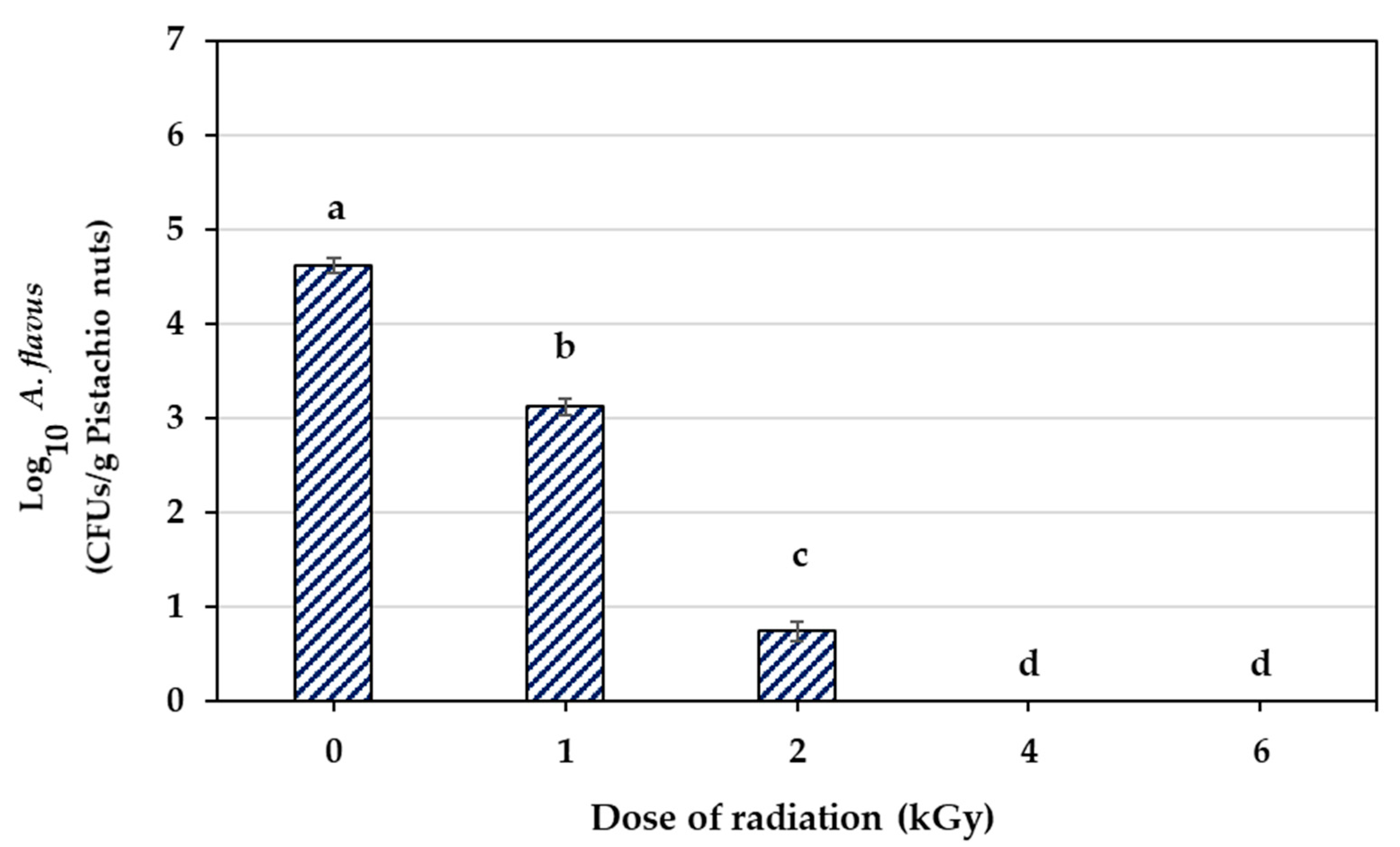
3.1. Inactivation of A. flavus by Electron Beam Irradiation
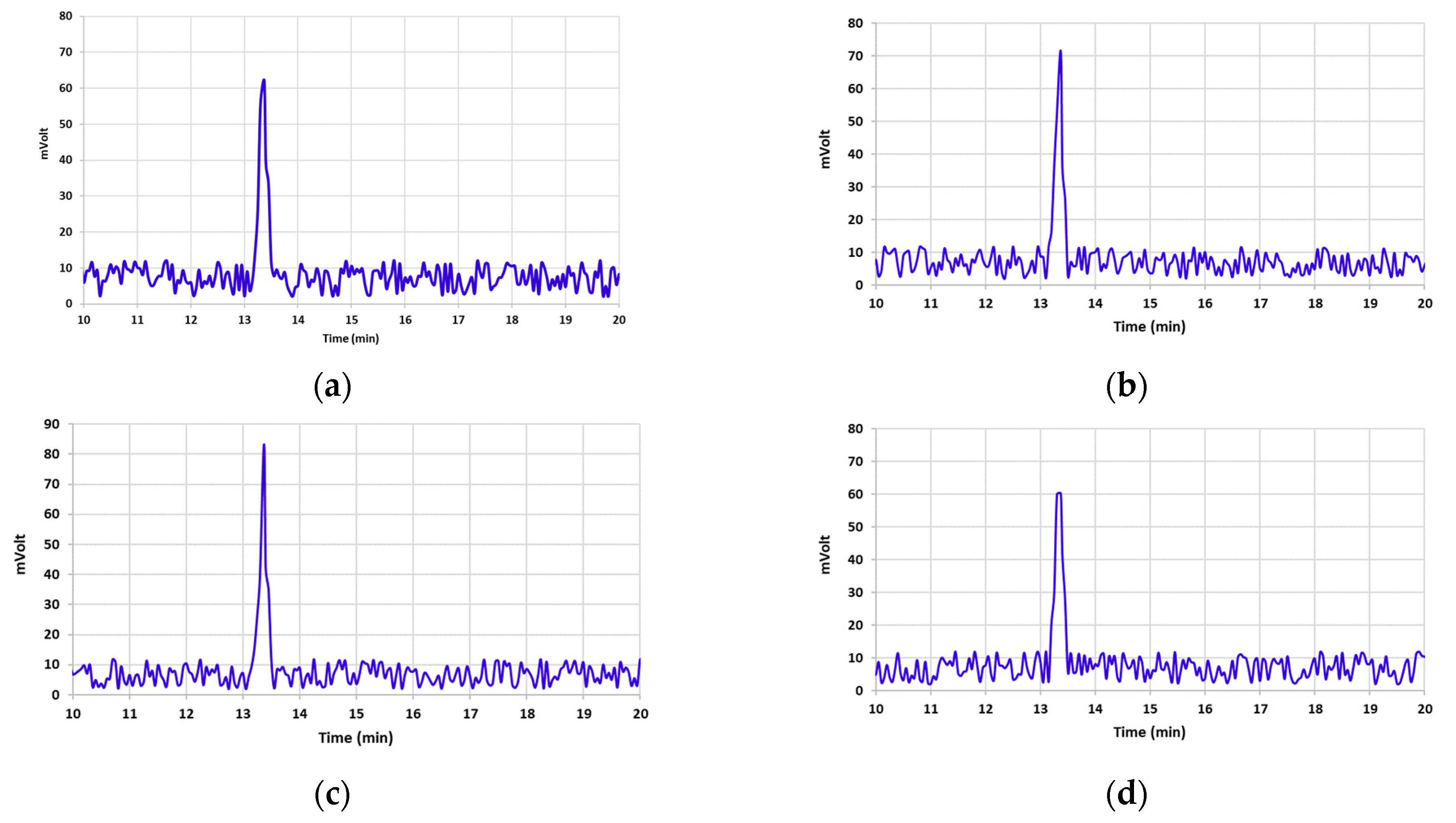
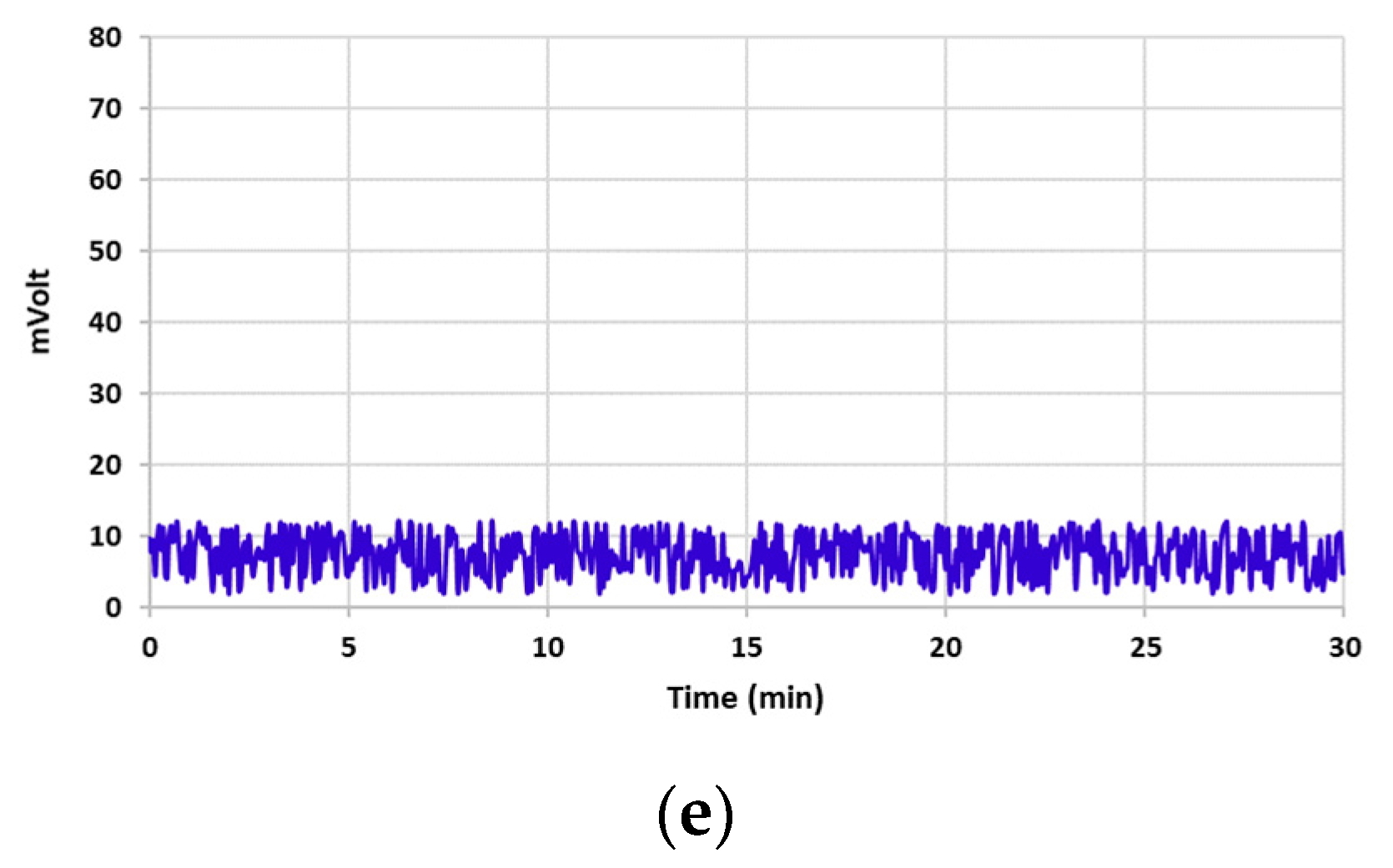
3.2. Aflatoxin B1 Degradation
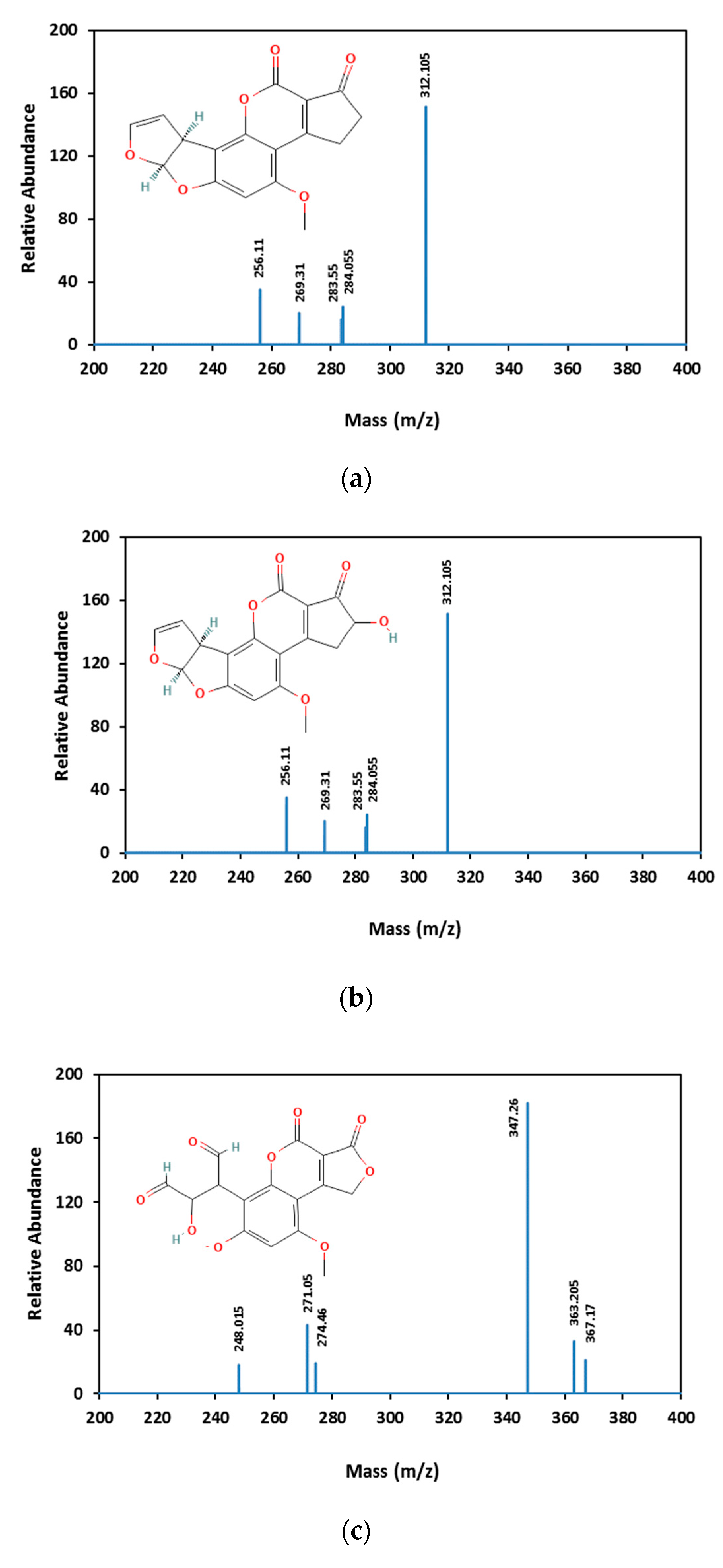
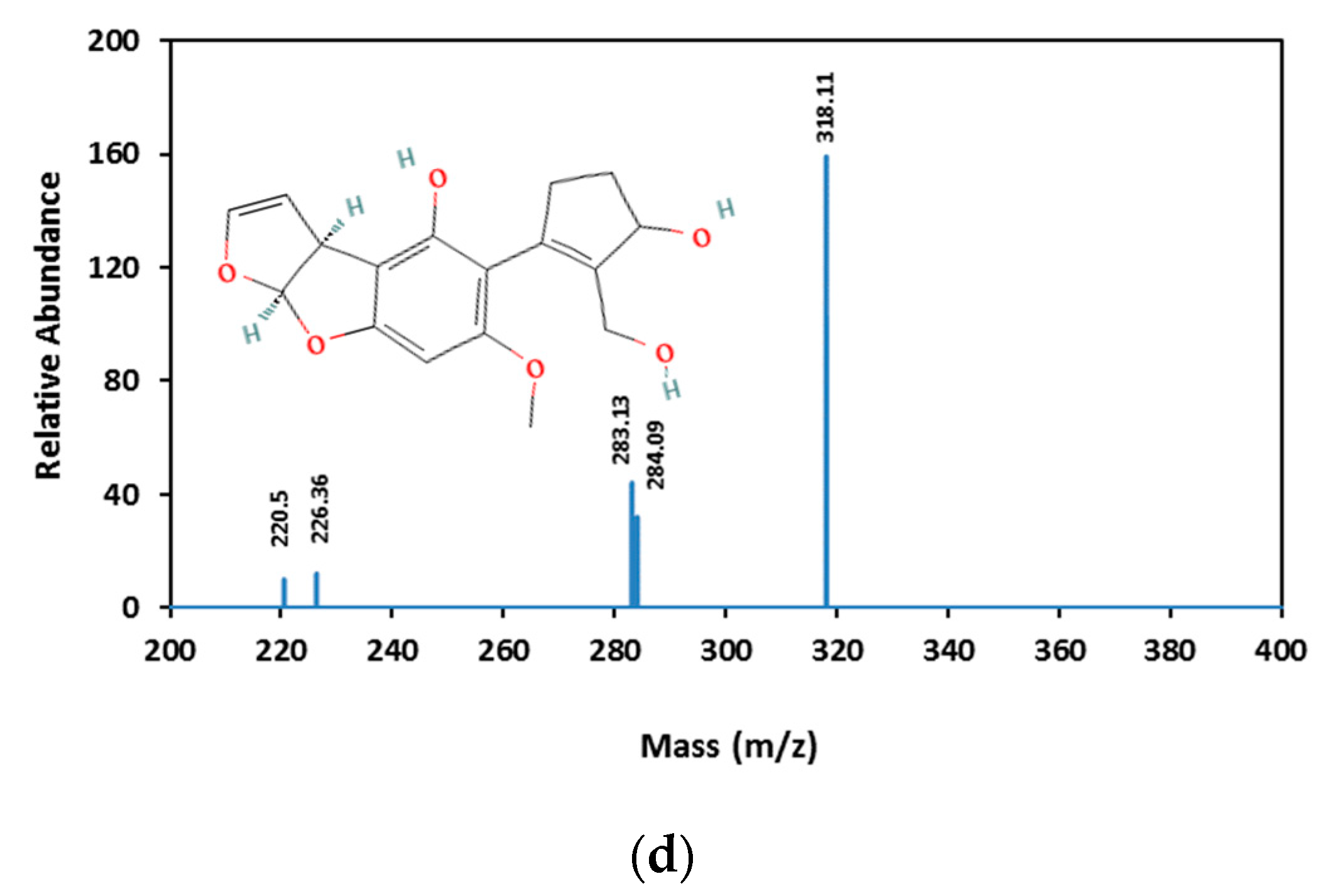
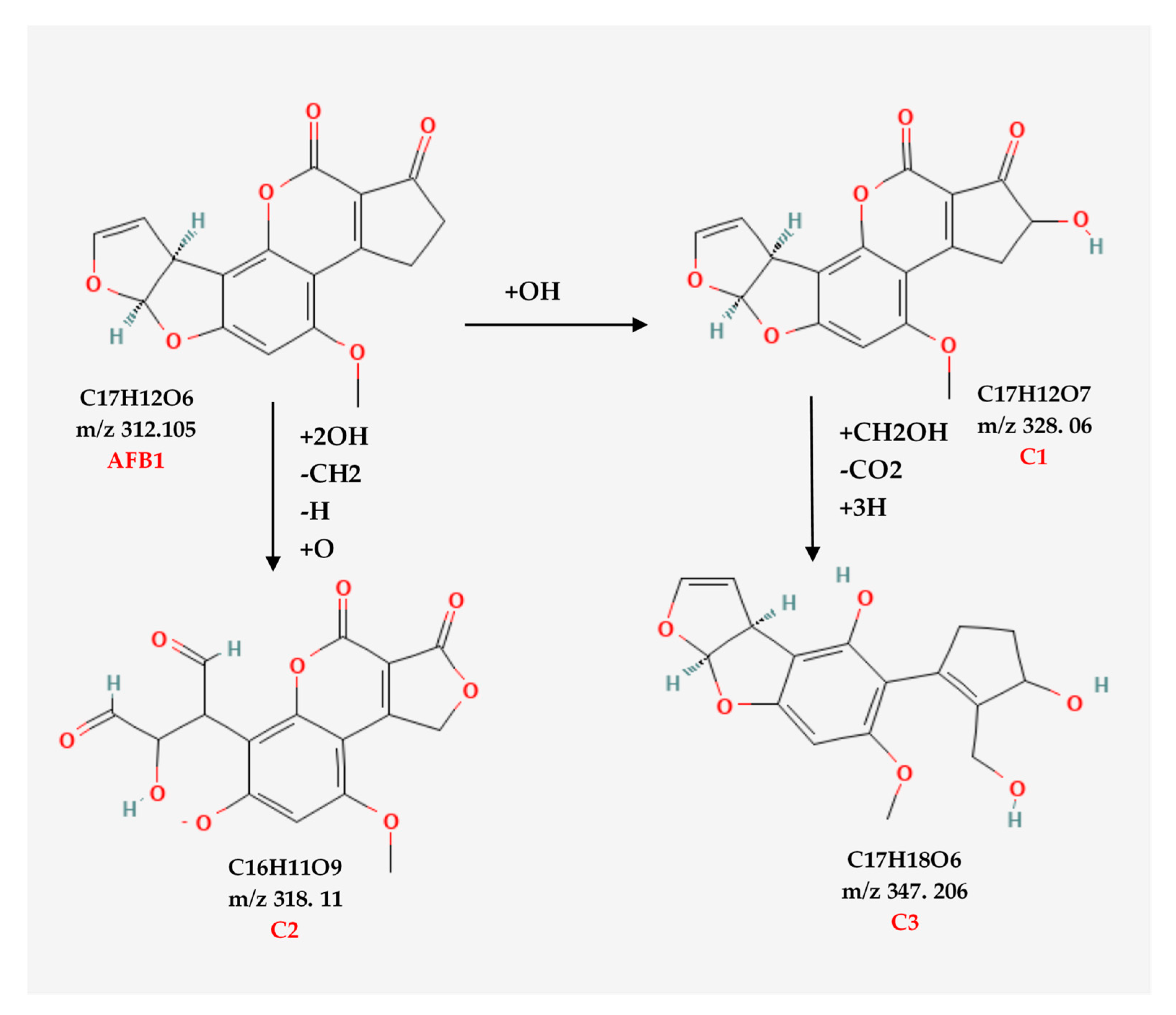
3.3. Aflatoxin B1 Degradation Products
3.4. Total Phenolic Content (TPC) and Antioxidant Activity (AOA)
3.5. Chlorophyll and Carotenoids Content
3.6. Color
3.7. Solubility of Proteins (TSP)
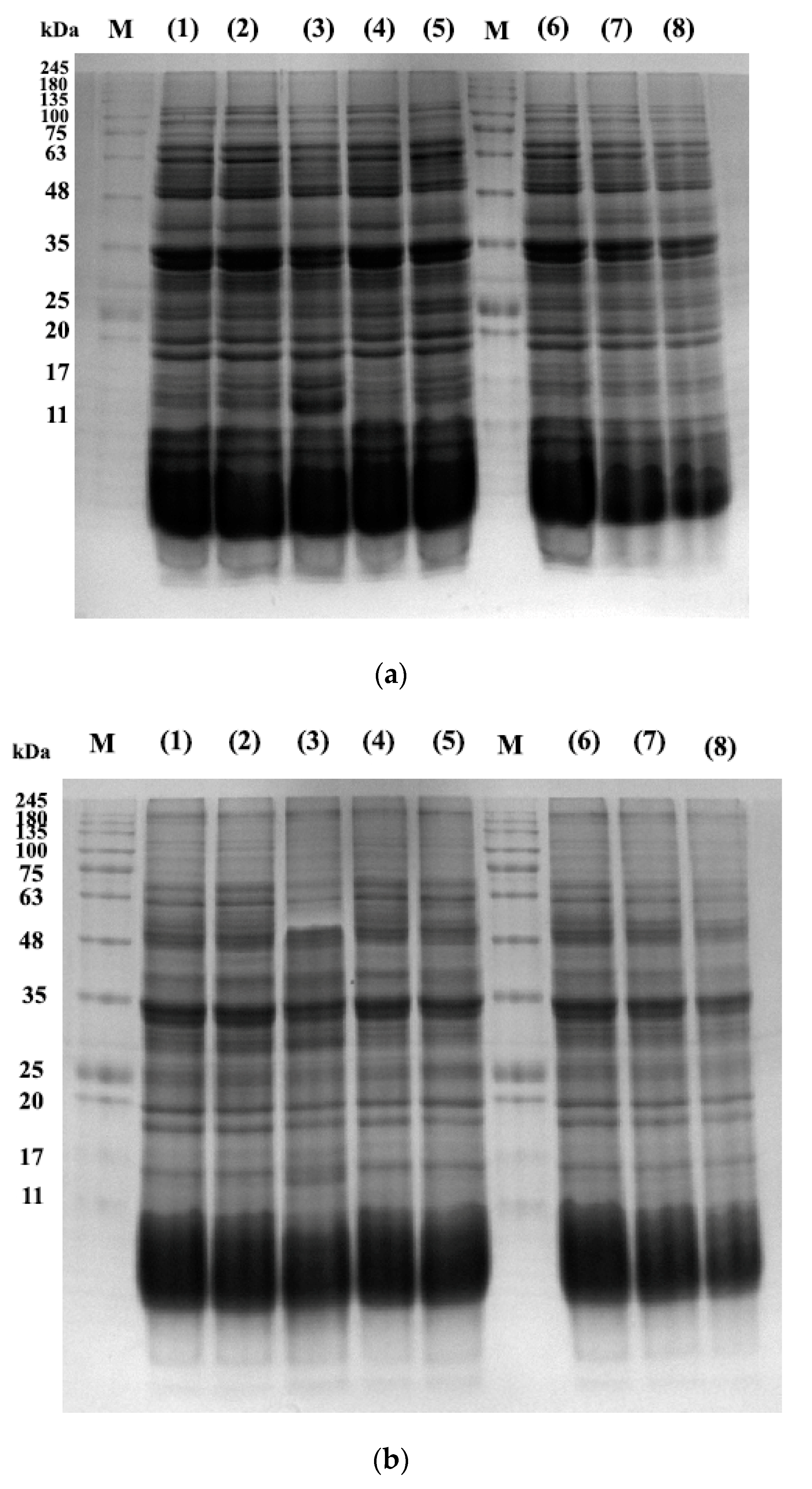
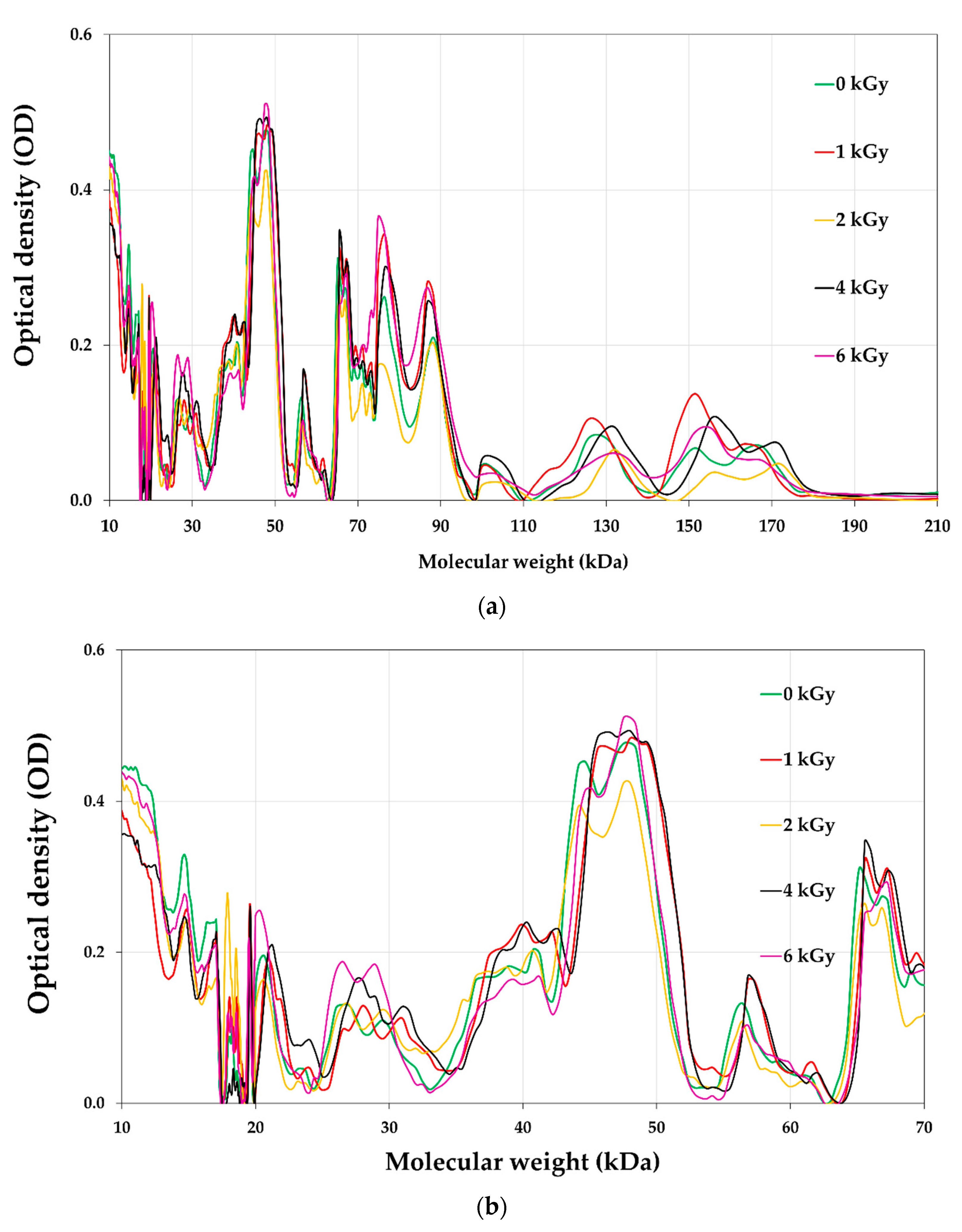
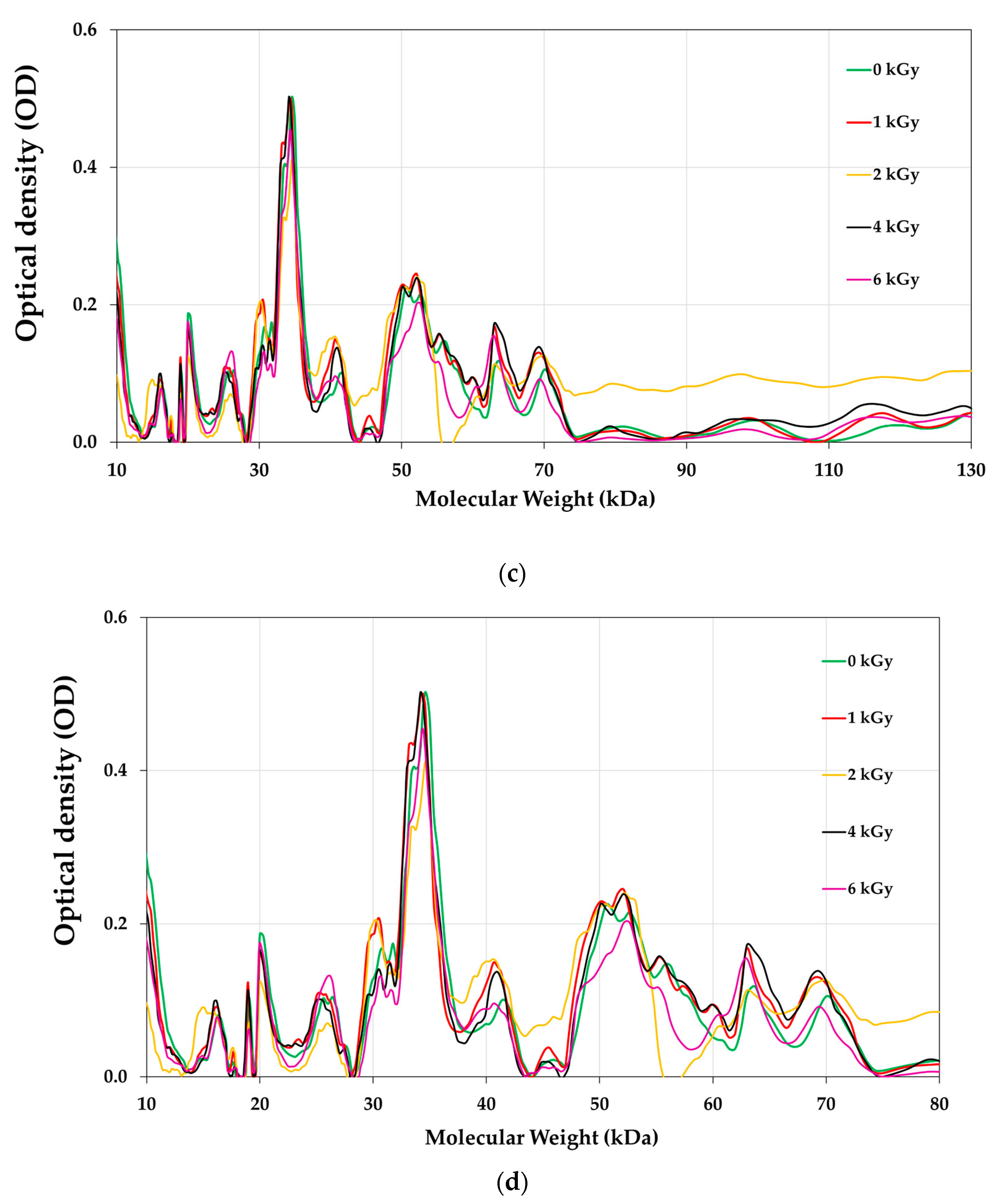
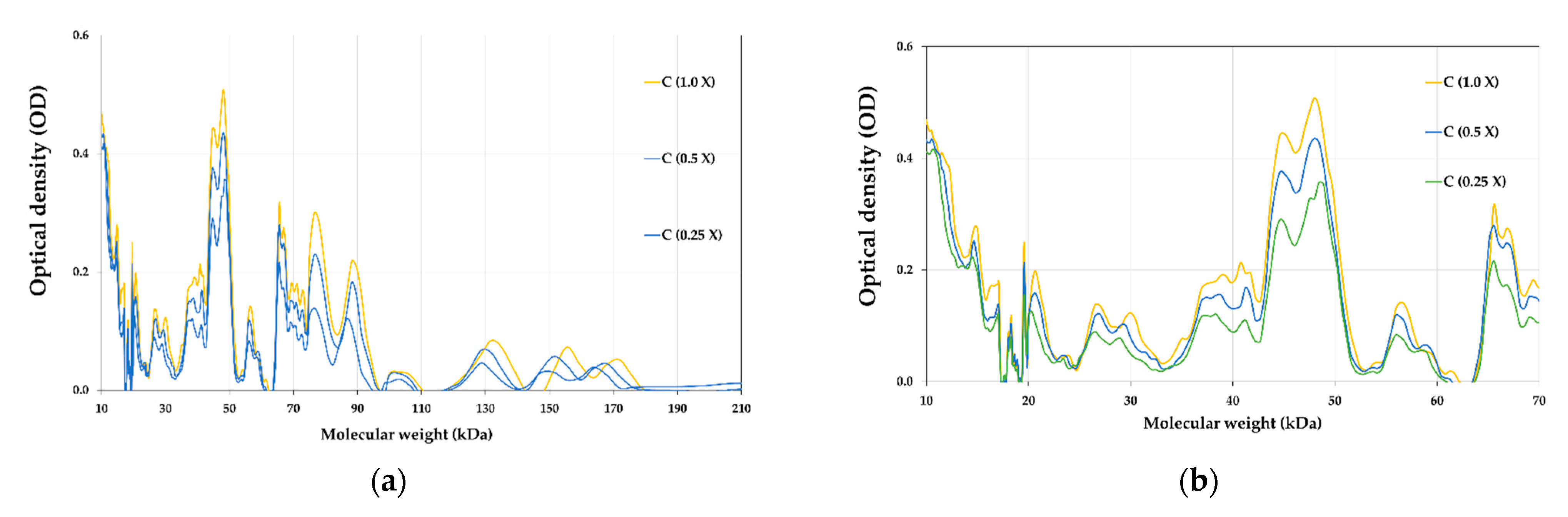
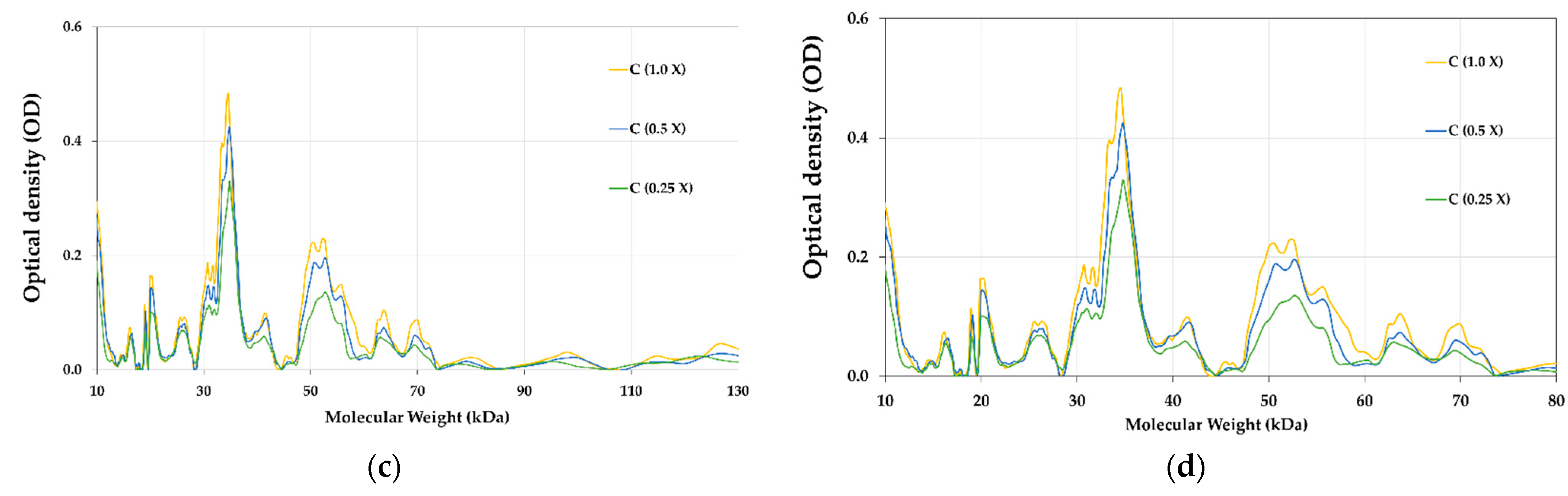
3.8. Proteins Profile of E-Beam Irradiated Pistachio Nuts
3.9. Malondialdehyde Content (MDA)
3.10. Profile of Fatty Acids
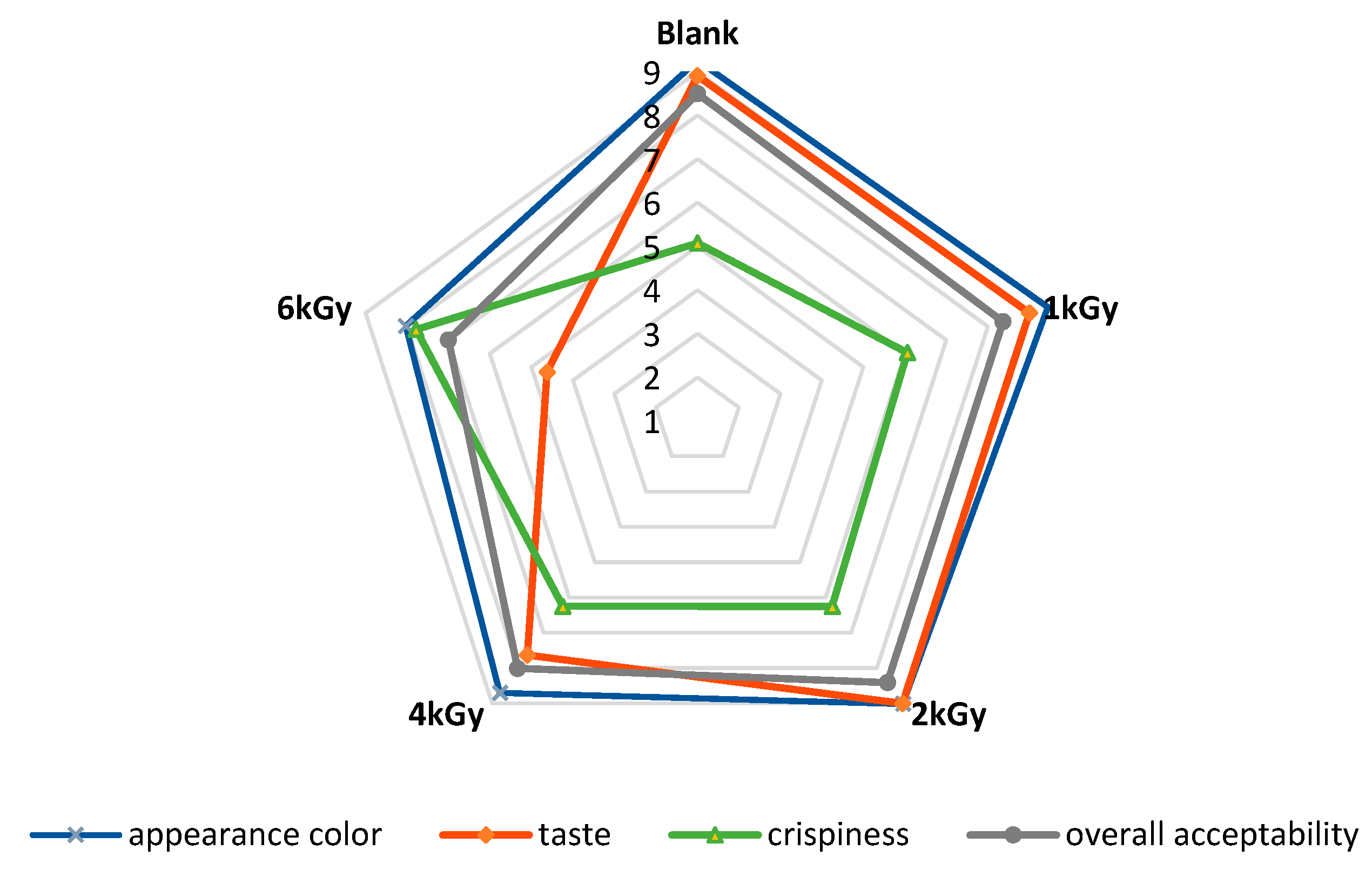
3.11. Sensory Evaluation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Markov, K.; Mihaljević, B.; Domijan, A.-M.; Pleadin, J.; Delaš, F.; Frece, J. Inactivation of aflatoxigenic fungi and the reduction of aflatoxin B1 in vitro and in situ using gamma irradiation. Food Control 2015, 54, 79–85. [Google Scholar] [CrossRef]
- Assuncao, E.; Reis, T.A.; Baquiao, A.C.; Correa, B. Effects of gamma and electron beam radiation on Brazil nuts artificially inoculated with Aspergillus flavus. J. Food Prot. 2015, 78, 1397–1401. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, I.; Orfi, M.; Shamma, M. Effect of gamma radiation on the inactivation of aflatoxin B1 in food and feed crops. Braz. J. Microbiol. 2008, 39, 787–791. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Sen, Y.; Onal-Ulusoy, B.; Mutlu, M. Detoxification of hazelnuts by different cold plasmas and gamma irradiation treatments. Innov. Food Sci. Emerg. Technol. 2019, 54, 252–259. [Google Scholar] [CrossRef]
- Makari, M.; Hojjati, M.; Shahbazi, S.; Askari, H. Effect of Co-60 gamma irradiation on Aspergillus flavus, Aflatoxin B1 and qualitative characteristics of pistachio nuts (Pistacia vera L.). J. Food Meas. Charact. 2021, 15, 5256–5265. [Google Scholar] [CrossRef]
- Khalil, O.A.; Hammad, A.A.; Sebaei, A.S. Aspergillus flavus and Aspergillus ochraceus inhibition and reduction of aflatoxins and ochratoxin A in maize by irradiation. Toxicon 2021, 198, 111–120. [Google Scholar] [CrossRef]
- Jalili, M.; Jinap, S.; Noranizan, M. Aflatoxins and ochratoxin a reduction in black and white pepper by gamma radiation. Radiat. Phys. Chem. 2012, 81, 1786–1788. [Google Scholar] [CrossRef]
- Iqbal, S.Z.; Bhatti, I.A.; Asi, M.R.; Zuber, M.; Shahid, M.; Parveen, I. Effect of γ irradiation on fungal load and aflatoxins reduction in red chillies. Radiat. Phys. Chem. 2013, 82, 80–84. [Google Scholar] [CrossRef]
- Boonchoo, T.; Jitareerat, P.; Photchanachai, S.; Chinaphuti, A. Effect of gamma irradiation on Aspergillus flavus and brown rice quality during storage. In Proceedings of the 2nd International Symposium on the New Frontier of Irradiated Food and Non Food Products, KMUTT, Bangkok, Thailand, 22–23 September 2005. [Google Scholar]
- Aquino, S.; Ferreira, F.; Ribeiro, D.H.B.; Corrêa, B.; Greiner, R.; Villavicencio, A.L.C.H. Evaluation of viability of Aspergillus flavus and aflatoxins degradation in irradiated samples of maize. Braz. J. Microbiol. 2005, 36, 352–356. [Google Scholar] [CrossRef]
- Freita-Silva, O.; de Oliveira, P.S.; Freire Júnior, M. Potential of electron beams to control mycotoxigenic fungi in food. Food Eng. Rev. 2015, 7, 160–170. [Google Scholar] [CrossRef]
- WHO. Wholesomeness of Irradiated Food; 659; World Health Organization: Geneva, Switzerland, 1981; p. 34.
- Kume, T.; Todoriki, S. Food irradiation in Asia, the European Union, and the United States: A status update. Radioisotopes 2013, 62, 291–299. [Google Scholar] [CrossRef]
- EC. Directive 1999/2/EC of the European parliament and of the council of 22 February 1999 on the approximation of the laws of the Member States concerning foods and food ingredients treated with ionising radiation. Off. J. Eur. Union 1999, L66, 16–22. [Google Scholar]
- EC. Directive 1999/3/EC of the European parliament and of the council of 22 February 1999 on the establishment of a Community list of foods and food ingredients treated with ionising radiation. Off. J. Eur. Union 1999, L66, 24–25. [Google Scholar]
- Sánchez-Bravo, P.; Noguera-Artiaga, L.; Gómez-López, V.M.; Carbonell-Barrachina, Á.A.; Gabaldón, J.A.; Pérez-López, A.J. Impact of Non-Thermal Technologies on the Quality of Nuts: A Review. Foods 2022, 11, 3891. [Google Scholar] [CrossRef]
- Farkas, J. Irradiation for better foods. Trends Food Sci. Technol. 2006, 17, 148–152. [Google Scholar] [CrossRef]
- Calado, T.; Venâncio, A.; Abrunhosa, L. Irradiation for mold and mycotoxin control: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1049–1061. [Google Scholar] [CrossRef]
- Mexis, S.F.; Kontominas, M.G. Effect of gamma irradiation on the physico-chemical and sensory properties of raw shelled peanuts (Arachis hypogaea L.) and pistachio nuts (Pistacia vera L.). J. Sci. Food Agric. 2009, 89, 867–875. [Google Scholar] [CrossRef]
- EC. Commission Regulation (EU) 2023/915 of 25 April 2023 on maximum levels for certain contaminants in food and repealing Regulation (EC) No 1881/2006. Off. J. Eur. Union 2023, L119, 103–157. [Google Scholar]
- Alinezhad, M.; Hojjati, M.; Barzegar, H.; Shahbazi, S.; Askari, H. Effect of gamma irradiation on the physicochemical properties of pistachio (Pistacia vera L.) nuts. J. Food Meas. Charact. 2021, 15, 199–209. [Google Scholar] [CrossRef]
- Krishnan, V.; Singh, A.; Thimmegowda, V.; Singh, B.; Dahuja, A.; Rai, R.D.; Sachdev, A. Low gamma irradiation effects on protein profile, solubility, oxidation, scavenger ability and bioavailability of essential minerals in black and yellow Indian soybean (Glycine max L.) varieties. J. Radioanal. Nucl. Chem. 2016, 307, 49–57. [Google Scholar] [CrossRef]
- Gecgel, U.; Gumus, T.; Tasan, M.; Daglioglu, O.; Arici, M. Determination of fatty acid composition of γ-irradiated hazelnuts, walnuts, almonds, and pistachios. Radiat. Phys. Chem. 2011, 80, 578–581. [Google Scholar] [CrossRef]
- Iram, W.; Anjum, T.; Iqbal, M.; Ghaffar, A.; Abbas, M.; Khan, A.M. Structural analysis and biological toxicity of aflatoxins B1 and B2 degradation products following detoxification by Ocimum basilicum and Cassia fistula aqueous extracts. Front. Microbiol. 2016, 7, 1105. [Google Scholar] [CrossRef] [PubMed]
- Hojjati, M.; Noguera-Artiaga, L.; Wojdyło, A.; Carbonell-Barrachina, Á.A. Effects of microwave roasting on physicochemical properties of pistachios (Pistaciavera L.). Food Sci. Biotechnol. 2015, 24, 1995–2001. [Google Scholar] [CrossRef]
- Byun, K.-H.; Cho, M.-J.; Park, S.-Y.; Chun, H.S.; Ha, S.-D. Effects of gamma ray, electron beam, and X-ray on the reduction of Aspergillus flavus on red pepper powder (Capsicum annuum L.) and gochujang (red pepper paste). Food Sci. Technol. Int. 2019, 25, 649–658. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Mexis, S.; Kontominas, M. Effect of γ-irradiation on the physicochemical and sensory properties of cashew nuts (Anacardium occidentale L.). LWT-Food Sci. Technol. 2009, 42, 1501–1507. [Google Scholar] [CrossRef]
- Jalili, M.; Scotter, M. A review of aflatoxin M1 in liquid milk. Iran. J. Health Saf. Environ. 2015, 2, 283–295. [Google Scholar]
- Liu, R.; Wang, R.; Lu, J.; Chang, M.; Jin, Q.; Du, Z.; Wang, S.; Li, Q.; Wang, X. Degradation of AFB1 in aqueous medium by electron beam irradiation: Kinetics, pathway and toxicology. Food Control 2016, 66, 151–157. [Google Scholar] [CrossRef]
- Liu, R.; Lu, M.; Wang, R.; Wang, S.; Chang, M.; Jin, Q.; Wang, X. Degradation of aflatoxin B1 in peanut meal by electron beam irradiation. Int. J. Food Prop. 2018, 21, 892–901. [Google Scholar] [CrossRef]
- Akbari, M.; Farajpour, M.; Aalifar, M.; Sadat Hosseini, M. Gamma irradiation affects the total phenol, anthocyanin and antioxidant properties in three different persian pistachio nuts. Nat. Prod. Res. 2018, 32, 322–326. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kim, M.-S.; Kim, H.-G.; Yook, H.-S. Inactivation of contaminated fungi and antioxidant effects of peach (Prunus persica L. Batsch cv Dangeumdo) by 0.5–2 kGy gamma irradiation. Radiat. Phys. Chem. 2010, 79, 495–501. [Google Scholar] [CrossRef]
- Dixit, A.K.; Bhatnagar, D.; Kumar, V.; Rani, A.; Manjaya, J.; Bhatnagar, D. Gamma irradiation induced enhancement in isoflavones, total phenol, anthocyanin and antioxidant properties of varying seed coat colored soybean. J. Agric. Food Chem. 2010, 58, 4298–4302. [Google Scholar] [CrossRef]
- Kiong, A.L.P.; Lai, A.G.; Hussein, S.; Harun, A.R. Physiological responses of Orthosiphon stamineus plantlets to gamma irradiation. Am.-Eurasian J. Sustain. Agric. 2008, 2, 135–149. [Google Scholar]
- Topuz, A.; Ozdemir, F. Influences of γ-irradiation and storage on the carotenoids of sun-dried and dehydrated paprika. J. Agric. Food Chem. 2003, 51, 4972–4977. [Google Scholar] [CrossRef]
- Prakash, A.; de Jesús Ornelas-Paz, J. Irradiation of fruits and vegetables. In Postharvest Technology of Perishable Horticultural Commodities; Elsevier, Woodhead Publishing: Duxford, UK, 2019; pp. 563–589. [Google Scholar]
- Ramamurthy, M.; Kamat, A.; Kakatkar, A.; Ghadge, N.; Bhushan, B.; Alur, M. Improvement of shelf-life and microbiological quality of minimally processed refrigerated capsicum by gamma irradiation. Int. J. Food Sci. Nutr. 2004, 55, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Gölge, E.; Ova, G. The effects of food irradiation on quality of pine nut kernels. Radiat. Phys. Chem. 2008, 77, 365–369. [Google Scholar] [CrossRef]
- Sánchez-Bel, P.; Egea, I.; Romojaro, F.; Martínez-Madrid, M.C. Sensorial and chemical quality of electron beam irradiated almonds (Prunus amygdalus). LWT-Food Sci. Technol. 2008, 41, 442–449. [Google Scholar] [CrossRef]
- Güler, S.K.; Bostan, S.Z.; Çon, A.H. Effects of gamma irradiation on chemical and sensory characteristics of natural hazelnut kernels. Postharvest Biol. Technol. 2017, 123, 12–21. [Google Scholar] [CrossRef]
- Li, T.; Wang, L.; Chen, Z.; Sun, D.; Li, Y. Electron beam irradiation induced aggregation behaviour, structural and functional properties changes of rice proteins and hydrolysates. Food Hydrocoll. 2019, 97, 105192. [Google Scholar] [CrossRef]
- Malik, M.A.; Saini, C.S. Gamma irradiation of alkali extracted protein isolate from dephenolized sunflower meal. LWT 2017, 84, 204–211. [Google Scholar] [CrossRef]
- Afify, A.E.-M.M.; Rashed, M.M.; Mahmoud, E.A.; El-Beltagi, H.S. Effect of gamma radiation on protein profile, protein fraction and solubility’s of three oil seeds: Soybean, peanut and sesame. Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 90–98. [Google Scholar] [CrossRef]
- Naei, V.Y.; Sankian, M.; Moghadam, M.; Farshidi, N.; Ayati, S.H.; Hamid, F.; Varasteh, A.-R. The influence of gamma radiation processing on the allergenicity of main pistachio allergens. Rep. Biochem. Mol. Biol. 2019, 7, 150. [Google Scholar]
- Sirisoontaralak, P.; Noomhorm, A. Changes to physicochemical properties and aroma of irradiated rice. J. Stored Prod. Res. 2006, 42, 264–276. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose of Irradiation (k Gy) | |||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 4 | 6 | |
| Aflatoxin concentration (ppb) | 692.1 ± 0.12 a | 546.2 ± 0.14 b | 168.5 ± 0.21 c | 2.10 ± 0.08 d | 0.0 ± 0.0 d |
| Properties | Dose of E-Beam Irradiation (kGy) | ||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 4 | 6 | |
| TPC (mg GAE/g) | 6.61 ± 0.10 bc | 6.48 ± 0.17 c | 7.65 ± 0.36 a | 6.85 ± 0.38 bc | 7.02 ± 0.15 b |
| Inhibitory of DPPH activity (1 g/30 mL, %) | 82.07 ± 1.45 d | 81.06 ± 0.95 b | 78.17 ± 1.28 a | 78.74 ± 1.05 c | 77.21 ± 0.73 e |
| Inhibitory of ABTS activity (1 g/30 mL, %) | 78.85 ± 0.42 a | 78.92 ± 0.21 a | 77.28 ± 0.26 b | 77.72 b ± 0.13 b | 76.03 ± 0.13 c |
| TSP (mg/g) | 1.59 ± 0.11 c | 1.81 ± 0.07 b | 2.10 ± 0.04 a | 1.67 ± 0.03 c | 1.71 ± 0.04 bc |
| MDA (n mol. g−1) | 7.01 ± 0.15 e | 7.61 ± 0.20 d | 8.78 ± 0.29 c | 11.00 ± 0.23 b | 12.18 ± 0.34 a |
| Pigments | |||||
| Chlorophyll a | 17.15 ± 0.93 a | 12.99 ± 1.00 b | 13.47 ± 0.87 b | 10.55 ± 0.53 c | 9.07 ± 0.28 d |
| Chlorophyll b | 22.40 ± 2.30 a | 22.73 ± 0.71 a | 22.41 ± 0.72 a | 12.88 ± 1.10 b | 14.24 ± 0.77 b |
| Total chlorophyll | 39.54 ± 3.21 a | 35.72 ± 1.28 b | 35.88 ± 0.32 b | 23.43 ± 1.65 c | 23.31 ± 0.91 c |
| Total carotenoid | 19.33 ± 0.65 a | 13.04 ± 0.54 d | 15.12 ± 0.90 b | 14.39 ± 0.60 bc | 13.96 ± 0.36 cd |
| Instrumental color | |||||
| Lightness (L) | 59.03 ± 1.19 a | 58.76 ± 0.72 a | 58.75 ± 0.61 a | 57.39 ± 0.59 b | 55.18 ± 1.09 c |
| Redness (a*) | −5.11 ± 0.16 b | −5.07 ± 0.15 b | −5.07 ± 0.11 b | −4.93 ± 0.05 b | −4.12 ± 0.09 a |
| Yellowness (b*) | 29.36 ± 0.89 a | 29.55 ± 0.48 a | 29.53 ± 0.41 a | 27.35 ± 0.63 b | 27.43 ± 0.96 b |
| ΔE | 00.00 ± 00.00 d | 0.33 ± 0.62 c | 0.33 ± 0.75 c | 2.60 ± 0.66 b | 4.42 ± 0.14 a |
| X-ray Irradiation Dose (kGy) | |||||
|---|---|---|---|---|---|
| Compounds | 0 (Blank) | 1 | 2 | 4 | 6 |
| Myristic acid (14:0) | 0.16 ± 0.03 a | 0.14 ± 0.02 a | 0.14 ± 0.02 a | 0.13 ± 0.01 a | 0.15 ± 0.01 a |
| Myristoleic acid (14:1) | 0.01 ± 0.01 a | 0.03 ± 0.00 a | nd b | nd b | nd b |
| Pentadecanoic acid (15:0) | 0.02 ± 0.01 a | 0.04 ± 0.01 a | 0.03 ± 0.02 a | 0.04 ± 0.02 a | 0.05 ± 0.02 a |
| Palmitic acid (16:0) | 11.21 ± 0.15 c | 11.32 ± 0.30 c | 11.38 ± 0.06 c | 12.17 ± 0.32 b | 14.58 ± 0.48 a |
| Palmitoleic acid (16:1) | 1.16 ± 0.09 ab | 1.15 ± 0.07 ab | 1.16 ± 0.12 ab | 1.09 ± 0.14 b | 1.27 ± 0.05 a |
| Margaric acid (17:0) | 0.02 ± 0.01 b | 0.03 ± 0.00 ab | 0.02 ± 0.01 b | 0.05 ± 0.00 a | 0.02 ± 0.01 ab |
| Stearic acid (18:0) | 1.78 ± 0.31 b | 1.59 ± 0.08 b | 1.86 ± 0.17 b | 2.97 ± 0.31 a | 3.68 ± 0.84 a |
| Oleic acid (18:1) | 61.34 ± 0.17 a | 61.73 ± 0.34 a | 61.77 ± 0.19 a | 60.53 ± 0.58 a | 58.16 ± 0.99 b |
| Linoleic acid (18:2) | 22.10 ± 0.37 a | 21.92 ± 0.12 a | 21.86 ± 0.25 a | 21.47 ± 0.16 a | 18.75 ± 0.62 b |
| Linolenic acid (18:3) | 0.56 ± 0.03 a | 0.58 ± 0.02 a | 0.57 ± 0.03 a | 0.55 ± 0.05 a | 0.44 ± 0.07 b |
| Arachidic acid (20:0) | 0.28 ± 0.01 a | 0.26 ± 0.01 a | 0.23 ± 0.02 a | 0.26 ± 0.07 a | 0.25 ± 0.04 a |
| Gondoic acid (20:1) | 0.83 ± 0.01 a | 0.79 ± 0.07 a | 0.83 ± 0.08 a | 0.82 ± 0.07 a | 0.82 ± 0.05 a |
| Behenic acid (22:0) | 0.07 ± 0.03 a | 0.08 ± 0.02 a | 0.07 ± 0.02 a | 0.06 ± 0.01 a | 0.08 ± 0.02 a |
| ƩMUFA | 63.34 ± 0.06 a | 63.68 ± 0.34 a | 63.76 ± 0.23 a | 62.45 ± 0.53 ab | 60.26 ± 1.05 c |
| ƩPUFA | 22.66 ± 0.31 a | 22.51 ± 0.11 a | 22.44 ± 0.25 a | 22.00 ± 0.11 a | 19.19 ± 0.62 b |
| ƩUSFA | 86.00 ± 0.38 a | 86.19 ± 0.37 a | 86.21 ± 0.10 a | 84.53 ± 0.63 b | 79.45 ± 1.64 c |
| ƩSFA | 13.45 ± 0.42 c | 13.41 ± 0.27 c | 13.74 ± 0.24 c | 15.68 ± 0.38 b | 18.83 ± 1.08 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hojjati, M.; Shahbazi, S.; Askari, H.; Makari, M. Use of X-Irradiations in Reducing the Waste of Aflatoxin-Contaminated Pistachios and Evaluation of the Physicochemical Properties of the Irradiated Product. Foods 2023, 12, 3040. https://doi.org/10.3390/foods12163040
Hojjati M, Shahbazi S, Askari H, Makari M. Use of X-Irradiations in Reducing the Waste of Aflatoxin-Contaminated Pistachios and Evaluation of the Physicochemical Properties of the Irradiated Product. Foods. 2023; 12(16):3040. https://doi.org/10.3390/foods12163040
Chicago/Turabian StyleHojjati, Mohammad, Samira Shahbazi, Hamed Askari, and Mina Makari. 2023. "Use of X-Irradiations in Reducing the Waste of Aflatoxin-Contaminated Pistachios and Evaluation of the Physicochemical Properties of the Irradiated Product" Foods 12, no. 16: 3040. https://doi.org/10.3390/foods12163040
APA StyleHojjati, M., Shahbazi, S., Askari, H., & Makari, M. (2023). Use of X-Irradiations in Reducing the Waste of Aflatoxin-Contaminated Pistachios and Evaluation of the Physicochemical Properties of the Irradiated Product. Foods, 12(16), 3040. https://doi.org/10.3390/foods12163040