
The Role of Green Tea on the Regulation of Gut Microbes and Prevention of High-Fat Diet-Induced Metabolic Syndrome in Mice


 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Green Tea Infusion
2.2. Animal and Treatments
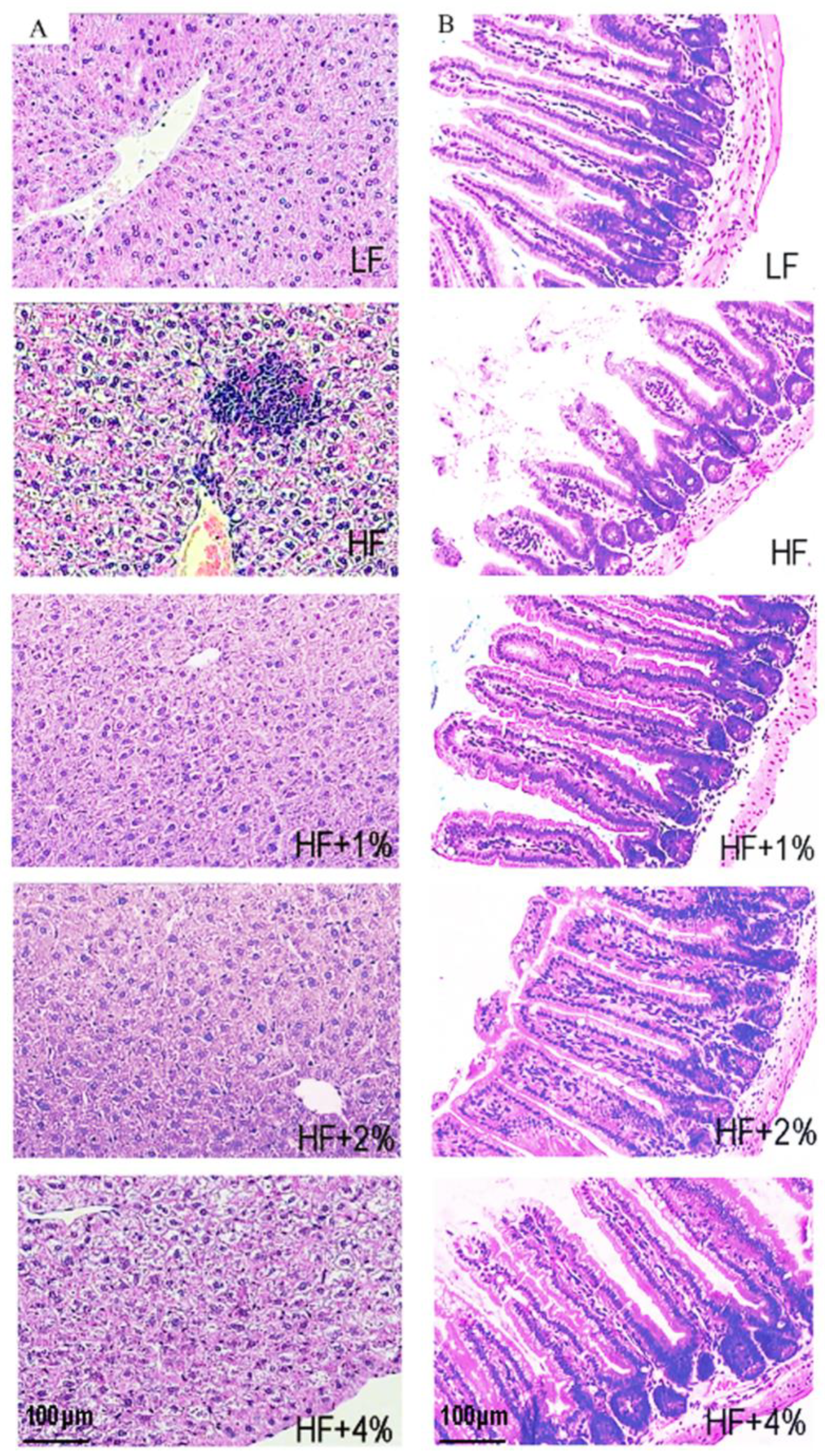
2.3. Tissue Preparation and Biomarker Assessments
2.4. Gut Microbiota Classification by Next Generation Sequencing (NGS)
2.5. Statistical Analysis
3. Results
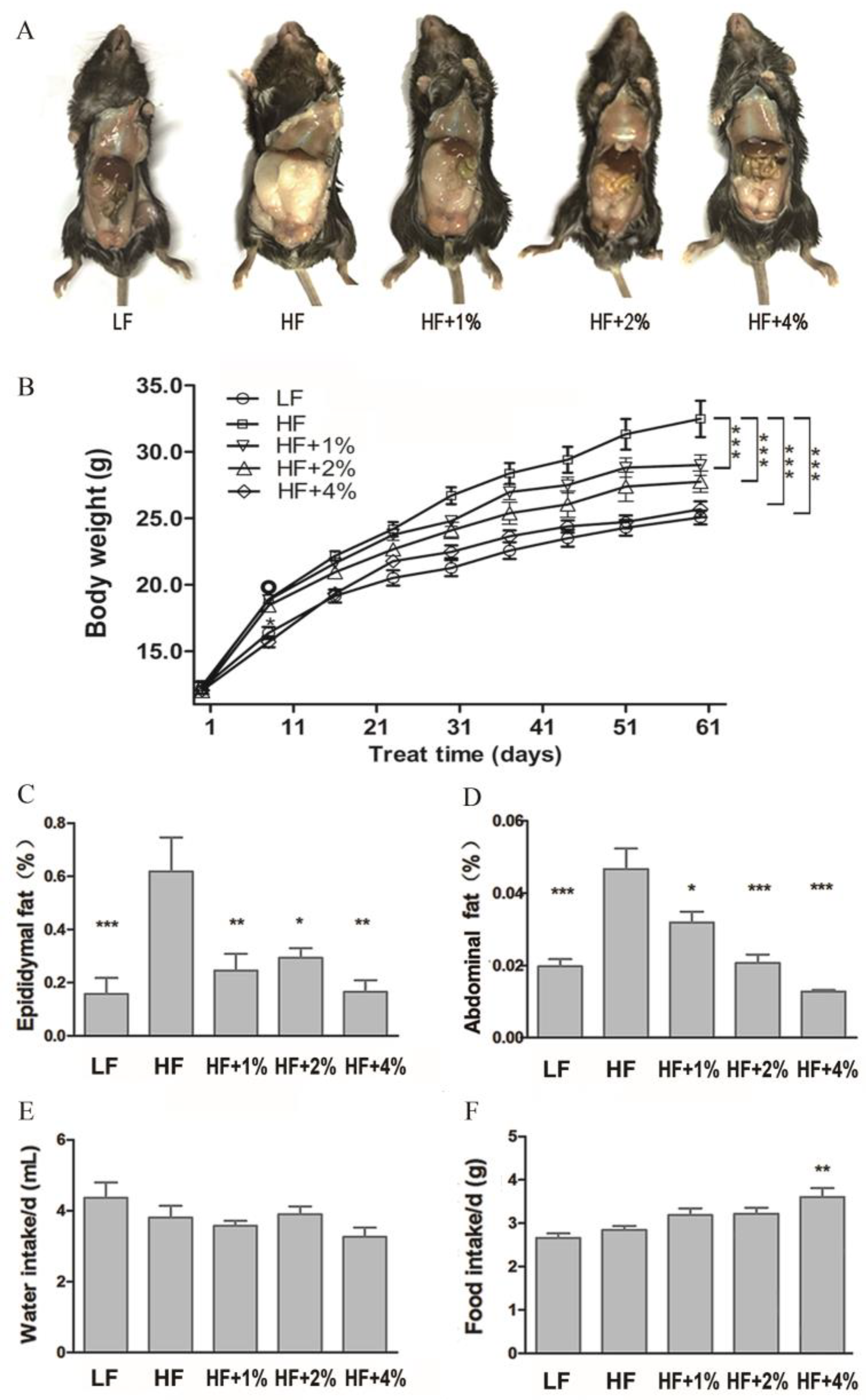
3.1. Effect of Green Tea on Body Weight- and Fat-Associated Biochemical Indices of HF-Induced Obese Mice
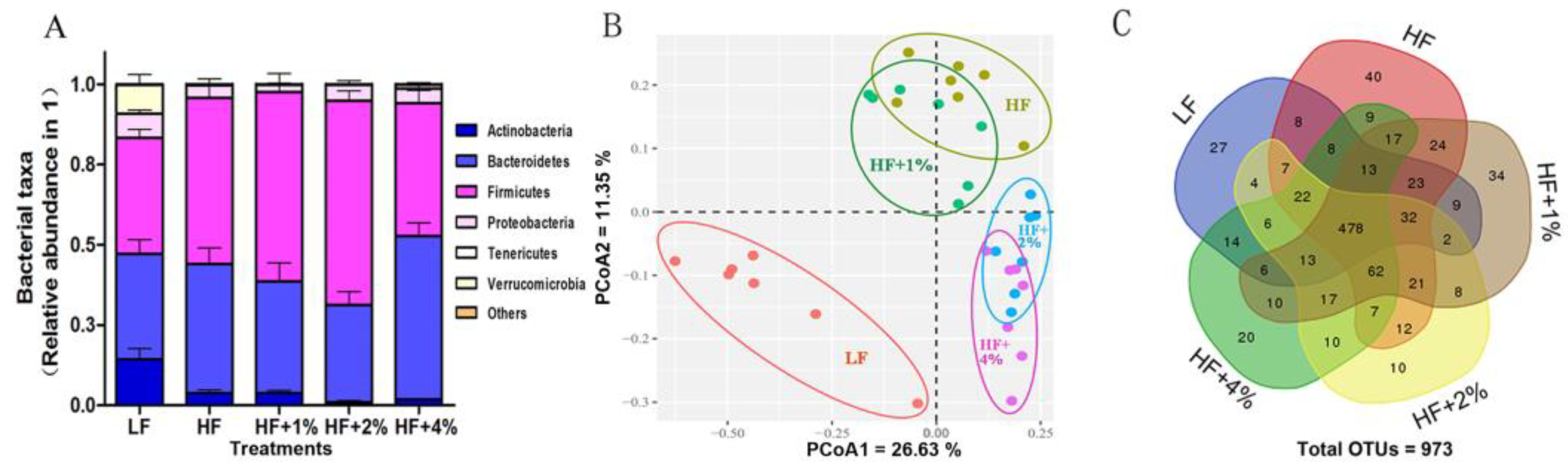
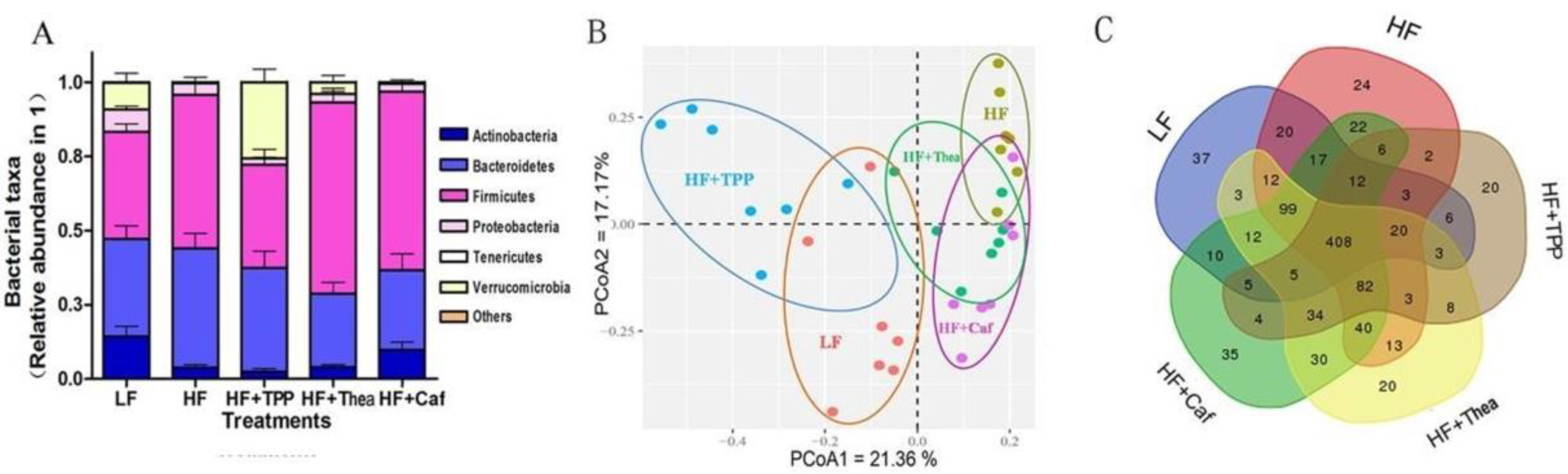
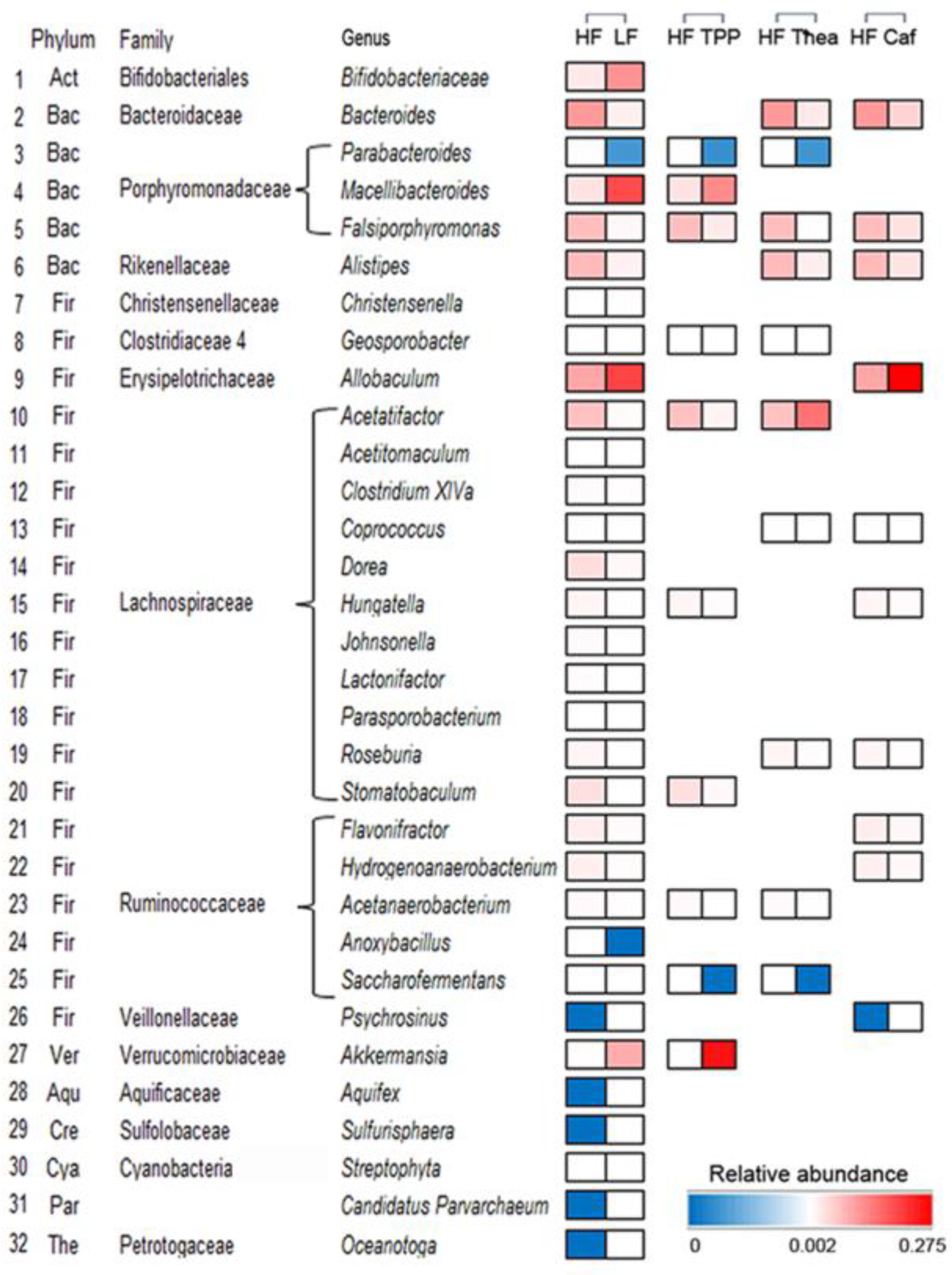
3.2. Effect of Green Tea on HF-Induced Gut Dysbiosis
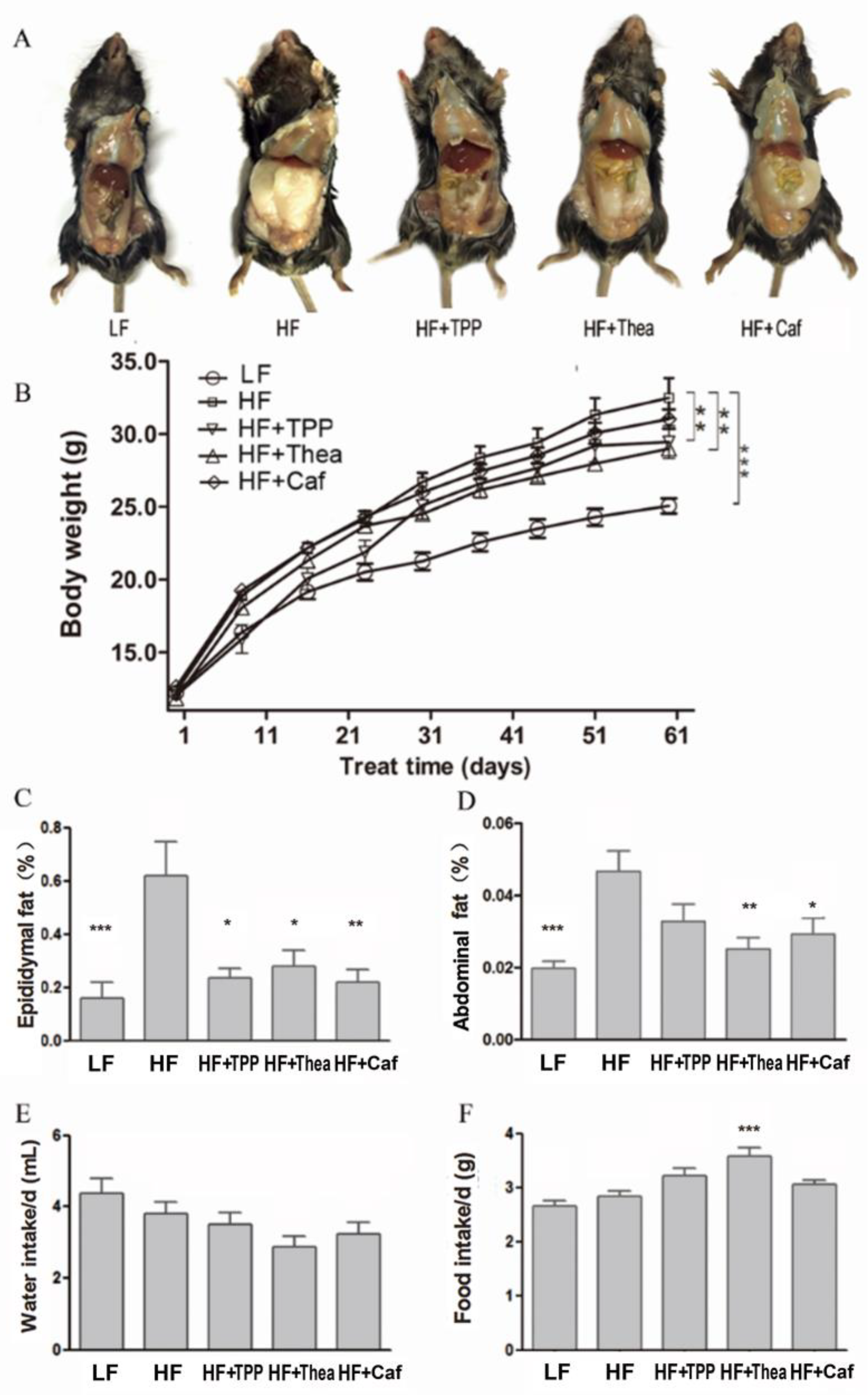
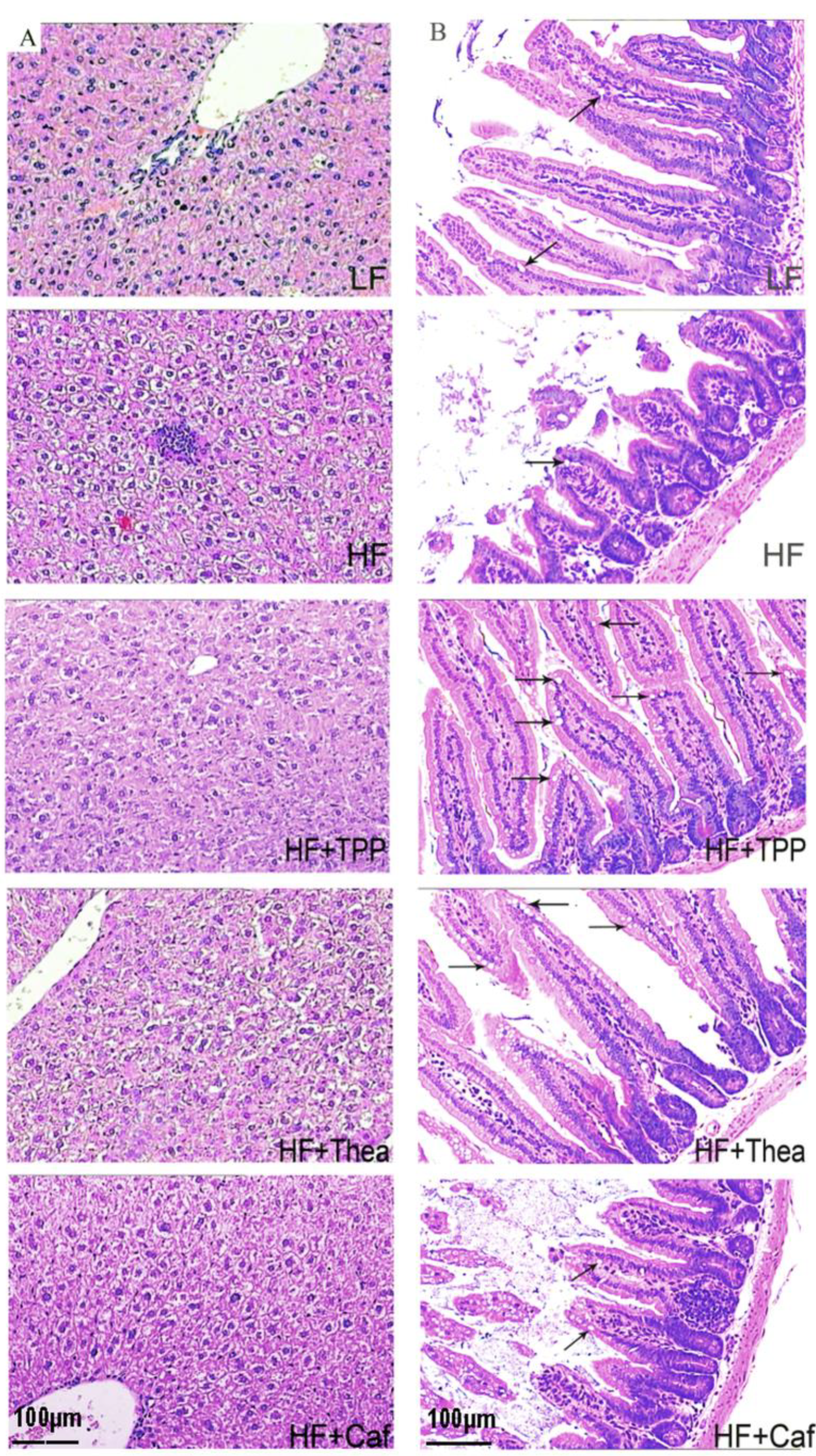
3.3. Influence of TPP, Thea, and Caf on Intestinal Bacteria and Obesity-Related Indices of HF-Induced Obese Mice
3.4. Influence of TPP, Thea, and Caf on Gut Dysbiosis of HF-Induced Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moustafa, J.; Froguel, P. From obesity genetics to the future of personalized obesity therapy. Nat. Rev. Endocrinol. 2013, 9, 402–413. [Google Scholar]
- Lee, H.; Jenner, A.; Low, C.; Lee, Y. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Kahn, R.; Buse, J.; Ferrannini, E.; Stern, M. The metabolic syndrome. Lancet 2005, 366, 1921–1922. [Google Scholar] [CrossRef]
- Ford, E.S. Prevalence of the metabolic syndrome defined by the international diabetes federation among adults in the U.S. Diabetes Care 2005, 28, 2745–2749. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zhou, J.; Zhang, L.; Liu, P.; Zheng, P.; Gao, S.; Song, C.; Yu, Y.; Gong, Z.; Wan, X. Gut microbiota-mediated improvement of metabolic disorders by qingzhuan tea in high fat diet-fed mice. J. Funct. Foods 2021, 78, 104366. [Google Scholar]
- Compare, D.; Rocco, A.; Zamparelli, M.; Nardone, G. The gut bacteria-driven obesity development. Dig. Dis. 2016, 34, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.; Ley, R.; Mahowald, M.; Magrini, V.; Mardis, E.; Gordon, J. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Cani, P.; Bibiloni, R.; Knauf, C.; Neyrinck, A.; Neyrinck, A.; Delzenne, N.; Bu-rcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Hu, S.; Chen, Y.; Zhao, S.; Sun, K.; Luo, L.; Zeng, L. Ripened Pu-Erh tea improved the enterohepatic circulation in a circadian rhythm disorder mice model. J. Agric. Food Chem. 2021, 69, 13533–13545. [Google Scholar] [CrossRef]
- Dumas, M.; Barton, R.; Toye, A.; Cloarec, O.; Blancher, C.; Rothwell, A.; Fearnside, J.; Tatoud, R.; Blanc, V.; Lindon, J.; et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc. Natl. Acad. Sci. USA 2006, 103, 12511–12516. [Google Scholar]
- Hua, F.; Zhou, P.; Liu, P.; Bao, G. Rat plasma protein binding of kaempferol-3-O-rutinoside from Lu’an GuaPian tea and its anti-inflammatory mechanism for cardiovascular protection. J. Food Biochem. 2021, 45, e13749. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.C.; Hu, X.; Li, M.; Rengasamy, K.R.R.; Cai, Y.; Liu, Z. Potential protective function of green tea polyphenol EGCG against high glucose-induced cardiac injury and aging. J. Funct. Foods 2023, 104, 105506. [Google Scholar] [CrossRef]
- Bose, M.; Lambert, J.; Ju, J.; Reuhl, K.; Shapses, S.; Yang, C. The major green tea polyphenol, (-)-Epigallocatechin-3-Gallate, inhibits obesity, metabolic syndrome, and fatty liver disease in high-fat-fed mice. J. Nutr. 2008, 138, 1677–1683. [Google Scholar] [CrossRef]
- Sae-tan, S.; Grove, K.; Lambert, J. Weight control and prevention of metabolic syndrome by green tea. Pharmacol. Res. 2011, 64, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Serisier, S.; Leray, V.; Poudroux, W.; Magot, T.; Ouguerram, K.; Nguyen, P. Effects of green tea on insulin sensitivity, lipid profile and expression of P-PARα and PPARγ and their target genes in obese dogs. Br. J. Nutr. 2008, 99, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhang, J.; Zhang, L.; Huang, J.; Wang, Y. Mechanisms of body weight reduction and metabolic syndrome alleviation by tea. Mol. Nutr. Food Res. 2016, 60, 160–174. [Google Scholar] [CrossRef]
- Wan, C.; Ouyang, J.; Li, M.; Rengasamy, K.; Liu, Z. Effects of green tea polyphenol extract and epigallocatechin-3-o-gallate on diabetes mellitus and diabetic complications: Recent advances. Crit. Rev. Food Sci. Nutr. 2022, 1–29. [Google Scholar] [CrossRef]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.; Queipo-Ortuno, M. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef]
- Seo, D.; Jeong, H.; Cho, D.; Lee, B.; Lee, J.; Choi, J.; Bae, I.; Lee, S. Fermented green tea extract alleviates obesity and related complications and alters gut microbiota composition in diet-induced obese mice. J. Med. Food 2015, 18, 549–556. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, L.; Luo, Y.; Zhang, J.; Wang, X.; Sun, K.; Zeng, L. Prebiotic properties of green and dark tea contribute to protective effects in chemical-induced colitis in mice: A fecal microbiota transplantation study. J. Agric. Food Chem. 2020, 68, 6368–6380. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.; Druart, C.; Bindels, L.; Guiot, Y.; Derrien, M.; Muccioli, G.; Delzenne, N.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Sheng, J.; Yang, F.; Hu, Q. Effect of enzyme inactivation by microwave and oven heating on preservation quality of green tea. J. Food Eng. 2007, 78, 687–692. [Google Scholar] [CrossRef]
- Walk, C.; Cowieson, A.; Remus, J.; Novak, C.; McElroy, A. Effects of dietary enzymes on performance and intestinal goblet cell number of broilers exposed to a live coccidia oocyst vaccine. Poult. Sci. 2011, 90, 91–98. [Google Scholar] [CrossRef]
- Milani, C.; Hevia, A.; Foroni, E.; Duranti, S.; Turroni, F.; Lugli, G.; Sanchez, B.; Martin, R.; Gueimonde, M.; van Sinderen, D.; et al. Assessing the fecal microbiota: An optimized ion torrent 16S rRNA gene-based analysis protocol. PLoS ONE 2013, 8, e68739. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, Q.; Zhou, J.; Wei, Q. Illumina amplicon sequencing of 16S rRNA tag reveals bacterial community development in the rhizosphere of apple nurseries at a replant disease site and a new planting site. PLoS ONE 2014, 9, e111744. [Google Scholar] [CrossRef]
- Caporaso, J.; Lauber, C.; Walters, W.; Berg-Lyons, D.; Lozupone, C.; Turnbaugh, P.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Saleh, I.; Ali, Z.; Abe, N.; Wilson, F.; Hamada, F.; Abd-Ellah, M.; Walker, L.; Khan, I.; Ashfaq, M. Effect of green tea and its polyphenols on mouse liver. Fitoterapia 2013, 90, 151–159. [Google Scholar] [CrossRef]
- Saleh, I.A.; Hamada, F.M.; Abd-Ellah, M.F.; Khan, I.A.; Walker, L.A.; Ashfaq, M.K. Consumption of EGCG can become hepatotoxic under inflammatory condition. Planta Med. 2013, 79, P95. [Google Scholar] [CrossRef]
- van Duynhoven, J.; Vaughan, E.; Jacobs, D.; Kemperman, R.; van Velzen, E.; Gross, G.; Roger, L.; Possemiers, S.; Smilde, A.; Dore, J.; et al. Metabolic fate of polyphenols in the human superorganism. Proc. Natl. Acad. Sci. USA 2011, 108, 4531–4538. [Google Scholar] [CrossRef] [PubMed]
- Higdon, J.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef] [PubMed]
- Candela, M.; Maccaferri, S.; Turroni, S.; Carnevali, P.; Brigidi, P. Functional intestinal microbiome, new frontiers in prebiotic design. Int. J. Food Microbiol. 2010, 140, 93–101. [Google Scholar] [CrossRef]
- Cheng, M.; Zhang, X.; Miao, Y.; Cao, J.; Wu, Z.; Weng, P. The modulatory effect of (-)-epigallocatechin 3-O-(3-O-methyl) gallate (EGCG3″Me) on intestinal microbiota of high fat diet-induced obesity mice model. Food Res. Int. 2017, 92, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Cheng, M.; Zhang, X.; Cao, J.; Wu, Z.; Weng, P. Green tea polyphenols reduce obesity in high-fat diet-induced mice by modulating intestinal microbiota composition. Int. J. Food Sci. Technol. 2017, 52, 1723–1730. [Google Scholar] [CrossRef]
- Henning, S.; Yang, J.; Hsu, M.; Lee, R.; Grojean, E.; Ly, A.; Tseng, C.; Heber, D.; Li, Z. Decaffeinated green and black tea polyphenols decrease weight gain and alter microbiome populations and function in diet-induced obese mice. Eur. J. Nutr. 2018, 57, 2759–2769. [Google Scholar] [CrossRef]
- Janssens, P.L.H.R.; Penders, J.; Hursel, R.; Budding, A.E.; Savelkoul, P.H.M.; Westerterp-Plantenga, M.S. Long-term green tea supplementation does not change the human gut microbiota. PLoS ONE 2016, 11, e0153134. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, Z.; Guo, H.; He, D.; Zhao, H.; Wang, Z.; Zhang, W.; Liao, L.; Zhang, C.; Ni, L. The modulatory effect of infusions of green tea, oolong tea, and black tea on gut microbiota in high-fat-induced obese mice. Food Funct. 2016, 7, 4869–4879. [Google Scholar] [CrossRef]
- Singh, D.; Singh, J.; Boparai, R.; Zhu, J.; Mantri, S.; Khare, P.; Khardori, R.; Kondepudi, K.; Chopra, K.; Bishnoi, M. Isomalto-oligosaccharides, a prebiotic, functionally augment green tea effects against high fat diet-induced metabolic alterations via preventing gut dysbacteriosis in mice. Pharmacol. Res. 2017, 123, 103–113. [Google Scholar] [CrossRef]
- Anhe, F.; Roy, D.; Pilon, G.; Dudonne, S.; Matamoros, S.; Varin, T.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef]
- Kemperman, R.; Gross, G.; Mondot, S.; Possemiers, S.; Marzorati, M.; Van de Wiele, T.; Dore, J.; Vaughan, E. Impact of polyphenols from black tea and red wine/grape juice on a gut model microbiome. Food Res. Int. 2013, 53, 659–669. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, X.; Cao, S.; Cromie, M.; Shen, Y.; Feng, Y.; Yang, H.; Li, L. Chlorogenic acid ameliorates experimental colitis by promoting growth of Akkermansia in mice. Nutrients 2017, 9, 677. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xue, Y.; Zhang, H.; Du, M.; Zhu, M. Favourable effects of grape seed extract on intestinal epithelial differentiation and barrier function in IL10-deficient mice. Br. J. Nutr. 2015, 114, 15–23. [Google Scholar] [CrossRef][Green Version]
- Martin, A.R.; La Casa, C.; Villegas, I.; Herrerias, J.M.; Alarcon de la Lastra, C. Resveratrol, a polyphenol found in grapes, suppresses oxidative damage and stimulates apoptosis during early colonic inflammation in rats. Biochem. Pharmacol. 2004, 67, 1399–1410. [Google Scholar]
- Roopchand, D.; Carmody, R.; Kuhn, P.; Moskal, K.; Rojas-Silva, P.; Turnbaugh, P.; Raskin, I. Dietary polyphenols promote growth of the gut bacterium Akkermansia muciniphila and attenuate high-fat diet-induced metabolic syndrome. Diabetes 2015, 64, 2847–2858. [Google Scholar] [CrossRef]
- Min, B.R.; Perkins, D.; Wright, C.; Dawod, A.; Min, B.J.; Terrill, T.H. Effects of feeding two different tannin-containing diets on ruminal fermentation profiles and microbial community changes in meat goats. Agric. Food Anal. Bacteriol. 2015, 5, 153–165. [Google Scholar]
- Ravussin, Y.; Koren, O.; Spor, A.; LeDuc, C.; Gutman, R.; Stombaugh, J.; Knight, R.; Ley, R.; Leibel, R. Responses of gut microbiota to diet composition and weight loss in lean and obese mice. Obesity 2012, 20, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.; Wallace, G.; Zhang, C.; Legge, R.; Benson, A.; Carr, T.; Moriyama, E.; Walter, J. Diet-induced metabolic improvements in a hamster model of hypercholesterolemia are strongly linked to alterations of the gut microbiota. Appl. Environ. Microbiol. 2009, 75, 4175–4184. [Google Scholar] [CrossRef]
- Rehman, A.; Sina, C.; Gavrilova, O.; Hasler, R.; Ott, S.; Baines, J.; Schreiber, S.; Rosenstiel, P. Nod2 is essential for temporal development of intestinal microbial communities. Gut 2011, 60, 1354–1362. [Google Scholar] [CrossRef]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut microbiome development along the colorectal adenoma-carcinoma sequence. Nat. Commun. 2015, 6, 6528. [Google Scholar] [CrossRef]
- Xie, G.; Wang, X.; Liu, P.; Wei, R.; Chen, W.; Rajani, C.; Hernandez, B.; Alegado, R.; Dong, B.; Li, D.; et al. Distinctly altered gut microbiota in the progression of liver disease. Oncotarget 2016, 7, 19355–19366. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Huang, J.; Fu, D.; Li, Y.; Liu, Z. Tea loses weight and its preventive effect on metabolic syndrome in the human body. Chinatea 2019, 5, 7–13. [Google Scholar]
- Janssens, P.L.H.R.; Hursel, R.; Westerterp-Plantenga, M.S. Nutraceuticals for body-weight management: The role of green tea catechins. Physiol. Behav. 2016, 162, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xie, M.; Dai, Z.; Wan, P.; Ye, H.; Zeng, X.; Sun, Y. Kudingcha and Fuzhuan brick tea prevent obesity and modulate gut microbiota in high-fat diet fed mice. Mol. Nutr. Food Res. 2018, 62, 1700485. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component (gL−1) | EC | EGC | ECG | EGCG | C | GC | GCG | CG | TPP | Caf | Thea |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 4% green tea | 0.23 | 0.61 | 0.69 | 1.91 | 0.07 | 0.34 | 0.31 | 0.18 | 6.07 | 1.52 | 1.50 |
| 6 gL−1 TPP | 0.15 | 0.01 | 0.55 | 2.35 | 0.01 | 0.22 | 0.36 | 0.14 | 6.00 | - | - |
| Treatment | LF | HF | HF+1% | HF+2% | HF+4% |
|---|---|---|---|---|---|
| TC (mmol/L) | 3.76 ± 0.14 *** | 6.05 ± 0.34 | 5.77 ± 0.30 | 6.18 ± 0.14 | 4.85 ± 0.17 ** |
| HDL (mmol/L) | 2.00 ± 0.27 | 1.73 ± 0.18 | 2.50 ± 0.21 * | 2.08 ± 0.10 | 2.25 ± 0.01 |
| LDL (mmol/L) | 0.84 ± 0.12 *** | 2.16 ± 0.25 | 1.66 ± 0.08 * | 1.93 ± 0.08 | 1.49 ± 0.06 ** |
| FBG (mmol/L) | 4.87 ± 0.19 *** | 7.67 ± 0.20 | 6.44 ± 0.38 * | 4.17 ± 0.30 *** | 7.01 ± 0.46 |
| INS (mIU/L) | 11.62 ± 1.35 | 9.69 ± 1.18 | 16.07 ± 2.39 * | 17.18 ± 1.40 * | 13.18 ± 0.83 |
| ALT (Kar U) | 4.83 ± 1.67 * | 11.92 ± 2.43 | 4.89 ± 1.38 * | 2.83 ± 1.01 ** | 1.95 ± 1.00 ** |
| AST (Kar U) | 148.00 ± 12.21 | 148.20 ± 4.54 | 90.22 ± 8.09 ** | 120.30 ± 18.39 | 111.20 ± 12.99 * |
| LPS (μg/L) | 207.90 ± 12.38 | 245.90 ± 12.84 | 239.10 ± 21.94 | 234.00 ± 11.44 | 237.90 ± 8.12 |
| TNF-α (ng/L) | 243.60 ± 23.51 * | 309.10 ± 17.32 | 299.50 ± 8.29 | 309.00 ± 19.94 | 275.80 ± 6.95 * |
| MCP-1 (ng/L) | 254.60 ± 33.97 * | 193.90 ± 15.58 | 222.50 ± 37.57 | 294.50 ± 42.64 ** | 186.10 ± 48.32 |
| TG (mmol/g pro) | 0.13 ± 0.01 ** | 0.23 ± 0.02 | 0.20 ± 0.02 | 0.13 ± 0.01 ** | 0.18 ± 0.02 |
| FFA (mmol/g pro) | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.07 ± 0.01* | 0.06 ± 0.02 ** | 0.06 ± 0.01 * |
| Treatment | LF | HF | HF + TPP | HF + Thea | HF + Caf |
|---|---|---|---|---|---|
| TC (mmol/L) | 3.76 ± 0.14 *** | 6.05 ± 0.34 | 5.09 ± 0.39 | 4.54 ± 0.32 ** | 5.59 ± 0.30 |
| HDL (mmol/L) | 2.00 ± 0.27 | 1.73 ± 0.18 | 2.75 ± 0.16 ** | 2.35 ± 0.19 | 2.36 ± 0.18 |
| LDL (mmol/L) | 0.84 ± 0.12 *** | 2.16 ± 0.25 | 1.56 ± 0.10 * | 1.53 ± 0.07 * | 1.61 ± 0.27 * |
| FBG (mmol/L) | 4.87 ± 0.19 *** | 7.67 ± 0.20 | 7.93 ± 0.24 | 8.02 ± 0.36 | 8.10 ± 0.21 |
| INS (mIU/L) | 11.62 ± 1.35 | 9.69 ± 1.18 | 10.17 ± 0.90 | 10.07 ± 2.07 | 12.79 ± 1.29 |
| ALT (Kar U) | 4.83 ± 1.67 * | 11.92 ± 2.43 | 12.55 ± 1.39 | 13.80 ± 0.82 | 11.30 ± 1.15 |
| AST (Kar U) | 148.00 ± 12.21 | 148.20 ± 4.54 | 70.55 ± 9.18 *** | 84.82 ± 9.73 *** | 103.1 ± 10.39 * |
| LPS (μg/L) | 207.90 ± 12.38 | 245.90 ± 12.84 | 186.00 ± 8.61 * | 241.34 ± 15.66 | 224.36 ± 16.95 |
| TNF-α (ng/L) | 243.60 ± 23.51 * | 309.10 ± 17.32 | 250.97 ± 11.29 * | 284.53 ± 21.73 | 292.21 ± 16.61 |
| MCP-1 (ng/L) | 254.60 ± 33.97 * | 193.90 ± 15.58 | 197.21 ± 19.30 | 270.79 ± 42.42 | 247.65 ± 36.67 |
| TG (mmol/g pro) | 0.13 ± 0.01 ** | 0.23 ± 0.02 | 0.14 ± 0.01 ** | 0.17 ± 0.02 | 0.19 ± 0.03 |
| FFA (mmol/g pro) | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.06 ± 0.01 ** | 0.09 ± 0.01 | 0.06 ± 0.01 ** |
| Treatment | LF | HF | HF + TPP | HF + Thea | HF + Caf |
|---|---|---|---|---|---|
| Goblet cell/villus | 5.36 ± 1.51 | 2.80 ± 0.33 | 27.67 ± 4.59 *** | 24.29 ± 7.08 *** | 10.44 ± 3.00 |
| Villus height (μm) | 347.50 ± 29.07 | 289.60 ± 29.71 | 390.50 ± 33.02 * | 411.30 ± 22.99 * | 248.70 ± 14.56 |
| Crypt depth (μm) | 134.40 ± 4.65 *** | 90.55 ± 5.64 | 133.40 ± 11.19 *** | 118.30 ± 4.52 * | 119.80 ± 4.78 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, H.; Li, J.; Liu, S.; Jeyaraj, A.; Zhuang, J.; Wang, Y.; Chen, X.; Yuan, Q.; Li, X. The Role of Green Tea on the Regulation of Gut Microbes and Prevention of High-Fat Diet-Induced Metabolic Syndrome in Mice. Foods 2023, 12, 2953. https://doi.org/10.3390/foods12152953
Mei H, Li J, Liu S, Jeyaraj A, Zhuang J, Wang Y, Chen X, Yuan Q, Li X. The Role of Green Tea on the Regulation of Gut Microbes and Prevention of High-Fat Diet-Induced Metabolic Syndrome in Mice. Foods. 2023; 12(15):2953. https://doi.org/10.3390/foods12152953
Chicago/Turabian StyleMei, Huiling, Jin Li, Shujing Liu, Anburaj Jeyaraj, Jing Zhuang, Yuhua Wang, Xuan Chen, Qijun Yuan, and Xinghui Li. 2023. "The Role of Green Tea on the Regulation of Gut Microbes and Prevention of High-Fat Diet-Induced Metabolic Syndrome in Mice" Foods 12, no. 15: 2953. https://doi.org/10.3390/foods12152953
APA StyleMei, H., Li, J., Liu, S., Jeyaraj, A., Zhuang, J., Wang, Y., Chen, X., Yuan, Q., & Li, X. (2023). The Role of Green Tea on the Regulation of Gut Microbes and Prevention of High-Fat Diet-Induced Metabolic Syndrome in Mice. Foods, 12(15), 2953. https://doi.org/10.3390/foods12152953