
Molecular and Genetic Characterization of Colicinogenic Escherichia coli Strains Active against Shiga Toxin-Producing Escherichia coli O157:H7

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Origin and Identification of E. coli and Indicator Strains
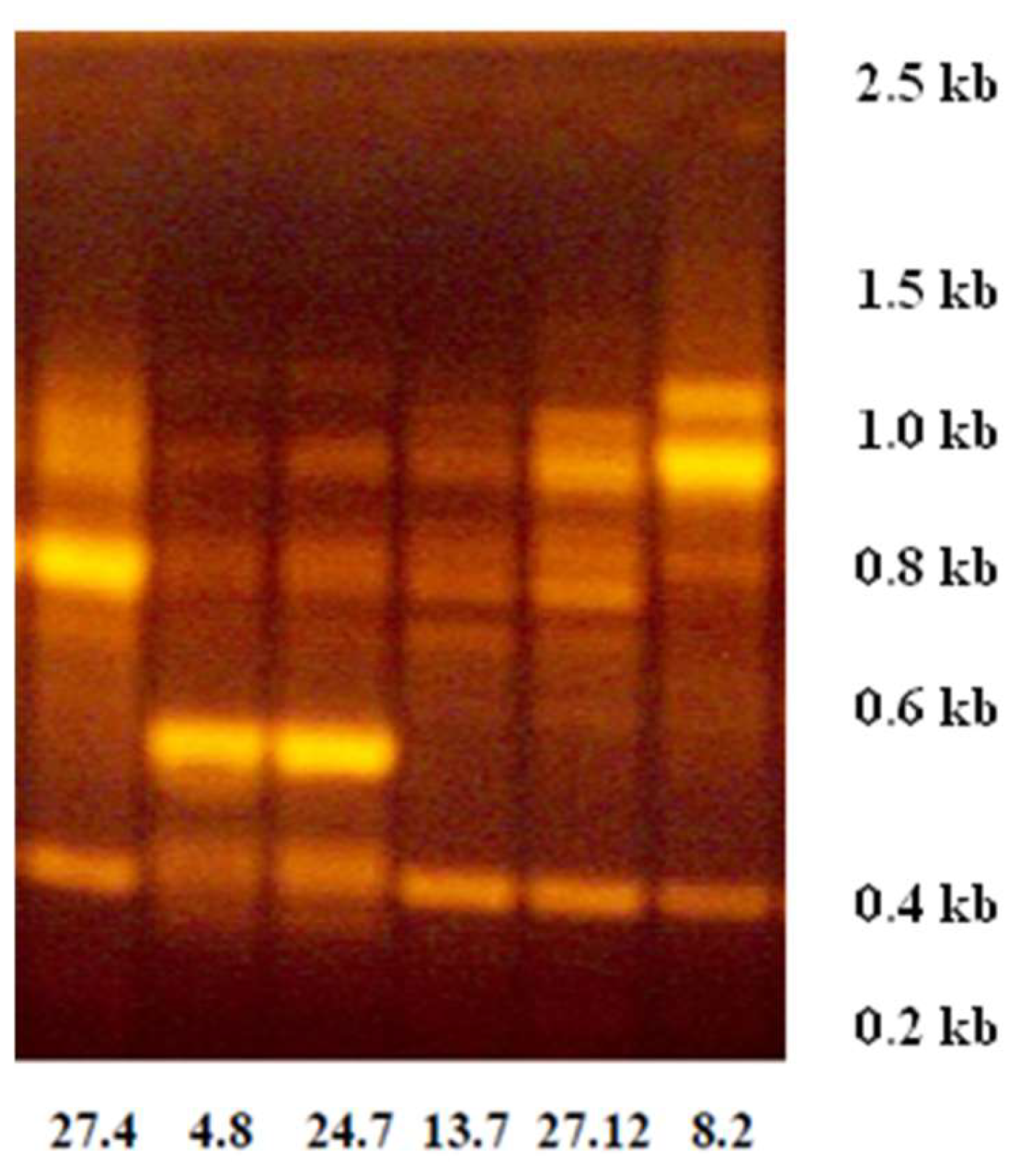
2.1.1. Genetic Profiles by Random Amplification of Polymorphic DNA (RAPD)
2.1.2. Serotyping
2.1.3. Origin of Indicator Strains
2.2. Detection of Colicin Genes
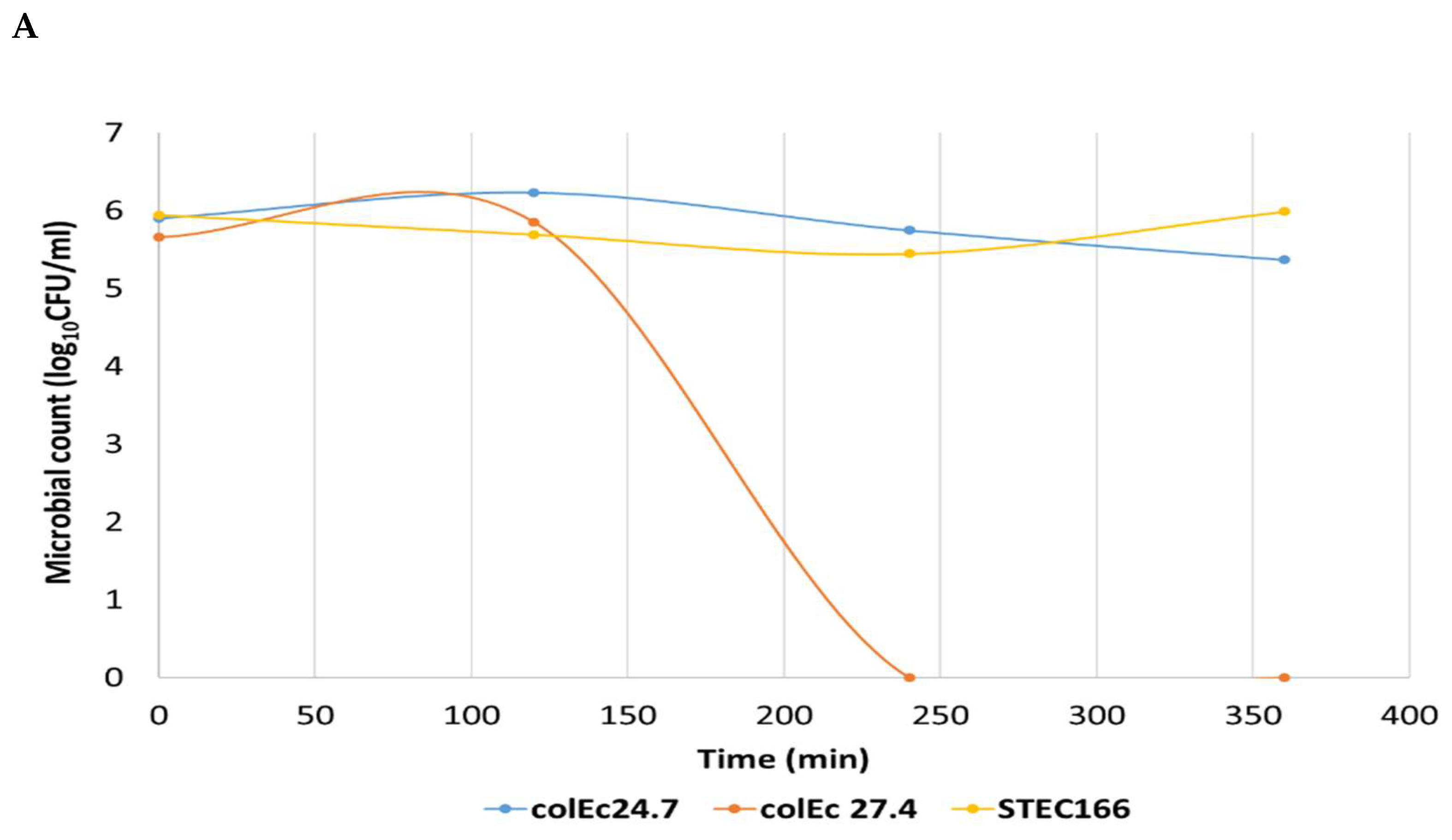
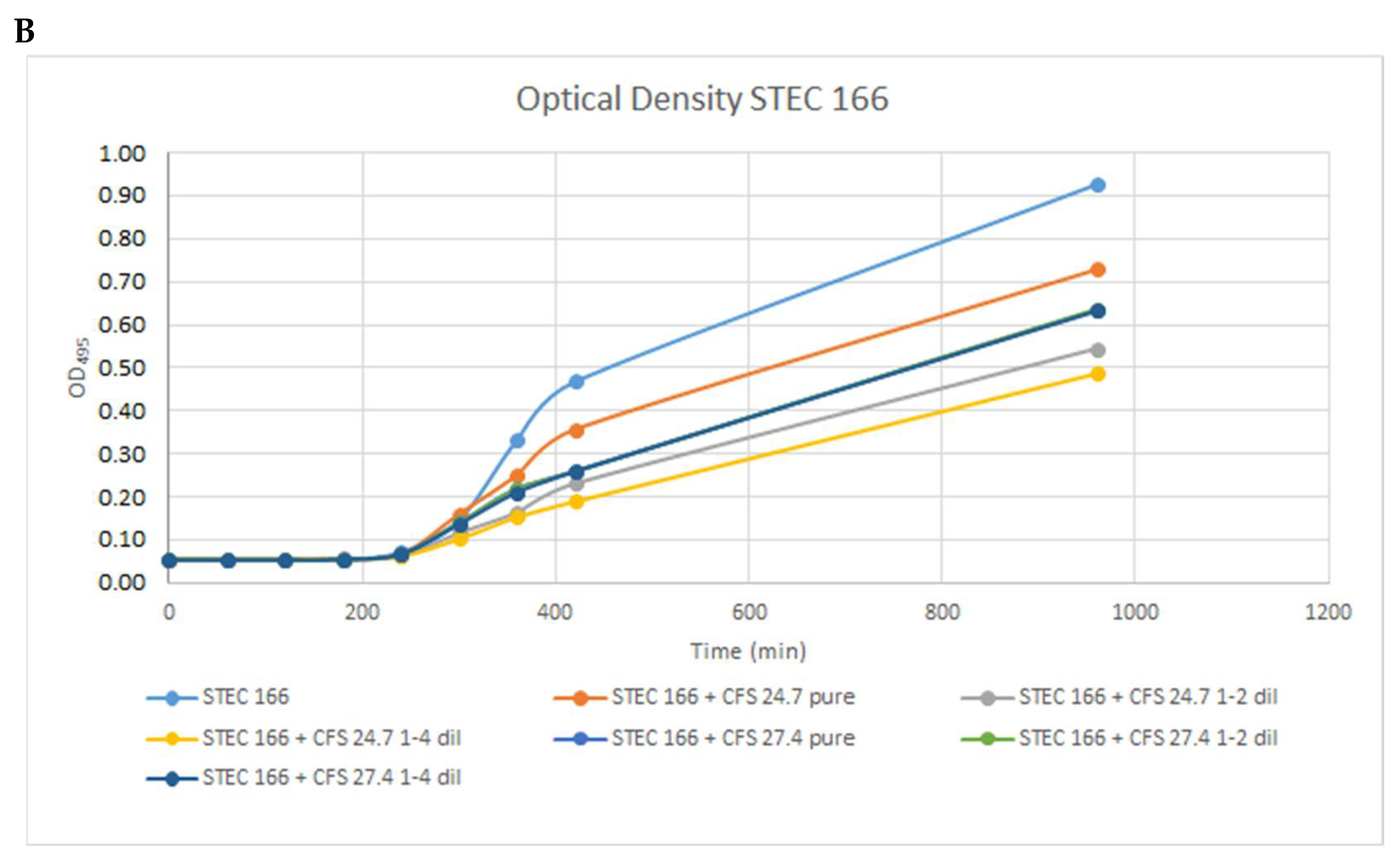
2.3. Determination of Inhibitory Activity of ColEc against STEC Strains
Determination of Inhibitory Activity of Colicins against STEC 166
2.4. Differentiation Assay of Growth Inhibition by Phage or Colicin
2.5. Stability of CFS to Physic-Chemical Treatments
- I.
- Temperature: The CFS extracts were heated in a thermal bath at 60 °C, 80 °C, and 100 °C; they were frozen at −20 and kept at 4 °C [38].
- II.
- pH changes: The pH of the CFS extracts were adjusted to approximately 4.5, 7.0, and 9.0 using HCl (Biopack 30%) and NaOH (Biopack 10N) (Larsen et al., 1993).
- III.
- Stability to protease: CFS extracts were incubated with Proteinase K (Sigma-Aldricht 20 ng/µL) at 37 °C for 15–60 min.
2.6. Virulence Genes Detection
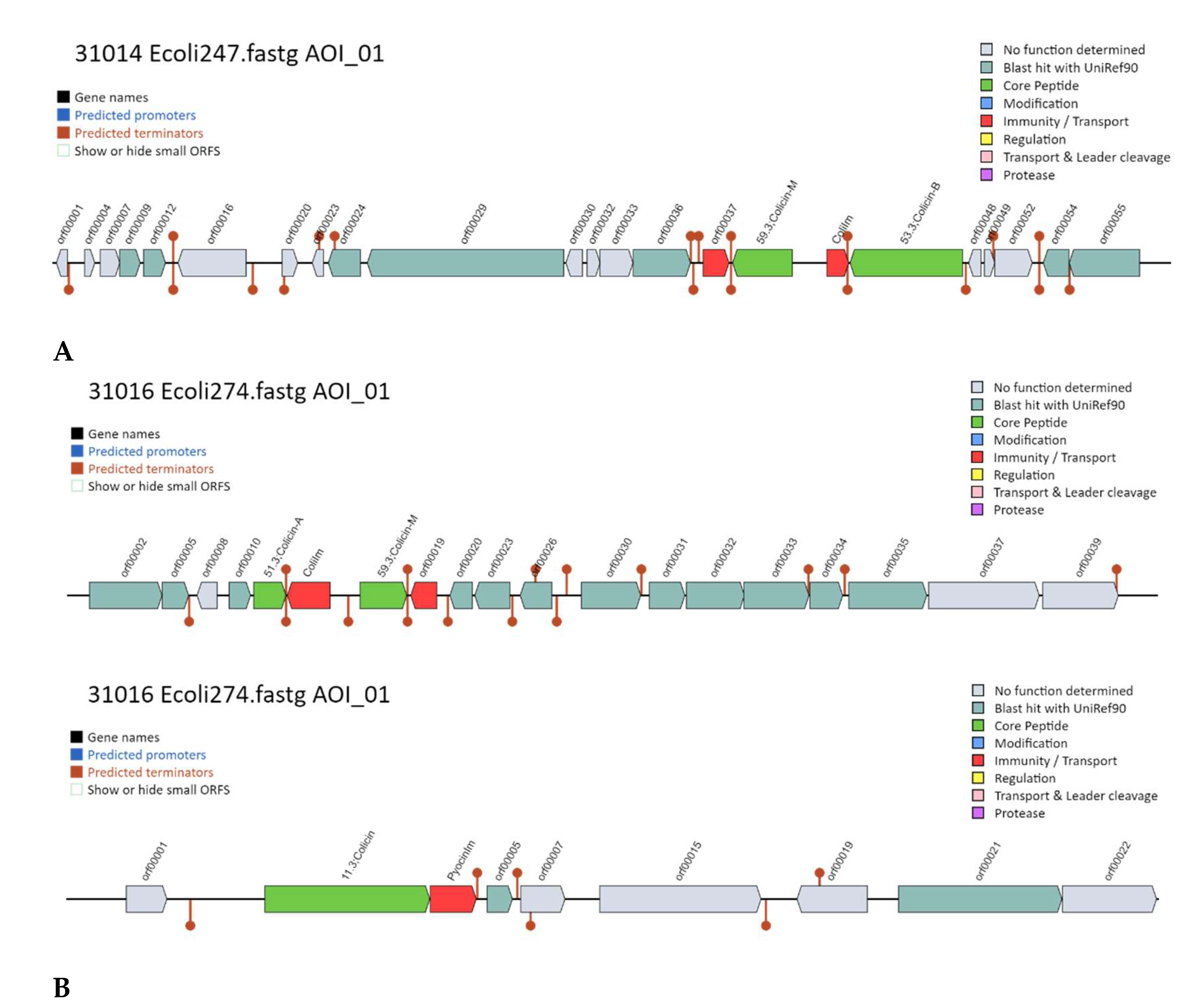
2.7. Strains Sequencing: Bacteriocin Mining
3. Results
3.1. Genetic Profile and Serotype Determination
3.2. PCR Screening
3.3. Inhibition Assays
3.4. Differentiation Assay of Growth Inhibition by Phage or Colicin
3.5. Stability Assays
3.6. Virulence Finder
3.7. Whole Genome Sequencing and Annotation: Bacteriocin Mining
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Rubio, L.; Haarmann, N.; Schwidder, M.; Muniesa, M.; Schmidt, H. Bacteriophages of Shiga Toxin-Producing E. coli and Their Contribution to Pathogenicity. Pathogens 2021, 10, 404. [Google Scholar] [CrossRef]
- Bai, X.; Ylinen, E.; Zhang, J.; Salmenlinna, S.; Halkilahti, J.; Saxen, H.; Narayanan, A.; Jahnukainen, T.; Matussek, A. Comparative Genomics of Shiga Toxin-Producing E. coli Strains Isolated from Pediatric Patients with and without Hemolytic Uremic Syndrome from 2000 to 2016 in Finland. Microbiol. Spectr. 2022, 10, e00660-22. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.; Chinen, I.; Guth, B.E.C. Enterohemorrhagic (Shiga Toxin-Producing) Escherichia Coli. In E. coli in the Americas; Torres, A.G., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 97–123. ISBN 978-3-319-45092-6. [Google Scholar]
- FAO/OMS. Informe de la República Argentina en Respuesta a la Solicitud de Datos Sobre E. coli Productor de Verocitotoxina (VTEC)/E. coli Productor de Toxina de Shiga (STEC). 2016. Available online: https://rsa.conicet.gov.ar/wp-content/uploads/2016/07/Informe-FAO-OMS-Escherichia-coli-VTEC-STEC-Argentina.pdf (accessed on 1 April 2021).
- Bell, B.P.; Goldoft, M.; Griffin, P.M.; Davis, M.A.; Gordon, D.C.; Tarr, P.I.; Bartleson, C.A.; Lewis, J.H.; Barrett, T.J.; Wells, J.G.; et al. A Multistate Outbreak of E. coli O157:H7—Associated Bloody Diarrhea and Hemolytic Uremic Syndrome from Hamburgers: The Washington Experience. JAMA 1994, 272, 1349–1353. [Google Scholar] [CrossRef]
- Fernández, D.; Sanz, M.E.; Parma, A.E.; Padola, N.L. Short Communication: Characterization of Shiga Toxin-Producing E. coli Isolated from Newborn, Milk-Fed, and Growing Calves in Argentina. J. Dairy Sci. 2012, 95, 5340–5343. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, E.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Gut Microbiota as a Source of Novel Antimicrobials. Gut Microbes 2019, 10, 1–21. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial Production of Bacteriocins: Latest Research Development and Applications. Biotechnol. Adv. 2018, 36, 2187–2200. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.T.; Wu, J.Q.; Xie, F.; Hu, S.H.; Mo, Y. Efficacy of Nisin in Treatment of Clinical Mastitis in Lactating Dairy Cows. J. Dairy Sci. 2007, 90, 3980–3985. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.G.; Lai, Z.W.; Tan, J.S. Bacteriocins from Lactic Acid Bacteria: Purification Strategies and Applications in Food and Medical Industries: A Review. Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 51. [Google Scholar] [CrossRef]
- Piper, C.; Cotter, P.D.; Ross, R.P.; Hill, C. Discovery of Medically Significant Lantibiotics. Curr. Drug Discov. Technol. 2009, 6, 1–18. [Google Scholar] [CrossRef]
- Šmajs, D.; Weinstock, G.M. Genetic Organization of Plasmid ColJs, Encoding Colicin Js Activity, Immunity, and Release Genes. J. Bacteriol. 2001, 183, 3949–3957. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cascales, E.; Buchanan, S.K.; Duché, D.; Kleanthous, C.; Lloubès, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin Biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef] [PubMed]
- Micenková, L.; Štaudová, B.; Bosák, J.; Mikalová, L.; Littnerová, S.; Vrba, M.; Ševčíková, A.; Woznicová, V.; Šmajs, D. Bacteriocin-Encoding Genes and ExPEC Virulence Determinants Are Associated in Human Fecal E. coli Strains. BMC Microbiol. 2014, 14, 109. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.A.; Chavan, M.A. (Eds.) Bacteriocins: Ecology and Evolution; Springer: Berlin, Germany; New York, NY, USA, 2007; ISBN 978-3-540-36603-4. [Google Scholar]
- Corsini, G.; Baeza, M.; Monasterio, O.; Lagos, R. The Expression of Genes Involved in Microcin Maturation Regulates the Production of Active Microcin E492. Biochimie 2002, 84, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Majeed, H.; Ghazaryan, L.; Herzberg, M.; Gillor, O. Bacteriocin Expression in Sessile and Planktonic Populations of Escherichia Coli. J. Antibiot. 2015, 68, 52–55. [Google Scholar] [CrossRef]
- Mader, A.; von Bronk, B.; Ewald, B.; Kesel, S.; Schnetz, K.; Frey, E.; Opitz, M. Amount of Colicin Release in E. coli Is Regulated by Lysis Gene Expression of the Colicin E2 Operon. PLoS ONE 2015, 10, e0119124. [Google Scholar] [CrossRef]
- Butala, M.; Sonjak, S.; Kamenšek, S.; Hodošček, M.; Browning, D.F.; Žgur-Bertok, D.; Busby, S.J.W. Double Locking of an E. coli Promoter by Two Repressors Prevents Premature Colicin Expression and Cell Lysis. Mol. Microbiol. 2012, 86, 129–139. [Google Scholar] [CrossRef]
- Monroy-Dasta, M.C.; Castro-Barrera, T.; Fernández-Perrino, F.J. Revisión Bibliográfica: Bacteriocinas Producidas Por Bacterias Probióticas. Available online: https://scholar.googleusercontent.com/scholar?q=cache:u-MgnYaDE8oJ:scholar.google.com/+18.%09Monroy-Dasta,+M.C.%3B+Castro-Barrera,+T.%3B+Fern%C3%A1ndez-Perrino,+F.J.+%E2%80%9CBacteriocinas+producidas+por+bacterias+probi%C3%B3ticas%E2%80%9D.+ContactoS.+2009,+73,+63%E2%80%9372.&hl=es&as_sdt=0,5 (accessed on 23 November 2022).
- Ruiz, M.J.; Colello, R.; Padola, N.L.; Etcheverría, A.I. Efecto inhibitorio de Lactobacillus spp. sobre bacterias implicadas en enfermedades transmitidas por alimentos. Rev. Argent. De Microbiol. 2017, 49, 174–177. [Google Scholar] [CrossRef]
- Kumar, P. Isolation and Characterization of Potential Probiotic E. coli Strains from Rat Faecal Samples. Am. J. Infect. Dis. 2009, 5, 112–117. [Google Scholar] [CrossRef]
- von Buenau, R.; Jaekel, L.; Schubotz, E.; Schwarz, S.; Stroff, T.; Krueger, M. E. coli Strain Nissle 1917: Significant Reduction of Neonatal Calf Diarrhea. J. Dairy Sci. 2005, 88, 317–323. [Google Scholar] [CrossRef]
- Schneider, T.; Hahn-Löbmann, S.; Stephan, A.; Schulz, S.; Giritch, A.; Naumann, M.; Kleinschmidt, M.; Tusé, D.; Gleba, Y. Plant-Made Salmonella Bacteriocins Salmocins for Control of Salmonella Pathovars. Sci. Rep. 2018, 8, 4078. [Google Scholar] [CrossRef]
- Schulz, S.; Stephan, A.; Hahn, S.; Bortesi, L.; Jarczowski, F.; Bettmann, U.; Paschke, A.-K.; Tusé, D.; Stahl, C.H.; Giritch, A.; et al. Broad and Efficient Control of Major Foodborne Pathogenic Strains of E. coli by Mixtures of Plant-Produced Colicins. Proc. Natl. Acad. Sci. USA 2015, 112, E5454–E5460. [Google Scholar] [CrossRef] [PubMed]
- Bosák, J.; Hrala, M.; Micenková, L.; Šmajs, D. Non-Antibiotic Antibacterial Peptides and Proteins of Escherichia Coli: Efficacy and Potency of Bacteriocins. Expert Rev. Anti-Infect. Ther. 2021, 19, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Lozo, J.; Vukasinovic, M.; Strahinic, I.; Topisirovic, L. Characterization and Antimicrobial Activity of Bacteriocin 217 Produced by Natural Isolate Lactobacillus paracasei Subsp. paracasei BGBUK2-16. J. Food Prot. 2004, 67, 2727–2734. [Google Scholar] [CrossRef]
- Dolejšová, T.; Sokol, A.; Bosák, J.; Šmajs, D.; Konopásek, I.; Mikušová, G.; Fišer, R. Colicin U from Shigella Boydii Forms Voltage-Dependent Pores. J. Bacteriol. 2019, 201, e00493-19. [Google Scholar] [CrossRef]
- Fathizadeh, H.; Saffari, M.; Esmaeili, D.; Moniri, R.; Salimian, M. Evaluation of Antibacterial Activity of Enterocin A-Colicin E1 Fusion Peptide. Iran. J. Basic Med. Sci. 2020, 23, 1471–1479. [Google Scholar] [CrossRef]
- Stephan, A.; Hahn-Löbmann, S.; Rosche, F.; Buchholz, M.; Giritch, A.; Gleba, Y. Simple Purification of Nicotiana Benthamiana-Produced Recombinant Colicins: High-Yield Recovery of Purified Proteins with Minimum Alkaloid Content Supports the Suitability of the Host for Manufacturing Food Additives. Int. J. Mol. Sci. 2018, 19, 95. [Google Scholar] [CrossRef] [PubMed]
- Etcheverría, A.I.; Arroyo, G.H.; Perdigon, G.; Parma, A.E. E. coli with Anti-O157:H7 Activity Isolated from Bovine Colon. J. Appl. Microbiol. 2006, 100, 384–389. [Google Scholar] [CrossRef]
- Birch, M.; Denning, D.W.; Law, D. Rapid Genotyping of E. coli O157 Isolates by Random Amplification of Polymorphic DNA. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 297–302. [Google Scholar] [CrossRef]
- Guinée, P.A.M.; Jansen, W.H.; Wadström, T.; Sellwood, R. E. coli Associated with Neonatal Diarrhoea in Piglets and Calves. In Laboratory Diagnosis in Neonatal Calf and Pig Diarrhoea: Proceedings of a Workshop on Diagnostic Techniques for Enteropathogenic Agents Associated with Neonatal Diarrhoea in Calves and Pigs, Central Veterinary Institute, Department of Virology, Lelystad, The Netherlands, June 3–5, 1980; de Leeuw, P.W., Guinée, P.A.M., Eds.; Current Topics in Veterinary Medicine and Animal Science; Springer Netherlands: Dordrecht, The Netherlands, 1981; pp. 126–162. ISBN 978-94-009-8328-1. [Google Scholar]
- Blanco, J.E.; Blanco, M.; Blanco, J.; Mora, A.; Balaguer, L.; Mouriño, M.; Juarez, A.; Jansen, W.H. O Serogroups, Biotypes, and Eae Genes in E. coli Strains Isolated from Diarrheic and Healthy Rabbits. J. Clin. Microbiol. 1996, 34, 3101–3107. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Blanco, M.; Alonso, M.P.; Blanco, J.E.; Garabal, J.; González, E.A. Serogroups of E. coli Strains Producing Cytotoxic Necrotizing Factors CNF1 and CNF2. FEMS Microbiol. Lett. 1992, 96, 155–159. [Google Scholar] [CrossRef]
- Pradel, N.; Livrelli, V.; De Champs, C.; Palcoux, J.-B.; Reynaud, A.; Scheutz, F.; Sirot, J.; Joly, B.; Forestier, C. Prevalence and Characterization of Shiga Toxin-Producing E. coli Isolated from Cattle, Food, and Children during a One-Year Prospective Study in France. J. Clin. Microbiol. 2000, 38, 1023–1031. [Google Scholar] [CrossRef]
- Ørskov, F.; Ørskov, I. 2 Serotyping of Escherichia Coli**The Terminology Used to Describe the Different Classes of Bacterial Antigens Is Explained in the Preface. However, the Authors Would like to Mention That a Different Convention Is Used in Some Laboratories, for Example O:L, K:L, H:7 Is Equivalent to O1:K1:H7. In Methods in Microbiology; Bergan, T., Ed.; Academic Press: Cambridge, MA, USA, 1984; Volume 14, pp. 43–112. [Google Scholar]
- Larsen, A.G.; Vogensen, F.K.; Josephsen, J. Antimicrobial Activity of Lactic Acid Bacteria Isolated from Sour Doughs: Purification and Characterization of Bavaricin A, a Bacteriocin Produced by Lactobacillus Bavaricus MI401. J. Appl. Bacteriol. 1993, 75, 113–122. [Google Scholar] [CrossRef]
- Malberg Tetzschner, A.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In Silico Genotyping of E. coli Isolates for Extraintestinal Virulence Genes by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2020, 58, e01269-20. [Google Scholar] [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A User-Friendly Web Server to Thoroughly Mine RiPPs and Bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A Viable Alternative to Antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Mushtaq, H.; Bakht, J.; Bacha, N. Screening of the Novel Colicinogenic Gram-Negative Rods against Pathogenic E. coli O157:H7. Indian J. Med. Microbiol. 2015, 33, S67–S72. [Google Scholar] [CrossRef]
- Pérez-Berezo, T.; Pujo, J.; Martin, P.; Le Faouder, P.; Galano, J.-M.; Guy, A.; Knauf, C.; Tabet, J.C.; Tronnet, S.; Barreau, F.; et al. Identification of an Analgesic Lipopeptide Produced by the Probiotic E. coli Strain Nissle 1917. Nat. Commun. 2017, 8, 1314. [Google Scholar] [CrossRef] [PubMed]
- Sonnenborn, U.; Schulze, J. The Non-Pathogenic E. coli Strain Nissle 1917—Features of a Versatile Probiotic. Microb. Ecol. Health Dis. 2009, 21, 122–158. [Google Scholar] [CrossRef]
- Micenková, L.; Bosák, J.; Smatana, S.; Novotný, A.; Budinská, E.; Šmajs, D. Administration of the Probiotic E. coli Strain A0 34/86 Resulted in a Stable Colonization of the Human Intestine During the First Year of Life. Probiotics Antimicrob. Proteins 2020, 12, 343–350. [Google Scholar] [CrossRef]
- Jordi, B.J.A.M.; Boutaga, K.; van Heeswijk, C.M.E.; van Knapen, F.; Lipman, L.J.A. Sensitivity of Shiga Toxin-Producing E. coli (STEC) Strains for Colicins under Different Experimental Conditions. FEMS Microbiol. Lett. 2001, 204, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Chérier, D.; Patin, D.; Blanot, D.; Touzé, T.; Barreteau, H. The Biology of Colicin M and Its Orthologs. Antibiotics 2021, 10, 1109. [Google Scholar] [CrossRef]
- Nougayrède, J.P.; Chagneau, C.V.; Motta, J.P.; Bossuet-Greif, N.; Belloy, M.; Taieb, F.; Gratadoux, J.J.; Thomas, M.; Langella, P.; Oswald, E. A Toxic Friend: Genotoxic and Mutagenic Activity of the Probiotic Strain E. coli Nissle 1917|MSphere. Available online: https://journals.asm.org/doi/full/10.1128/mSphere.00624-21 (accessed on 22 December 2022).
- Pleguezuelos-Manzano, C.; Puschhof, J.; Rosendahl Huber, A.; van Hoeck, A.; Wood, H.M.; Nomburg, J.; Gurjao, C.; Manders, F.; Dalmasso, G.; Stege, P.B.; et al. Mutational Signature in Colorectal Cancer Caused by Genotoxic Pks+ E. coli. Nature 2020, 580, 269–273. [Google Scholar] [CrossRef]
- Azam, M.W.; Khan, A.U. CRISPRi-Mediated Suppression of E. coli Nissle 1917 Virulence Factors: A Strategy for Creating an Engineered Probiotic Using CsgD Gene Suppression. Front. Nutr. 2022, 9, 938989. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Weiss, A.A. Probiotic Properties of E. coli Nissle in Human Intestinal Organoids. mBio 2020, 11, e01470-20. [Google Scholar] [CrossRef]
- Tu, M.; Cheng, S.; Lu, W.; Du, M. Advancement and Prospects of Bioinformatics Analysis for Studying Bioactive Peptides from Food-Derived Protein: Sequence, Structure, and Functions. TrAC Trends Anal. Chem. 2018, 105, 7–17. [Google Scholar] [CrossRef]
- Hammami, R.; Zouhir, A.; Le Lay, C.; Ben Hamida, J.; Fliss, I. BACTIBASE Second Release: A Database and Tool Platform for Bacteriocin Characterization. BMC Microbiol. 2010, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Cameron, A.; Zaheer, R.; Adator, E.H.; Barbieri, R.; Reuter, T.; McAllister, T.A. Bacteriocin Occurrence and Activity in E. coli Isolated from Bovines and Wastewater. Toxins 2019, 11, 475. [Google Scholar] [CrossRef] [PubMed]
- Sabino, Y.N.V.; de Araújo, K.C.; Assis, F.G.d.V.d.; Moreira, S.M.; Lopes, T.d.S.; Mendes, T.A.d.O.; Huws, S.A.; Mantovani, H.C. In Silico Screening Unveil the Great Potential of Ruminal Bacteria Synthesizing Lasso Peptides. Front. Microbiol. 2020, 11, 576738. [Google Scholar] [CrossRef]
- Poptsova, M.S.; Gogarten, J.P. Using Comparative Genome Analysis to Identify Problems in Annotated Microbial Genomes. Microbiology 2010, 156, 1909–1917. [Google Scholar] [CrossRef]
- Kassaa, I.A.; Rafei, R.; Moukhtar, M.; Zaylaa, M.; Gharsallaoui, A.; Asehraou, A.; Omari, K.E.; Shahin, A.; Hamze, M.; Chihib, N.-E. LABiocin Database: A New Database Designed Specifically for Lactic Acid Bacteria Bacteriocins. Int. J. Antimicrob. Agents 2019, 54, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Micenková, L.; Bosák, J.; Kucera, J.; Hrala, M.; Dolejšová, T.; Šedo, O.; Linke, D.; Fišer, R.; Šmajs, D. Colicin Z, a Structurally and Functionally Novel Colicin Type That Selectively Kills Enteroinvasive E. coli and Shigella Strains. Sci. Rep. 2019, 9, 11127. [Google Scholar] [CrossRef] [PubMed]
- Marković, K.G.; Grujović, M.Ž.; Koraćević, M.G.; Nikodijević, D.D.; Milutinović, M.G.; Semedo-Lemsaddek, T.; Djilas, M.D. Colicins and Microcins Produced by Enterobacteriaceae: Characterization, Mode of Action, and Putative Applications. Int. J. Environ. Res. Public Health 2022, 19, 11825. [Google Scholar] [CrossRef] [PubMed]
- Tahamtan, Y.; Golestan, F.; Moazamian, E. Evaluation of Colicin Effect on the Induction of Treated Mice in Prevention of Infection Caused by E. coli K99. Int. J. Enteric Pathog. 2016, 4, 28–32. [Google Scholar] [CrossRef][Green Version]
- Gillor, O.; Kirkup, B.C.; Riley, M.A. Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2004; ISBN 978-0-08-049020-5. [Google Scholar]
- Jin, X.; An, S.; Kightlinger, W.; Zhou, J.; Hong, S.H. Engineering E. coli to Produce and Secrete Colicins for Rapid and Selective Biofilm Cell Killing. AIChE J. 2021, 67, e17466. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | Code | Serotype | Virulence Genes | Origin | n | Code | Serotype | Virulence Genes | Origin |
|---|---|---|---|---|---|---|---|---|---|
| 1 | SHU-26 | O157:H7 | stx2 eae | HUS | 41 | BD 99 | O157:H7 | stx2, eae, hlyA | HC |
| 2 | E65-01 | O157:H7 | stx2 eae | HUS | 42 | BD 104 | O157:H7 | stx2, eae, hlyA | HC |
| 3 | E89-01 | O157:H7 | stx2 eae | HUS | 43 | BD 105 | O157:H7 | stx2, eae, hlyA | HC |
| 4 | E104-01 | O157:H7 | stx2 eae | HUS | 44 | BD 111 | O157:H7 | stx2, eae, hlyA | HC |
| 5 | E109-01 | O157:H7 | stx2 eae | HUS | 45 | 9 | O157:H7 | stx2, eae, hlyA | HUS |
| 6 | E110-01 | O157:H7 | stx2 eae | HUS | 46 | HUS 58 | O157:H7 | stx2, eae, hlyA | HUS |
| 7 | E111-01 | O157:H7 | stx2 eae | HUS | 47 | 72 | O157:H7 | stx2, eae, hlyA | HUS |
| 8 | E112-01 | O157:H7 | stx2 eae | WD | 48 | HUS 73 | O157:H7 | stx2, eae, hlyA | HUS |
| 9 | E113-01 | O157:H7 | stx2 eae | WD | 49 | 76 | O157:H7 | stx2, eae, hlyA | HUS |
| 10 | E114-01 | O157:H7 | stx2 eae | WD | 50 | 77 | O157:H7 | stx2, eae, hlyA | HUS |
| 11 | E115-01 | O157:H7 | stx2 eae | WD | 51 | INE 84-3 | O157:H7 | stx2, eae, hlyA | HUS |
| 12 | E116-01 | O157:H7 | stx2 eae | WD | 52 | 123 | O157:H7 | stx2, eae, hlyA | HUS |
| 13 | E109-02 | O157:H7 | stx2 eae | WD | 53 | HUS 125 | O157:H7 | stx2, eae, hlyA | HUS |
| 14 | E92-02 | O157:H7 | stx2 eae | HUS | 54 | HUS 126 | O157:H7 | stx2, eae, hlyA | HUS |
| 15 | E44-02 | O157:H7 | stx2 eae | HUS | 55 | HUS 127 | O157:H7 | stx2, eae, hlyA | HUS |
| 16 | E42-02 | O157:H7 | stx2 eae | WD | 56 | 144 | O157:H7 | stx2, eae, hlyA | HUS |
| 17 | E123-01 | O157:H7 | stx2 eae | HC | 57 | 147 | O157:H7 | stx2, eae, hlyA | HUS |
| 18 | E117-01 | O157:H7 | stx2 eae | WD | 58 | 148 | O157:H7 | stx2, eae, hlyA | HUS |
| 19 | E118-01 | O157:H7 | stx2 eae | WD | 59 | HUS 156 | O157:H7 | stx2, eae, hlyA | HUS |
| 20 | E309-02 | O157:H7 | stx2 eae | HUS | 60 | HUS 161 | O157:H7 | stx2, eae, hlyA | HUS |
| 21 | E192-02 | O157:H7 | stx2 eae | WD | 61 | HUS 179 | O157:H7 | stx2, eae, hlyA | HUS |
| 22 | E165-02 | O157:H7 | stx2 eae | WD | 62 | HUS 196 | O157:H7 | vt1, stx2, eae, hlyA | HUS |
| 23 | E161-02 | O157:H7 | stx2 eae | WD | 63 | INE 198-4 | O157:H7 | stx2, eae, hlyA | HUS |
| 24 | E121-02 | O157:H7 | stx2 eae | WD | 64 | 226 | O157:H7 | stx2, eae, hlyA | HUS |
| 25 | E111-02 | O157:H7 | stx2 eae | WD | 65 | 230 | O157:H7 | stx2, eae, hlyA | HUS |
| 26 | E110-02 | O157:H7 | stx2 eae | WD | 66 | 238 | O157:H7 | stx2, eae, hlyA | HUS |
| 27 | E310-03 | O157:H7 | stx2 eae | WD | 67 | HUS 239 | O157:H7 | stx2, eae, hlyA | HUS |
| 28 | E211-03 | O157:H7 | stx2 eae | HC | 68 | 272 | O157:H7 | stx2, eae, hlyA | HUS |
| 29 | E205-03 | O157:H7 | stx2 eae | HC | 69 | 291 | O157:H7 | stx2, eae, hlyA | HUS |
| 30 | E121-03 | O157:H7 | stx2 eae | HC | 70 | HUS 303 | O157:H7 | stx2, eae, hlyA | HUS |
| 31 | E111-03 | O157:H7 | stx2 eae | HC | 71 | 307 | O157:H7 | stx2, eae, hlyA | HUS |
| 32 | E92-03 | O157:H7 | stx2 eae | HUS | 72 | HUS 312 | O157:H7 | stx2, eae, hlyA | HUS |
| 33 | E16-03 | O157:H7 | stx2 eae | HC | 73 | HUS 316 | O157:H7 | stx2, eae, hlyA | HUS |
| 34 | E14-03 | O157:H7 | stx2 eae | HC | 74 | HUS 331 | O157:H7 | stx2, eae, hlyA | HUS |
| 35 | GB | O157:H7 | stx2 eae | HC | 75 | HUS 349 | O157:H7 | stx2, eae, hlyA | HUS |
| 36 | E30-00 | O157:H7 | stx2 eae | HUS | 76 | HUS 352 | O157:H7 | stx2, eae, hlyA | HUS |
| 37 | E3-99 | O157:H7 | stx2 eae | HUS | 77 | HUS 357 | O157:H7 | stx2, eae, hlyA | HUS |
| 38 | E93-03 | O157:H7 | stx2 eae | HC | 78 | 358 | O157:H7 | stx2, eae, hlyA | HUS |
| 39 | BD 8 | O157:H7 | stx2, eae, hlyA | HC | 79 | HUS 360 | O157:H7 | stx2, eae, hlyA | HUS |
| 40 | BD 14 | O157:H7 | stx2, eae, hlyA | HC | 80 | HUS 361-4 | O157:H7 | stx2, eae, hlyA | HUS |
| Colicin | Primer | Sequence | Amplicon Size (Bp) |
|---|---|---|---|
| B | colicinB-F | AAGAAAATGACGAGAAGACG | 493 |
| colicinB-R | GAAAGACCAAAGGCTATAAGG | ||
| E1 | colicinE1-F | TGTGGCATCGGGCGAGAATA | 650 |
| colicinE1-R | CTGCTTCCTGAAAAGCCTTTT | ||
| E7 | ColE7-F | GCATTCTGCCATCTGAAAT | 431 |
| ColE7-R | CTTCTGCCCACTTTCTTTCG | ||
| Ia | ColIa-F | GCATGCAAATGACGCTCTTA | 473 |
| ColIa-R | GAGGACGCCAGTTCTCTGTC | ||
| K | ColK-F | CAGAGGTCGCTGAACATGAA | 469 |
| ColK-R | TCCGCTAAATCCTGAGCAAT | ||
| M | ColM-F | GCTTACCACTTCGCAAAACC | 429 |
| ColM-R | GAGCGACTCTCCGATAATGC | ||
| S4 | ColS4-F | TATATGGCCCAACTGCTGGT | 456 |
| ColS4-R | CGTAAGGACGGACACCTGTT | ||
| B17 | microcin B17-F | TCACGCCAGTCTCCATTAGGTGTTGGCATT | 135 |
| microcin B17-R | TTCCGCCGCTGCCACCGTTTCCACCACTAC | ||
| H47 | microcin H47-F | CACTTTCATCCCTTCGGATTG | 227 |
| microcin H47-R | AGCTGAAGTCGCTGGCGCACCTCC | ||
| J25 | microcin J25-F | TCAGCCATAGAAAGATATAGGTGTACCAAT | 175 |
| microcin J25-R | TGATTAAGCATTTTCATTTTAATAAAGTGT | ||
| V | microcin V-F | CACACACAAAACGGGAGCTGTT | 680 |
| microcin V-R | CTTCCCGCAGCATAGTTCCAT |
| Colicigenic E. coli Strains | STEC Strains Inhibited | Colicin | Microcin | |
|---|---|---|---|---|
| ID Strain | Serogroup | % (Positive Reading) | ||
| 4.8 | ONT | 90.1 (73) | B | M, H47 |
| 8.2 | O77 | 90.1 (73) | Ia | C7 |
| 13.7 | O174 | 95.1 (77) | Ia, E7, B | C7, J25 |
| 24.7 | ONT | 95.1 (77) | B | M, H47 |
| 27.4 | O26 | 92.6 (75) | Ia, E7, B | C7, J25 |
| 27.12 | O141 | 92.6 (75) | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García, M.D.; Ruiz, M.J.; Medina, L.M.; Vidal, R.; Padola, N.L.; Etcheverria, A.I. Molecular and Genetic Characterization of Colicinogenic Escherichia coli Strains Active against Shiga Toxin-Producing Escherichia coli O157:H7. Foods 2023, 12, 2676. https://doi.org/10.3390/foods12142676
García MD, Ruiz MJ, Medina LM, Vidal R, Padola NL, Etcheverria AI. Molecular and Genetic Characterization of Colicinogenic Escherichia coli Strains Active against Shiga Toxin-Producing Escherichia coli O157:H7. Foods. 2023; 12(14):2676. https://doi.org/10.3390/foods12142676
Chicago/Turabian StyleGarcía, Mauro D., María J. Ruiz, Luis M. Medina, Roberto Vidal, Nora L. Padola, and Analía I. Etcheverria. 2023. "Molecular and Genetic Characterization of Colicinogenic Escherichia coli Strains Active against Shiga Toxin-Producing Escherichia coli O157:H7" Foods 12, no. 14: 2676. https://doi.org/10.3390/foods12142676
APA StyleGarcía, M. D., Ruiz, M. J., Medina, L. M., Vidal, R., Padola, N. L., & Etcheverria, A. I. (2023). Molecular and Genetic Characterization of Colicinogenic Escherichia coli Strains Active against Shiga Toxin-Producing Escherichia coli O157:H7. Foods, 12(14), 2676. https://doi.org/10.3390/foods12142676