
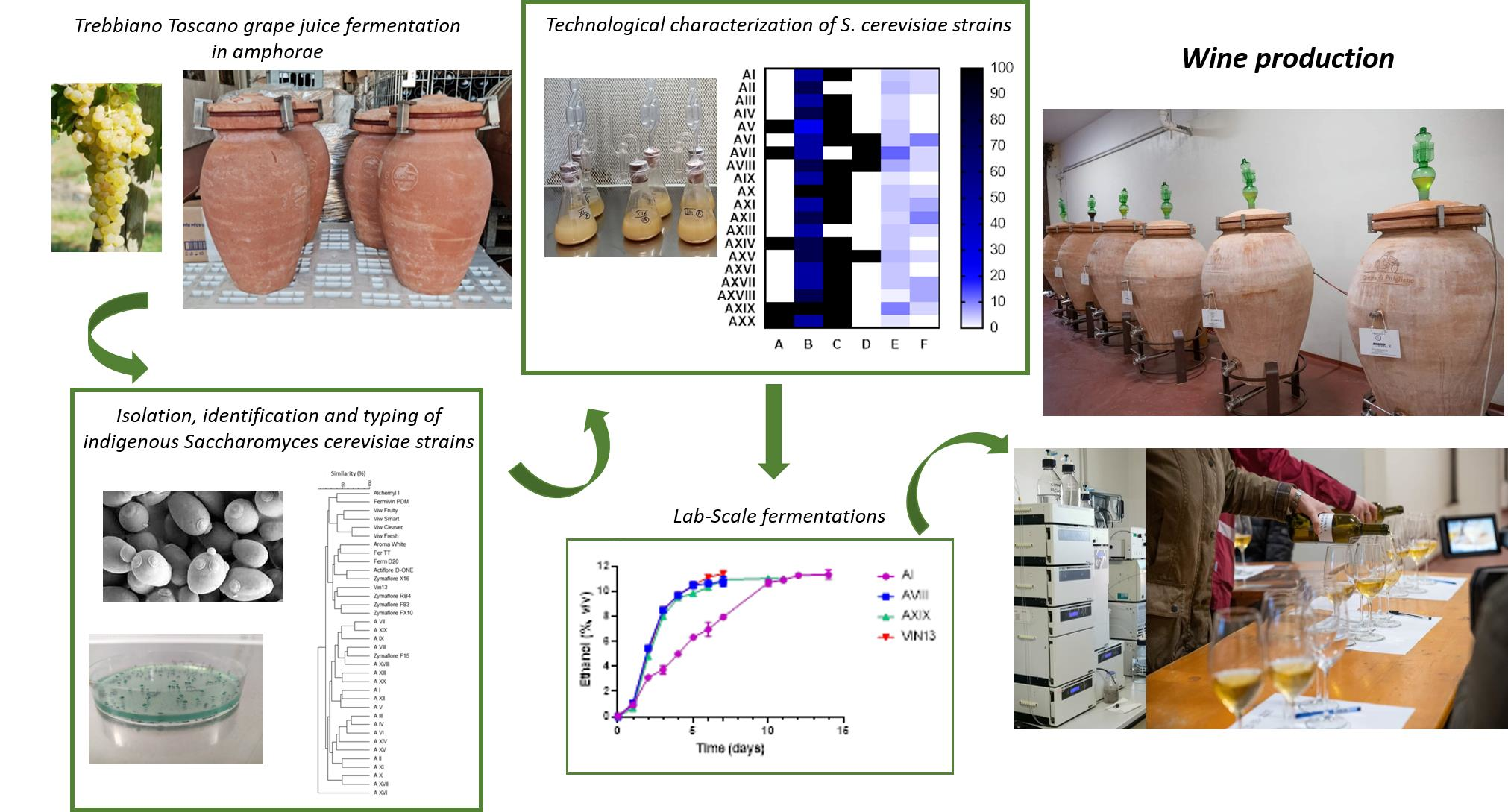
Management of in-Amphora “Trebbiano Toscano” Wine Production: Selection of Indigenous Saccharomyces cerevisiae Strains and Influence on the Phenolic and Sensory Profile

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cellar Fermentations to Isolate Indigenous S. cerevisiae Yeasts
2.2. Microbiological Analysis
2.3. Identification and Typing of Saccharomyces cerevisiae Strains
2.4. Technological Characterization of S. cerevisiae Strains
2.4.1. Killer Activity Tests
2.4.2. Hydrogen Sulfide Production
2.4.3. β-glucosidase Activity
2.4.4. Protease Activity
2.4.5. Fermentative Capacity
2.5. Lab-Scale Fermentations
2.6. Cellar Fermentations
2.7. Chemical Analysis
2.8. Phenolic Compounds Determination
2.9. Biogenic Amines Determination
2.10. Sensory ANALYSIS
2.11. Statistical Analysis
3. Results
3.1. Alcoholic Fermentation Kinetics in 300 L Amphorae
3.2. Chemical and Microbial Characteristics of the Wines during Ageing in 300 L Amphorae
3.3. Biodiversity of Saccharomyces cerevisiae during Alcoholic Fermentation in 300 L Amphorae
3.4. Selection of Indigenous S. cerevisiae Strains
3.5. Fermentation Performances of Selected Yeast Strains in 20L Amphorae Fermentations
3.6. Use of Indigenous S. cerevisiae Strains in Cellar Vinifications Carried out in 300 L Amphorae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Egaña-Juricic, M.E.; Gutiérrez-Gamboa, G.; Moreno-Simunovic, Y. Making wine in Pañul’s craft pottery vessels: A first approach in the study of the dynamic of alcoholic fermentation and wine volatile composition. Ciênc. Téc. Vitiviníc 2022, 37, 29–38. [Google Scholar] [CrossRef]
- Wine Producers Have Started Making Wine in Amphora. Available online: https://www.bkwine.com/features/winemaking-viticulture/making-wine-inamphora/ (accessed on 10 February 2023).
- Gil i Cortiella, M.G.; Úbeda, C.; Covarrubias, J.I.; Peña-Neira, Á. Chemical, physical, and sensory attributes of Sauvignon blanc wine fermented in different kinds of vessels. Innov. Food Sci. Emerg. Technol. 2020, 66, 102521. [Google Scholar] [CrossRef]
- Gil i Cortiella, M.; Ubeda, C.; Covarrubias, J.I.; Laurie, V.F.; Peña-Neira, Á. Chemical and physical implications of the use of alternative vessels to oak barrels during the production of white wines. Molecules 2021, 26, 554. [Google Scholar] [CrossRef] [PubMed]
- Baiano, A.; Varva, G. Evolution of physico-chemical and sensory characteristics of Minutolo white wines during aging in amphorae: A comparison with stainless steel tanks. LWT Food Sci. Technol. 2019, 103, 78–87. [Google Scholar] [CrossRef]
- Baiano, A.; Mentana, A.; Quinto, M.; Centonze, D.; Longobardi, F.; Ventrella, A.; Agostiano, A.; Varva, G.; De Gianni, A.; Terracone, C.; et al. The effect of in-amphorae aging on oenological parameters, phenolic profile and volatile composition of Minutolo white wine. Food Res. Int. 2015, 74, 294–305. [Google Scholar] [CrossRef]
- Cabrita, M.J.; Martins, N.; Barrulas, P.; Garcia, R.; Dias, C.B.; Pérez-Álvarez, E.P.; Costa Freitas, A.M.; Garde-Cerdán, T. Multi-element composition of red, white and palhete amphora wines from Alentejo by ICPMS. Food Control 2018, 92, 80–85. [Google Scholar] [CrossRef]
- Martins, N.; Garcia, R.; Mendes, D.; Freitas, A.M.C.; da Silva, M.G.; Cabrita, M.J. An ancient winemaking technology: Exploring the volatile composition of amphora wines. LWT Food Sci. Technol. 2018, 96, 288–295. [Google Scholar] [CrossRef]
- Capece, A.; Siesto, G.; Poeta, C.; Pietrafesa, R.; Romano, P. Indigenous yeast population from Georgian aged wines produced by traditional “Kakhetian” method. Food Microb. 2013, 36, 447–455. [Google Scholar] [CrossRef]
- Seo, G.H.; Yun, J.H.; Chung, S.K.; Park, W.P.; Lee, D.S. Physical properties of Korean earthenware containers affected by soy sauce fermentation use. Food Sci. Biotechnol. 2006, 15, 168–172. [Google Scholar]
- Rossetti, F.; Boselli, E. Effects of in-amphorae winemaking on the chemical and sensory profile of Chardonnay wine. Sci. Agric. Bohemic 2017, 48, 39–46. [Google Scholar] [CrossRef]
- Day, M.P.; Schmidt, S.A.; Smith, P.A.; Wilkes, E.N. Use and impact of oxygen during winemaking. Aust. J. Grape Wine Res. 2015, 21, 693–704. [Google Scholar] [CrossRef]
- Pozzatti, M.; Guerra, C.C.; Martins, G.; dos Santos, I.D.; Wagner, R.; Ferrão, M.F.; Manfroi, V. Effects of winemaking on’marselan’red wines: Volatile compounds and sensory aspects. Ciênc. Téc. Vitiviníc 2020, 35, 63–75. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A.; Sroka, P.; Siuta, M. The impact of oxygen at various stages of vinification on the chemical composition and the antioxidant and sensory properties of white and red wines. Int. J. Food Sci. 2020, 2020, 7902974. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma-a review. S. Afr. J. Enol. Vitic. 2020, 21, 97–129. [Google Scholar] [CrossRef]
- Vilanova, M.; Sieiro, C. Contribution by Saccharomyces cerevisiae yeast to fermentative flavour compounds in wines from cv. Albariño. J. Ind. Microbiol. Biotechnol. 2006, 33, 929–933. [Google Scholar] [CrossRef] [PubMed]
- King, E.S.; Swiegers, J.H.; Travis, B.; Francis, I.L.; Bastian, S.E.; Pretorius, I.S. Coinoculated fermentations using Saccharomyces yeasts affect the volatile composition and sensory properties of Vitis vinifera L. cv. Sauvignon blanc wines. J. Agric. Food Chem. 2008, 56, 10829–10837. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Francesca, N.; Chiurazzi, M.; Romano, R.; Aponte, M.; Settanni, L.; Moschetti, G. Indigenous yeast communities in the environment of “Rovello bianco” grape variety and their use in commercial white wine fermentation. World J. Microbiol. Biotechnol. 2010, 26, 337–351. [Google Scholar] [CrossRef]
- Ganucci, D.; Guerrini, S.; Mangani, S.; Vincenzini, M.; Granchi, L. Quantifying the effects of ethanol and temperature on the fitness advantage of predominant Saccharomyces cerevisiae strains occurring in spontaneous wine fermentations. Front. Microbiol. 2018, 9, 1563. [Google Scholar] [CrossRef]
- Granchi, L.; Ganucci, D.; Buscioni, G.; Mangani, S.; Guerrini, S. The biodiversity of Saccharomyces cerevisiae in spontaneous wine fermentation: The occurrence and persistence of winery-strains. Fermentation 2019, 5, 86. [Google Scholar] [CrossRef]
- Guerrini, S.; Barbato, D.; Guerrini, L.; Mari, E.; Buscioni, G.; Mangani, S.; Romboli, R.; Galli, V.; Parenti, A.; Granchi, L. Selection of indigenous Saccharomyces cerevisiae strains and exploitation of a pilot-plant to produce fresh yeast starter cultures in a winery. Fermentation 2021, 7, 99. [Google Scholar] [CrossRef]
- Spagna, G.; Barbagallo, R.N.; Pifferi, P.G. Fining treatments of white wines by means of polymeric adjuvants for their stabilization against browning. J. Agric. Food Chem. 2000, 48, 4619–4627. [Google Scholar] [CrossRef]
- Mateo, J.J.; Jimenez, M. Monoterpenes in grape juice and wines: Review. J. Chromatography A 2000, 881, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Granchi, L.; Bosco, M.; Messini, A.; Vincenzini, M. Rapid detection and quantification of yeast species during spontaneous wine fermentation by PCR–RFLP analysis of the rDNA ITS region. J. Appl. Microbiol. 1999, 87, 949–956. [Google Scholar] [CrossRef]
- Legras, J.L.; Karst, F. Optimisation of interdelta analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef]
- Shannon, S.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Philliskirk, G.; Young, T.W. The occurrence of killer character in yeasts of various genera. Antonie Van Leeuwenhoek 1975, 41, 147–151. [Google Scholar] [CrossRef]
- Aponte, M.; Romano, R.; Villano, C.; Blaiotta, G. Dominance of S. cerevisiae Commercial Starter Strains during Greco di Tufo and Aglianico Wine Fermentations and Evaluation of Oenological Performances of Some Indigenous/Residential Strains. Foods 2020, 9, 1549. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, L.F.; Espinosa, J.C.; Fernandez-Gonzalez, M.; Briones, A. ß-Glucosidase activity in a Saccharomyces cerevisiae wine strain. Int. J. Food Microbiol. 2003, 80, 171–176. [Google Scholar] [CrossRef]
- Flores, M.G.; Rodríguez, M.E.; Oteiza, J.M.; Barbagelata, R.J.; Lopes, C.A. Physiological characterization of Saccharomyces uvarum and Saccharomyces eubayanus from Patagonia and their potential for cidermaking. Int. J. Food Microbiol. 2017, 249, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, S.; Galli, V.; Barbato, D.; Facchini, G.; Mangani, S.; Pierguidi, L.; Granchi, L. Effects of Saccharomyces cerevisiae and Starmerella bacillaris on the physicochemical and sensory characteristics of sparkling pear cider (Perry). Eur. Food Res. Technol. 2023, 249, 341–352. [Google Scholar] [CrossRef]
- Tuberoso, C.I.G.; Congiu, F.; Serreli, G.; Mameli, S. Determination of dansylated amino acids and biogenic amines in Cannonau and Vermentino wines by HPLC-FLD. Food Chem. 2015, 175, 29–35. [Google Scholar] [CrossRef]
- Anonymous. OIV Standard for International Wine and Spirituous Beverages of Vitivinicultural Origin Competitions Resolution; OIV/CONCOURS 332A/2009; International Organization of Vine and Wine General Assembly (OIV): Zagreb, Croatia, 2009. [Google Scholar]
- Lange, J.N.; Faasse, E.; Tantikachornkiat, M.; Gustafsson, F.S.; Halvorsen, L.C.; Kluftinger, A.; Ledderhof, D.; Durall, D.M. Implantation and persistence of yeast inoculum in Pinot noir fermentations at three Canadian wineries. Int. J. Food Microbiol. 2014, 180, 56–61. [Google Scholar] [CrossRef]
- Delteil, D. Aspectos prácticos del levadurado en condiciones mediterráneas. Técnica de inoculación y relación entre la población seleccionada y la población indígena. Rev. Fr. Oenologie 2004, 189, 1–13. [Google Scholar]
- Clavijo, A.; Calderon, I.L.; Paneque, P. Effect of the use of commercial Saccharomyces strains in a newly established winery in Rond (Malaga, Spain). Antonie Van Leeuwenhoek 2011, 99, 727–731. [Google Scholar] [CrossRef]
- Maqueda, M.; Nevado, F.P.; Regodón, J.A.; Zamora, E.; Álvarez, M.L.; Rebollo, J.E.; Ramírez, M. A low-cost procedure for the production of fresh autochthonous wine yeast. J. Ind. Microbiol. Biotechnol. 2011, 38, 459–469. [Google Scholar] [CrossRef]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Spano, G.; Capozzi, V.; Logrieco, A.; Mita, G.; Grieco, F. Biodiversity and safety aspects of yeast strains characterized from vineyards and spontaneous fermentations in the Apulia Region, Italy. Food Microbiol. 2013, 36, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Tristezza, M.; Fantastico, L.; Vetrano, C.; Bleve, G.; Corallo, D.; Grieco, F.; Mita, G. Molecular and Technological Characterization of Saccharomyces cerevisiae strains Isolated from Natural Fermentation of Susumaniello Grape Must in Apulia, Southern Italy. Int. J. Microbiol. 2014, 2014, 897428. [Google Scholar] [CrossRef] [PubMed]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Grieco, F.; Tufariello, M.; Quarta, A.; Mita, G.; Spano, G. Autochthonous fermentation starters for the industrial production of Negroamaro wines. J. Ind. Microbiol. Biotechnol. 2012, 39, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Garofalo, C.; Chiriatti, M.A.; Grieco, F.; Spano, G. Microbial terroir and food innovation: The case of yeast biodiversity in wine. Microbiol. Res. 2015, 181, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; El Khoury, M.; Lucas, P.; Bely, M.; Russo, P.; Spano, G.; Capozzi, V. Autochthonous starter cultures and indigenous grape variety for regional wine production. J. Appl. Microbiol. 2015, 118, 1395–1408. [Google Scholar] [CrossRef]
- Grieco, F.; Tristezza, M.; Vetrano, C.; Bleve, G.; Panico, E.; Mita, G.; Logrieco, A. Exploitation of autochthonous microorganism potential to enhance the quality of Apulian wines. Ann. Microbiol. 2011, 61, 67–73. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romaniello, R.; Condelli, N.; Romano, P. Selected Indigenous Saccharomyces cerevisiae Strains as Profitable Strategy to Preserve Typical Traits of Primitivo Wine. Fermentation 2019, 5, 87. [Google Scholar] [CrossRef]
- Tesfaye, W.; Morales, M.L.; García-Parrilla, M.C.; Troncoso, A.M. Evolution of phenolic compounds during an experimental aging in wood of Sherry vinegar. J. Agric. Food Chem. 2002, 50, 7043–7061. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Heras, M.; González-Sanjosé, M.L.; González-Huerta, C. Consideration of the influence of aging process, type of wine and oenological classic parameters on the levels of wood volatile compounds present in red wines. Food Chem. 2007, 103, 1434–1448. [Google Scholar] [CrossRef]
- Suárez, R.; Suárez-Lepe, J.A.; Morata, A.; Calderón, F. The production of ethylphenols in wine by yeasts of the genera Brettanomyces and Dekkera. A review. Food Chem. 2007, 102, 10–21. [Google Scholar] [CrossRef]
- Oro, L.; Canonico, L.; Marinelli, V.; Ciani, M.; Comitini, F. Occurrence of Brettanomyces bruxellensis on Grape Berries and in Related Winemaking Cellar. Front. Microbiol. 2019, 10, 415. [Google Scholar] [CrossRef]
- Vincenzini, M.; Guerrini, S.; Mangani, S.; Granchi, L. Amino Acid Metabolisms and Production of Biogenic Amines and Ethyl Carbamate. In Biology of Microorganisms on Grapes, in Must and Wine; Part III; Helmut, K., Gottfried, U.-d., Jurgen, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 167–180. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A1+2 (Anchor VIN13) | A3+4 (Aroma White Enartis) | A5+6 (Spontaneous) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Months | 4 | 6 | 12 | 4 | 6 | 12 | 4 | 6 | 12 |
| Glucose (g/L) | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 |
| Fructose (g/L) | 0.2 ± 0.0 a | 0.2 ± 0.0 a | 0.2 ± 0.0 a | 0.1 ± 0.0 a | 0.1 ± 0.0 a | 0.1 ± 0.0 a | 0.5 b | 0.5 b | 0.2 a |
| Ethanol (%, v/v) | 11.8 ± 0.3 a | 11.8 ± 0.1 a | 11.7 ± 0.1 a | 11.8 ± 0.0 a | 11.8 ± 0.0 a | 11.7 ± 0.1 a | 11.8 ± 0.0 a | 11.8 ± 0.1 a | 11.9 ± 0.0 a |
| Glycerol (g/L) | 5.8 ± 0.3 a | 5.8 ± 0.0 a | 5.8 ± 0.1 a | 5.7 ± 0.3 a | 5.7 ± 0.1 a | 5.8 ± 0.0 a | 5.8 ± 0.1 a | 5.8 ± 0.0 a | 5.8 ± 0.1 a |
| Acetic acid (g/L) | 0.23 ± 0.00 a | 0.23 ± 0.00 a | 0.30 ± 0.01 b | 0.21 ± 0.00 a | 0.21 ± 0.01 a | 0.25 ± 0.01 a | 0.18 ± 0.01 a | 0.18 ± 0.00 a | 0.20 ± 0.01 a |
| Lactic acid (g/L) | 1.87 ± 0.03 a | 1.87 ± 0.04 a | 1.87 ± 0.07 a | 1.83 ± 0.04 a | 1.83 ± 0.06 a | 1.83 ± 0.00 a | 1.95 ± 0.00 a | 1.95 ± 0.04 a | 1.95 ± 0.04 a |
| S. cerevisiae (CFU/mL) | 5.60 × 103 a | 6.40 × 102 b | <10 | 2.40 × 102 a | 30 b | <10 | 3.23 × 104 a | 4.9 × 102 b | <10 |
| O. oeni (CFU/mL) | 7.60 × 103 a | 1.66 × 102 b | <10 | 3.00 × 103 a | 1.52 × 102 b | <10 | 6.56 × 104 a | 1.18 × 104 b | <10 |
| B. bruxellensis (CFU/mL) | 1.60 × 103 | <10 | <10 | 1.70 × 102 a | 6.10 × 102 b | <10 | 3.00 × 103 | <10 | <10 |
| % | Amphora 1 | Amphora 2 | Amphora 3 | Amphora 4 | Amphora 5 | Amphora 6 |
|---|---|---|---|---|---|---|
| Indigenous S. cerevisiae strains | ||||||
| AI | - | - | 5.6 | 7.6 | 6.7 | 2.2 |
| AII | - | - | 6.4 | 6.8 | 6.7 | 4.4 |
| AIII | - | - | - | - | 4.4 | 2.2 |
| AIV | - | - | - | - | 4.4 | 2.2 |
| AV | - | - | - | - | 6.7 | 2.2 |
| AVI | 7.5 | 5.7 | 14.0 | 13.0 | 2.2 | 6.7 |
| AVII | - | - | 7.1 | 6.1 | 13.3 | 11.1 |
| AVIII | - | - | 6.7 | 6.5 | 4.4 | 8.9 |
| AIX | - | - | - | - | 4.4 | - |
| AX | 7.6 | 5.6 | - | - | 4.4 | 4.4 |
| AXI | 12.5 | 14.5 | - | - | 4.4 | 4.4 |
| AXII | - | - | 20.1 | 19.9 | 2.2 | 4.4 |
| AXIII | 7.7 | 5.5 | - | - | 2.2 | 6.7 |
| AXIV | - | - | - | - | 2.2 | 6.7 |
| AXV | 7.1 | 6.1 | - | - | 4.4 | 6.7 |
| AXVI | - | - | - | - | 6.7 | 2.2 |
| AXVII | - | - | 12.5 | 14.5 | 4.4 | 4.4 |
| AXVIII | 13.0 | 14.0 | - | - | 2.2 | - |
| AXIX | 5.6 | 7.6 | - | - | 11.1 | 8.9 |
| AXX | - | - | - | - | 2.2 | 2.2 |
| Commercial strains | ||||||
| Anchor VIN13 | 25.6 | 27.6 | 6.5 | 6.9 | - | 8.9 |
| Aroma White Enartis | 5.7 | 7.7 | 14.3 | 12.3 | - | - |
| Zymaflore X16 | 7.7 | 5.7 | 6.8 | 6.4 | - | - |
| Enartis Ferm TT | - | - | 5.6 | 7.6 | - | - |
| Biodiversity indeces | ||||||
| H | 1.87 | 1.91 | 1.82 | 1.86 | 2.85 | 2.83 |
| e | 0.85 | 0.87 | 0.76 | 0.78 | 0.75 | 0.76 |
| Strain | AI | AII | AVII | AVIII | AXV | AXIX | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Glucose (g/L) | <0.1 | - | <0.1 | - | <0.1 | - | <0.1 | - | <0.1 | - | <0.1 | - |
| Fructose (g/L) | <0.1 | - | <0.1 | - | <0.1 | - | <0.1 | - | <0.1 | - | <0.1 | - |
| Ethanol (%, v/v) | 11.1 | 0.1 | 11.2 | 0.0 | 11.2 | 0.0 | 11.3 | 0.0 | 11.2 | 0.1 | 11.2 | 0.1 |
| Glycerol (g/L) | 5.8 a | 0.1 | 6.0 a | 0.1 | 5.8 a | 0.1 | 6.1 a | 0.0 | 5.4 b | 0.1 | 5.5 b | 0.1 |
| meso 2,3-butanediol (g/L) | 0.07 ab | 0.01 | 0.09 a | 0.01 | 0.07 ab | 0.01 | 0.02 b | 0.01 | 0.03 ab | 0.03 | 0.01 b | 0.00 |
| raceme 2,3-butanediol (g/L) | 0.18 ab | 0.01 | 0.15 ab | 0.03 | 0.13 a | 0.03 | 0.14 a | 0.02 | 0.15 ab | 0.01 | 0.23 b | 0.03 |
| Succinic acid (g/L) | 1.09 a | 0.01 | 0.86 ab | 0.03 | 0.87 ab | 0.01 | 0.90 ab | 0.04 | 0.75 b | 0.11 | 0.90 ab | 0.16 |
| Acetic acid (g/L) | 0.21 | 0.04 | 0.26 | 0.01 | 0.24 | 0.01 | 0.20 | 0.02 | 0.26 | 0.01 | 0.18 | 0.01 |
| Lactic acid (g/L) | 0.20 a | 0.01 | 0.15 ab | 0.01 | 0.17 ab | 0.01 | 0.13 b | 0.03 | 0.16 ab | 0.01 | 0.20 a | 0.01 |
| Fermentation rate (μmax, h−1) | 0.100 a | 0.002 | 0.090 a | 0.0004 | 0.093 a | 0.002 | 0.114 b | 0.006 | 0.097 a | 0.008 | 0.100 a | 0.004 |
| AI | AVIII | AXIX | VIN13 | |||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Glucose (g/L) | 1.10 a | - | 0.10 b | 0.07 | <0.05 | - | 0.10 b | - |
| Fructose (g/L) | 0.8 ac | 0.1 | 0.3 ab | 0.1 | 0.1 b | 0.1 | 0.9 c | 0.1 |
| Ethanol (% v/v) | 11.3 | 0.4 | 10.8 | 0.4 | 11.0 | 0.1 | 11.4 | 0.0 |
| Glycerol (g/L) | 7.7 ab | 0.1 | 8.1 a | 0.1 | 7.4 b | 0.1 | 7.3 b | 0.1 |
| Lactic acid (g/L) | 1.23 a | 0.14 | 0.19 b | 0.01 | 1.50 a | 0.15 | 0.12 b | 0.03 |
| Acetic acid (g/L) | 0.31 a | 0.01 | 0.12 b | 0.02 | 0.30 a | 0.03 | 0.15 b | 0.03 |
| Malic acid | <0.05 | - | 1.94 | 0.05 | <0.05 | - | 1.97 | 0.08 |
| Fermentation rate (h−1) | 0.056 a | 0.004 | 0.190 b | 0.008 | 0.167 b | 0.010 | 0.174 b | 0.008 |
| S. cerevisiae (CFU/mL) | 5.43 × 106 a | 0.41 × 106 | 6.80 × 107 b | 0.65 × 107 | 8.40 × 104 c | 0.48 × 104 | 1.50 × 106 d | 0.32 × 106 |
| O. oeni (CFU/mL) | 4.05 × 107 a | 0.09 × 107 | 9.10 × 103 b | 0.15 × 103 | 8.00 × 107 c | 0.12 × 107 | 8.80 × 104 d | 0.60 × 104 |
| % | Amphora A (AVIII) | Amphora B (AI+AVIII) | Amphora C (VIN13) |
|---|---|---|---|
| AI | - | 25 | 7 |
| AVIII | 75 | 54 | 40 |
| VIN13 | - | - | 25 |
| AXI | 7 | - | - |
| AXIX | - | - | 7 |
| AXXII | 11 | 11 | - |
| AXXIII | - | 7 | - |
| AXXIV | - | - | 3.5 |
| AXXV | 7 | - | 7 |
| AXXVII | 3 | - | |
| AXXIX | - | - | 3.5 |
| AXXXI | - | - | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrini, S.; Barbato, D.; Mangani, S.; Ganucci, D.; Buscioni, G.; Galli, V.; Triossi, A.; Granchi, L. Management of in-Amphora “Trebbiano Toscano” Wine Production: Selection of Indigenous Saccharomyces cerevisiae Strains and Influence on the Phenolic and Sensory Profile. Foods 2023, 12, 2372. https://doi.org/10.3390/foods12122372
Guerrini S, Barbato D, Mangani S, Ganucci D, Buscioni G, Galli V, Triossi A, Granchi L. Management of in-Amphora “Trebbiano Toscano” Wine Production: Selection of Indigenous Saccharomyces cerevisiae Strains and Influence on the Phenolic and Sensory Profile. Foods. 2023; 12(12):2372. https://doi.org/10.3390/foods12122372
Chicago/Turabian StyleGuerrini, Simona, Damiano Barbato, Silvia Mangani, Donatella Ganucci, Giacomo Buscioni, Viola Galli, Andrea Triossi, and Lisa Granchi. 2023. "Management of in-Amphora “Trebbiano Toscano” Wine Production: Selection of Indigenous Saccharomyces cerevisiae Strains and Influence on the Phenolic and Sensory Profile" Foods 12, no. 12: 2372. https://doi.org/10.3390/foods12122372
APA StyleGuerrini, S., Barbato, D., Mangani, S., Ganucci, D., Buscioni, G., Galli, V., Triossi, A., & Granchi, L. (2023). Management of in-Amphora “Trebbiano Toscano” Wine Production: Selection of Indigenous Saccharomyces cerevisiae Strains and Influence on the Phenolic and Sensory Profile. Foods, 12(12), 2372. https://doi.org/10.3390/foods12122372