

Brassinin Abundant in Brassicaceae Suppresses Melanogenesis through Dual Mechanisms of Tyrosinase Inhibition



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Ferric Reducing Antioxidant Power (FRAP) Assay
2.3. Cell-Free Mushroom Tyrosinase Activity Assay
2.4. Cell Culture
2.5. Cell Viability Assay
2.6. Measurement of Melanin Content
2.7. Cellular Tyrosinase Activity Assay
2.8. Real-Time PCR
2.9. Immunofluorescence Staining
2.10. Docking Simulation
2.11. Statistical Analysis
3. Results
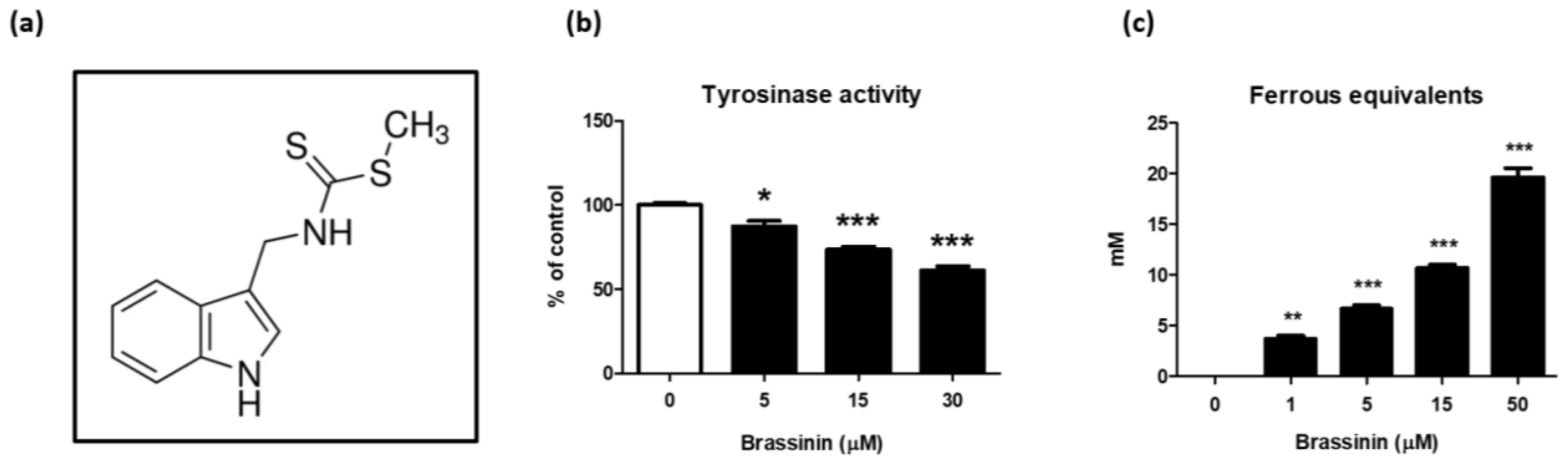
3.1. Brassinin Is an Antioxidant and Tyrosinase Inhibitor
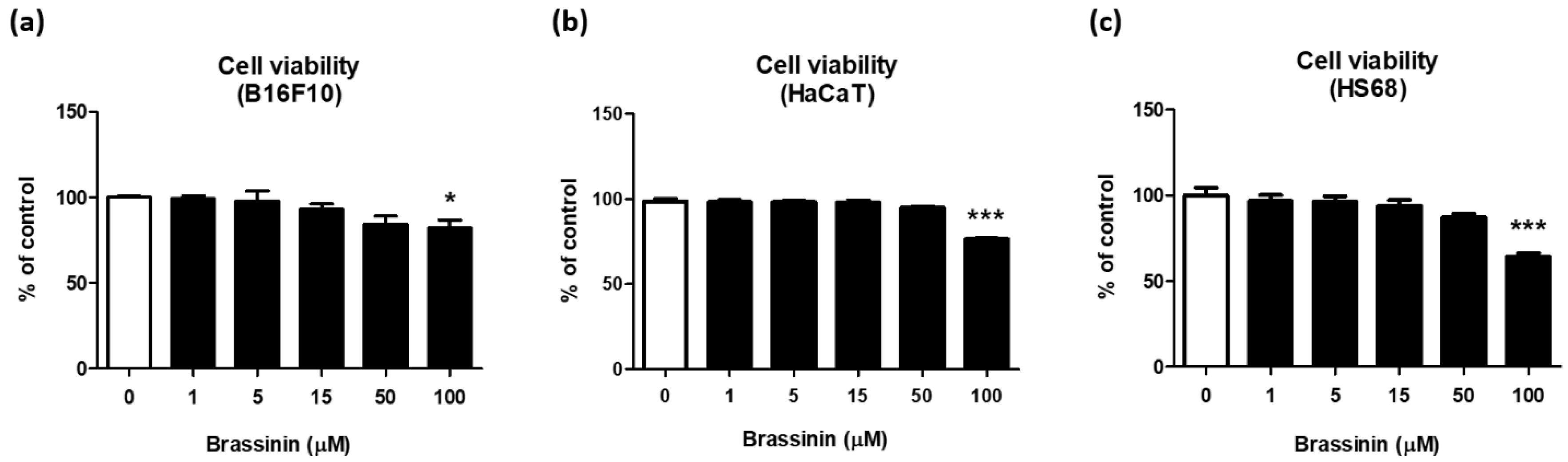
3.2. Brassinin Did Not Affect Cell Viability in the Range of 1 to 50 µM
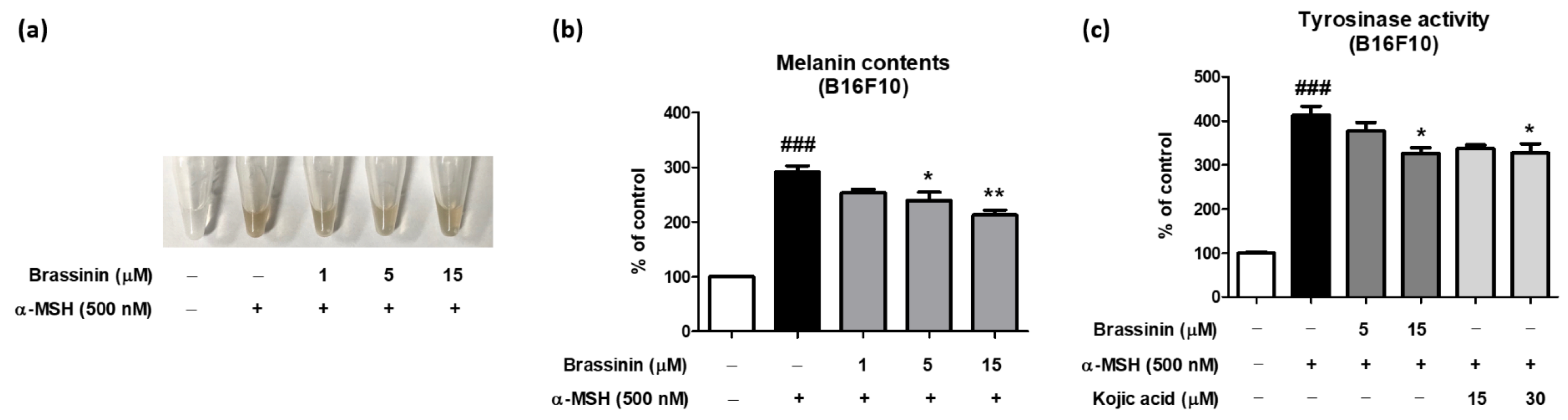
3.3. Brassinin Inhibits α-MSH-Induced Intracellular Melanin Accumulation and Tyrosinase Activity in B16F10 Cells
3.4. Brassinin Inhibits MITF Translocation into the Nucleus and Decreases the mRNA Level of Tyrosinase in α-MSH-Stimulated B16F10 Cells
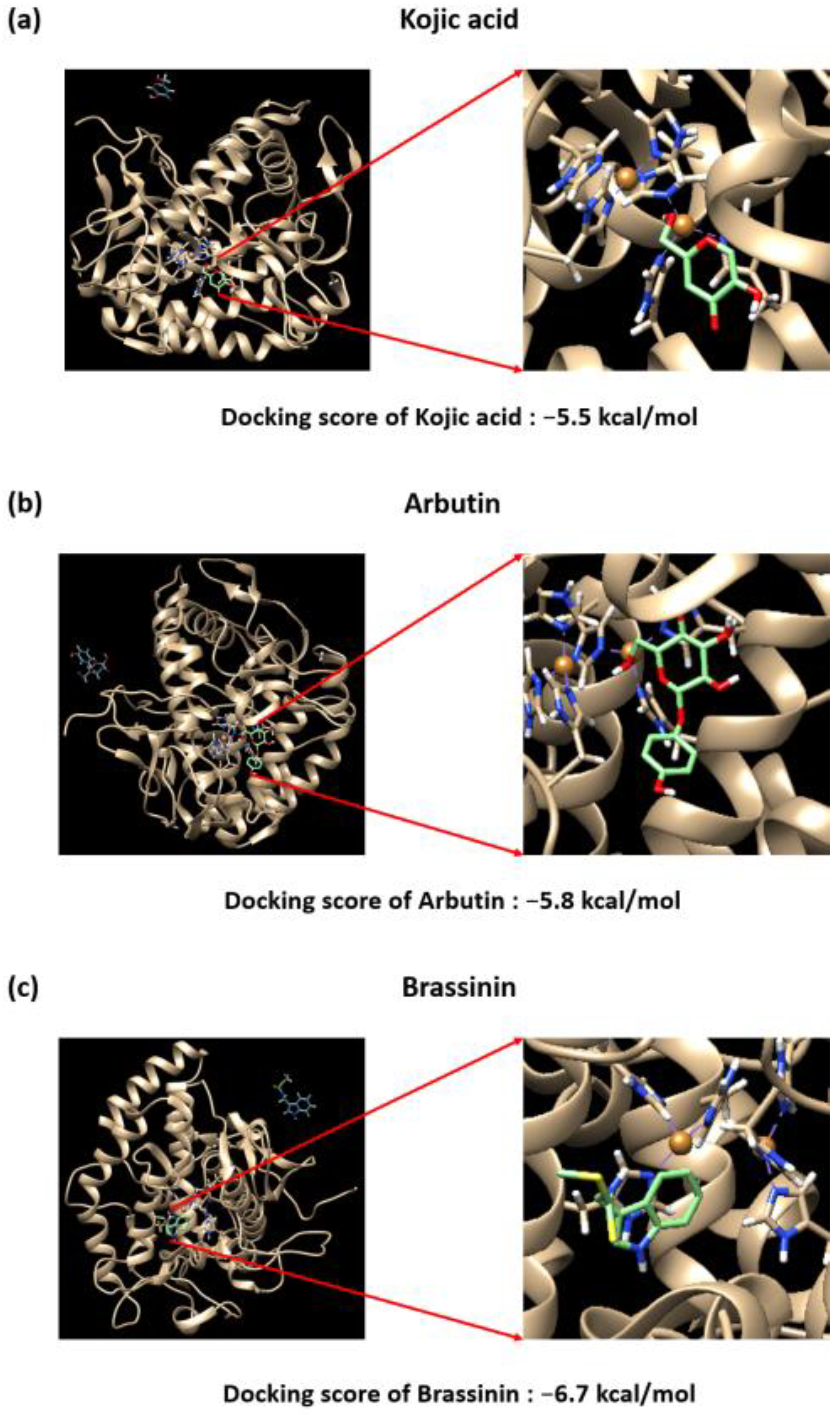
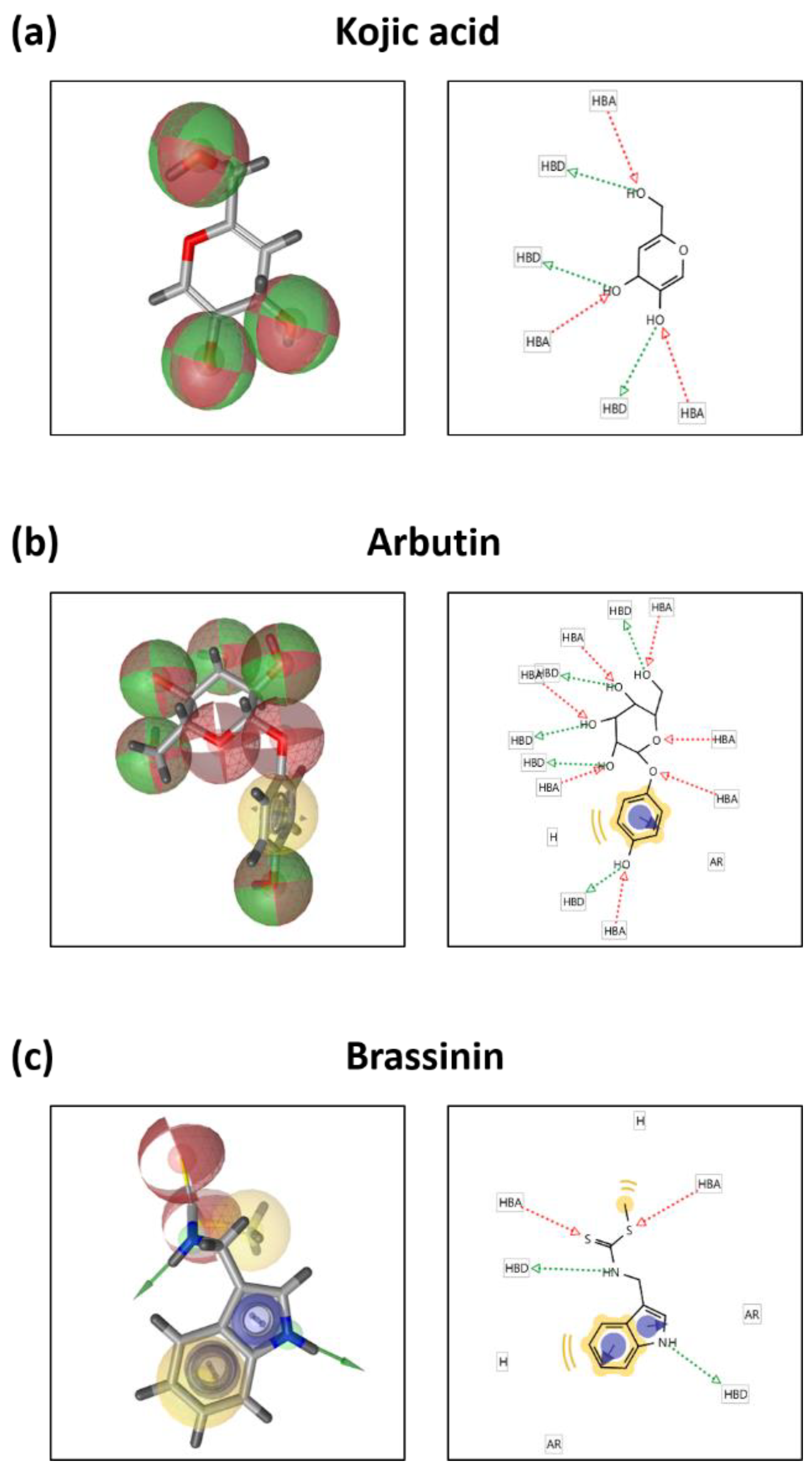
3.5. Brassinin May Bind to and Inactivate Tyrosinase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhou, Y.; Zeng, H.L.; Wen, X.Y.; Jiang, L.; Fu, C.H.; Hu, Y.B.; Lei, X.X.; Zhang, L.; Yu, X.; Yang, S.Y.; et al. Selaginellin Inhibits Melanogenesis via the MAPK Signaling Pathway. J. Nat. Prod. 2022, 85, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-H.; Jung, J.-G.; Kim, J.-E.; Bang, M.-A. Anti-melanogenic effects of Hordeum vulgare L. barely sprout extract in murine B16F10 melanoma cells. J. Nutr. Health 2019, 52, 168. [Google Scholar] [CrossRef]
- Beguerie, J.; Diego, C. Freckles, Lentigos, Melanocytic Nevi, and Melanoma. In Atlas of Dermatology, Dermatopathology and Venereology: Cutaneous Infectious and Neoplastic Conditions and Procedural Dermatology; Springer: Berlin/Heidelberg, Germany, 2022; pp. 403–433. [Google Scholar]
- Iriyama, S.; Ono, T.; Aoki, H.; Amano, S. Hyperpigmentation in solar lentigo is promoted by heparanase-induced loss of heparan sulfate chains at the dermal-epidermal junction. J. Dermatol. Sci. 2011, 64, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Konda, S.; Geria, A.; Halder, R. New Horizons in Treating Disorders of Hyperpigmentation in Skin of Color. Semin. Cutan. Med. Surg. 2012, 31, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Imen, M.B.; Chaabane, F.; Nadia, M.; Soumaya, K.J.; Kamel, G.; Leila, C.G. Anti-melanogenesis and antigenotoxic activities of eriodictyol in murine melanoma (B16-F10) and primary human keratinocyte cells. Life Sci. 2015, 135, 173–178. [Google Scholar] [CrossRef]
- Chen, Y.M.; Su, W.C.; Li, C.; Shi, Y.; Chen, Q.X.; Zheng, J.; Tang, D.L.; Chen, S.M.; Wang, Q. Anti-melanogenesis of novel kojic acid derivatives in B16F10 cells and zebrafish. Int. J. Biol. Macromol. 2019, 123, 723–731. [Google Scholar] [CrossRef]
- Maranduca, M.A.; Branisteanu, D.; Serban, D.N.; Branisteanu, D.C.; Stoleriu, G.; Manolache, N.; Serban, I.L. Synthesis and physiological implications of melanic pigments. Oncol. Lett. 2019, 17, 4183–4187. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Hearing, V.J. Physiological factors that regulate skin pigmentation. Biofactors 2009, 35, 193–199. [Google Scholar] [CrossRef]
- Son, H.-U.; Nam, D.-Y.; Kim, M.-A.; Cha, Y.-S.; Kim, J.-M.; Shin, Y.-K.; Lee, S.-H. Inhibitory Effect of Angelica keiskei Extracts on Melanogenesis. Korean J. Food Preserv. 2011, 18, 998–1001. [Google Scholar] [CrossRef]
- Hirata, N.; Naruto, S.; Ohguchi, K.; Akao, Y.; Nozawa, Y.; Iinuma, M.; Matsuda, H. Mechanism of the melanogenesis stimulation activity of (-)-cubebin in murine B16 melanoma cells. Bioorg. Med. Chem. 2007, 15, 4897–4902. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Chemistry of mixed melanogenesis--pivotal roles of dopaquinone. Photochem. Photobiol. 2008, 84, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Land, E.J.; Ramsden, C.A.; Riley, P.A. Quinone chemistry and melanogenesis. Methods Enzym. 2004, 378, 88–109. [Google Scholar] [CrossRef]
- Son, H.-U.; Lee, S.; Kim, M.-A.; Park, H.-J.; Lee, S.-H. Comparison of Melanogenesis-Inhibiting Activity by Extracts of Prunus persica Flower and Calyx. Korean J. Food Preserv. 2012, 19, 946–950. [Google Scholar] [CrossRef][Green Version]
- Lazou, K.; Sadick, N.; Kurfurst, R.; Bonnet, M.; Neveu, M.; Nizard, C.; Heusele, C.; Schnebert, S.; Perrier, E. The use of antisense strategy to modulate human melanogenesis. J. Drugs Dermatol. JDD 2007, 6, s2–s7. [Google Scholar] [PubMed]
- Chou, T.-H.; Ding, H.-Y.; Hung, W.; Liang, C.-H. Antioxidative characteristics and inhibition of alpha-melanocyte-stimulating hormone-stimulated melanogenesis of vanillin and vanillic acid from Origanum vulgare. Exp. Dermatol. 2010, 19, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Kim, H.-K.; Lee, K.; Jeon, H.; Cui, S.; Lee, Y.; Moon, B.; Kim, Y.-H.; Lee, Y.-S. Inhibitory effect of glyceollin isolated from soybean against melanogenesis in B16 melanoma cells. BMB Rep. 2010, 43, 461–467. [Google Scholar] [CrossRef]
- Lee, S.Y.; Baek, N.; Nam, T.G. Natural, semisynthetic and synthetic tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2016, 31, 1–13. [Google Scholar] [CrossRef]
- Draelos, Z.D. Skin lightening preparations and the hydroquinone controversy. Dermatol. Ther. 2007, 20, 308–313. [Google Scholar] [CrossRef]
- Serra-Baldrich, E.; Tribo, M.J.; Camarasa, J.G. Allergic contact dermatitis from kojic acid. Contact Dermat. 1998, 39, 86–87. [Google Scholar] [CrossRef]
- Ratnam, A.V.; Sastry, P.B.; Satyanarayana, B.V. Ascorbic acid and melanogenesis. Br. J. Derm. 1977, 97, 201–204. [Google Scholar] [CrossRef]
- Hu, Z.-M.; Zhou, Q.; Lei, T.-C.; Ding, S.-F.; Xu, S.-Z. Effects of hydroquinone and its glucoside derivatives on melanogenesis and antioxidation: Biosafety as skin whitening agents. J. Dermatol. Sci. 2009, 55, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.C.; Nycholat, C.M.; Montaut, S.; Xu, Y.; Khan, A.Q. Chemical defenses of crucifers: Elicitation and metabolism of phytoalexins and indole-3-acetonitrile in brown mustard and turnip. Phytochemistry 2002, 59, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.; Zheng, Q.-a.; Sarma-Mamillapalle, V. The Phytoalexins from Brassicaceae: Structure, Biological Activity, Synthesis and Biosynthesis. Nat. Prod. Commun. 2007, 2, 319–330. [Google Scholar] [CrossRef]
- Hong, T.; Ham, J.; Song, J.; Song, G.; Lim, W. Brassinin inhibits proliferation in human liver cancer cells via mitochondrial dysfunction. Cells 2021, 10, 332. [Google Scholar] [CrossRef] [PubMed]
- Schepici, G.; Bramanti, P.; Mazzon, E. Efficacy of sulforaphane in neurodegenerative diseases. Int. J. Mol. Sci. 2020, 21, 8637. [Google Scholar] [CrossRef]
- Kang, B.; Kim, C.Y.; Hwang, J.; Suh, H.J.; Choi, H.S. Brassinin, a phytoalexin in cruciferous vegetables, suppresses obesity-induced inflammatory responses through the Nrf2-HO-1 signaling pathway in an adipocyte-macrophage co-culture system. Phytother. Res. 2019, 33, 1426–1437. [Google Scholar] [CrossRef]
- Kang, B.; Hwang, J.; Choi, H.-S. Brassinin, a brassica-derived phytochemical, regulates monocyte-to-macrophage differentiation and inflammatory responses in human monocytes and murine macrophages. J. Pharm. Pharmacol. 2020, 72, 1245–1255. [Google Scholar] [CrossRef]
- Yang, M.H.; Lee, J.H.; Ko, J.H.; Jung, S.H.; Sethi, G.; Ahn, K.S. Brassinin Represses Invasive Potential of Lung Carcinoma Cells through Deactivation of PI3K/Akt/mTOR Signaling Cascade. Molecules 2019, 24, 1584. [Google Scholar] [CrossRef]
- Yang, M.H.; Baek, S.H.; Ha, I.J.; Um, J.Y.; Ahn, K.S. Brassinin enhances the anticancer actions of paclitaxel by targeting multiple signaling pathways in colorectal cancer cells. Phytother. Res. 2021, 35, 3875–3885. [Google Scholar] [CrossRef]
- Lee, M.K.; Hwang, Y.H.; Ryu, H.; Lee, A.; Jeong, H.H.; Baek, J.; Kim, M.J.; Lee, J.Y.; Van, J.Y.; Liu, Y.; et al. Galla rhois water extract inhibits enzymatic browning in apple juice partly by binding to and inactivating polyphenol oxidase. Food Chem. 2022, 383, 132277. [Google Scholar] [CrossRef]
- Moon, K.M.; Yang, J.H.; Lee, M.K.; Kwon, E.B.; Baek, J.; Hwang, T.; Kim, J.I.; Lee, B. Maclurin Exhibits Antioxidant and Anti-Tyrosinase Activities, Suppressing Melanogenesis. Antioxidants 2022, 11, 1164. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gwon, W.-G.; Lee, S.-G.; Kim, J.-I.; Kim, Y.-M.; Kim, S.-B.; Kim, H.-R. Hexane fraction from the ethanolic extract of Sargassum serratifolium suppresses cell adhesion molecules via regulation of NF-κB and Nrf2 pathway in human umbilical vein endothelial cells. Fish. Aquat. Sci. 2019, 22, 7. [Google Scholar] [CrossRef]
- Lee, B.; Moon, K.M.; Lee, B.S.; Yang, J.H.; Park, K.I.; Cho, W.K.; Ma, J.Y. Swertiajaponin inhibits skin pigmentation by dual mechanisms to suppress tyrosinase. Oncotarget 2017, 8, 95530–95541. [Google Scholar] [CrossRef]
- Arias, E.; Gonzalez, J.; Peiró, J.; Oria, R.; Lopez-Buesa, P. Browning prevention by ascorbic acid and 4-hexylresorcinol: Different mechanisms of action on polyphenol oxidase in the presence and in the absence of substrates. J. Food Sci. 2007, 72, C464–C470. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-K.; Lee, G.-D.; Kwon, J.-H.; Kim, K.-H. Monitoring on extraction yields and functional properties of Brassica oleracea var. capita extracts. Food Sci. Biotechnol. 2005, 14, 836–840. [Google Scholar]
- Vachtenheim, J.; Borovansky, J. “Transcription physiology” of pigment formation in melanocytes: Central role of MITF. Exp. Derm. 2010, 19, 617–627. [Google Scholar] [CrossRef]
- Yin, W.-M.; Cao, X.-B.; Li, S.-X.; Zhang, F.; Guan, Y.-F. Brassinin inhibits proliferation and induces cell cycle arrest and apoptosis in nasopharyngeal cancer C666-1 cells. Arab. J. Chem. 2022, 15, 104018. [Google Scholar] [CrossRef]
- Al-abbasy, O. The Effect of Some Antioxidants Compounds on Enzymatic and Nonenzymic Browning of Quince Juice During Thermal Treatment. J. Educ. Sci. 2020, 29, 206–221. [Google Scholar]
- Yuriev, E.; Agostino, M.; Ramsland, P.A. Challenges and advances in computational docking: 2009 in review. J. Mol. Recognit. JMR 2011, 24, 149–164. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-K.; Ryu, H.; Jeong, H.H.; Lee, B. Brassinin Abundant in Brassicaceae Suppresses Melanogenesis through Dual Mechanisms of Tyrosinase Inhibition. Foods 2023, 12, 121. https://doi.org/10.3390/foods12010121
Lee M-K, Ryu H, Jeong HH, Lee B. Brassinin Abundant in Brassicaceae Suppresses Melanogenesis through Dual Mechanisms of Tyrosinase Inhibition. Foods. 2023; 12(1):121. https://doi.org/10.3390/foods12010121
Chicago/Turabian StyleLee, Min-Kyeong, Heeyeon Ryu, Hyeon Hak Jeong, and Bonggi Lee. 2023. "Brassinin Abundant in Brassicaceae Suppresses Melanogenesis through Dual Mechanisms of Tyrosinase Inhibition" Foods 12, no. 1: 121. https://doi.org/10.3390/foods12010121
APA StyleLee, M.-K., Ryu, H., Jeong, H. H., & Lee, B. (2023). Brassinin Abundant in Brassicaceae Suppresses Melanogenesis through Dual Mechanisms of Tyrosinase Inhibition. Foods, 12(1), 121. https://doi.org/10.3390/foods12010121