
Tilapia-Head Chondroitin Sulfate Protects against Nonalcoholic Fatty Liver Disease via Modulating the Gut–Liver Axis in High-Fat-Diet-Fed C57BL/6 Mice

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of TH-CS
2.3. Characterization of TH-CS
2.3.1. FT-IR Analysis
2.3.2. TH-CS Molecular Weight Determination
2.3.3. Monosaccharide Composition Determination
2.3.4. Nuclear Magnetic Resonance (NMR) Spectroscopy
2.4. Animals and Treatment
2.5. Determination of IR Index
2.6. Serum and Hepatic Biochemical Assay
2.7. Haematoxylin and Eosin (H&E) Staining and Oil Red O Staining
2.8. Gut Microbiota Analysis
2.9. Quantification of SCFAs
2.10. Quantitative RT-PCR
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
3.1. Chemical Properties of TH-CS
3.2. TH-CS Prevented Obesity in NAFLD Mice
3.3. TH-CS Alleviated Liver Function in NAFLD Mice
3.4. TH-CS Alleviated IR in NAFLD Mice
3.5. TH-CS Attenuated Lipid Accumulation in the Sera and Livers of NAFLD Mice
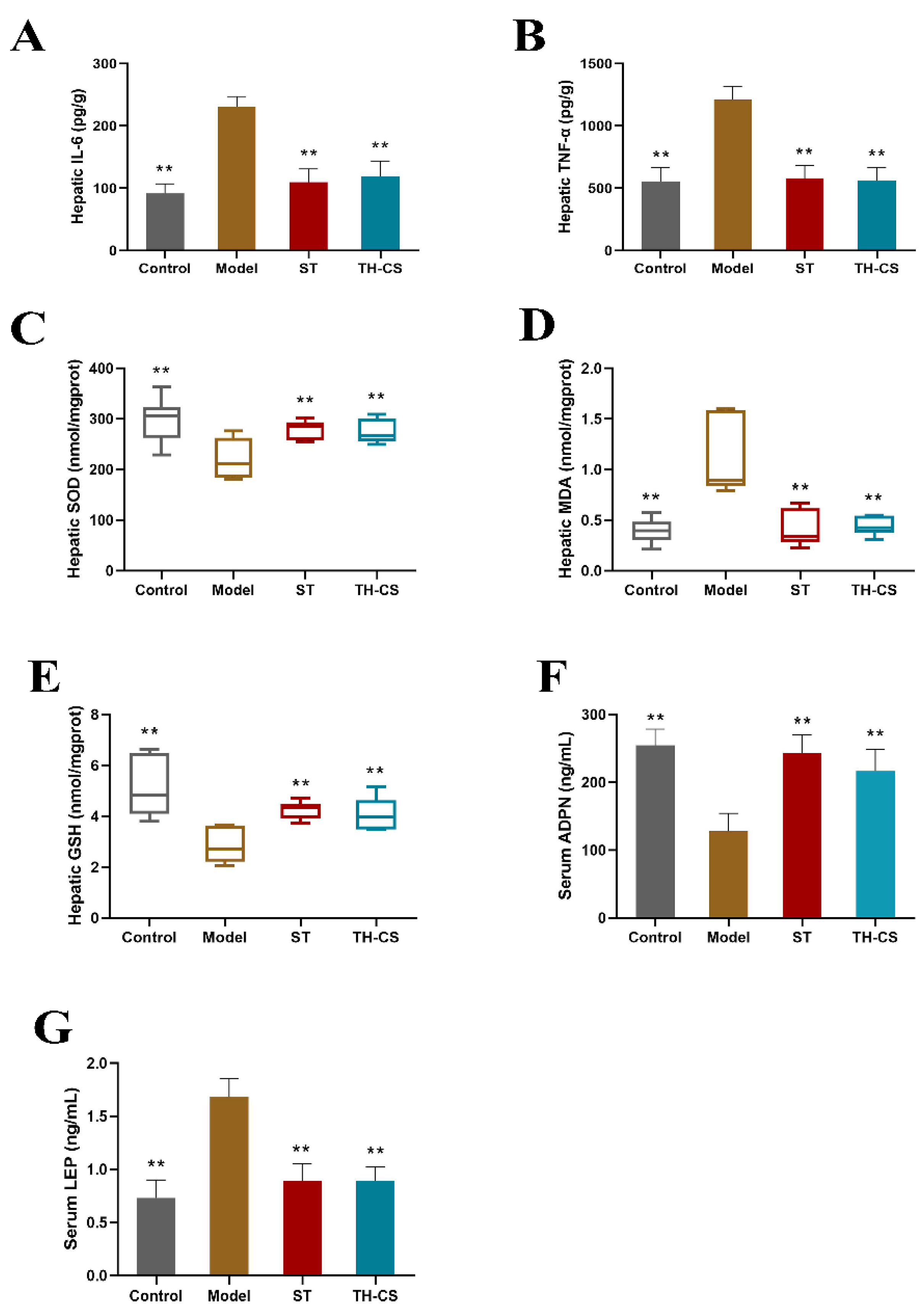
3.6. TH-CS Reduced Liver Inflammation in NAFLD Mice
3.7. TH-CS Regulated Hepatic Oxidative Stress in NAFLD Mice
3.8. TH-CS Affected Obesity Factors in NAFLD Mice
3.9. TH-CS Improved Gut Microbiota Composition in NAFLD Mice
3.10. TH-CS Regulated the Levels of SCFAs in NAFLD Mice
3.11. TH-CS Inhibited Hepatic Lipid Accumulation via Modulation of Metabolic Gene Expression in NAFLD Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Milic, S.; Lulic, D.; Stimac, D. Non-alcoholic fatty liver disease and obesity: Biochemical, metabolic and clinical presentations. World J. Gastroenterol. 2014, 20, 9330–9337. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Y. Review article: Is non-alcoholic fatty liver disease a spectrum, or are steatosis and non-alcoholic steatohepatitis distinct conditions? Aliment. Pharmacol. Ther. 2012, 36, 815–823. [Google Scholar] [CrossRef]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metab.-Clin. Exp. 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, L.; Chen, L.; Lin, X.; Xu, Y.; Ren, J.; Fu, J.; Qiu, Y. Effect of oleoylethanolamide on diet-induced nonalcoholic fatty liver in rats. J. Pharmacol. Sci. 2015, 127, 244–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.; Rivera, L.; Furness, J.B.; Angus, P.W. The role of the gut microbiota in NAFLD. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 412–425. [Google Scholar] [CrossRef]
- Magni, P.; Macchi, C.; Morlotti, B.; Sirtori, C.R.; Ruscica, M. Risk identification and possible countermeasures for muscle adverse effects during statin therapy. Eur. J. Intern. Med. 2015, 26, 82–88. [Google Scholar] [CrossRef]
- Cheng, F.; Yan, X.; Zhang, M.; Chang, M.; Yun, S.; Meng, J.; Liu, J.; Feng, C.P. Regulation of RAW 264.7 cell-mediated immunity by polysaccharides from Agaricus blazei Murill via the MAPK signal transduction pathway. Food Funct. 2017, 8, 1475–1480. [Google Scholar] [CrossRef]
- Orak, H.H.; Yagar, H.; Isbilir, S.S. Comparison of antioxidant activities of juice, peel, and seed of pomegranate (Punica granatum L.) and inter-relationships with total phenolic, Tannin, anthocyanin, and flavonoid contents. Food Sci. Biotechnol. 2012, 21, 373–387. [Google Scholar] [CrossRef]
- Li, D.; Cui, Y.; Wang, X.; Liu, F.; Li, X. Apple Polyphenol Extract Improves High-Fat Diet-Induced Hepatic Steatosis by Regulating Bile Acid Synthesis and Gut Microbiota in C57BL/6 Male Mice. J. Agric. Food Chem. 2021, 69, 6829–6841. [Google Scholar] [CrossRef]
- Shang, Q.; Shi, J.; Song, G.; Zhang, M.; Cai, C.; Hao, J.; Li, G.; Yu, G. Structural modulation of gut microbiota by chondroitin sulfate and its oligosaccharide. Int. J. Biol. Macromol. 2016, 89, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Betti, M.; Pietrasik, Z. Extraction, isolation and analysis of chondroitin sulfate glycosaminoglycans. Recent Pat. Food Nutr. Agric. 2010, 2, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Therapeutic importance of sulfated polysaccharides from seaweeds: Updating the recent findings. 3 Biotech 2012, 2, 171–185. [Google Scholar] [CrossRef] [Green Version]
- Martel-Pelletier, J.; Farran, A.; Montell, E.; Verges, J.; Pelletier, J.-P. Discrepancies in Composition and Biological Effects of Different Formulations of Chondroitin Sulfate. Molecules 2015, 20, 4277–4289. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Kobayashi, T.; Moroi, S.; Kotake, H.; Ikoma, T.; Saeki, H.; Ura, K.; Takagi, Y. Anti-obesity effects of chondroitin sulfate oligosaccharides from the skate Raja pulchra. Carbohydr. Polym. 2019, 214, 303–310. [Google Scholar] [CrossRef]
- Medeiros, L.H.C.; Vasconcelos, B.M.F.; Silva, M.B.; Souza-Junior, A.A.; Chavante, S.F.; Andrade, G.P.V. Chondroitin sulfate from fish waste exhibits strong intracellular antioxidant potential. Braz. J. Med. Biol. Res. 2021, 54. [Google Scholar] [CrossRef]
- Kosakai, M.; Yosizawa, Z. A partial modification of the carbazole method of Bitter and Muir for quantitation of hexuronic acids. AnBio 1979, 93, 295–298. [Google Scholar] [CrossRef]
- Sarzanini, C.; Bruzzoniti, M.C.; Mentasti, E. Determination of epichlorohydrin by ion chromatography. J. Chromatogr. A 2000, 884, 251–259. [Google Scholar] [CrossRef]
- Ma, P.; Sun, C.; Li, W.; Deng, W.; Adu-Frimpong, M.; Yu, J.; Xu, X. Extraction and structural analysis of Angelica sinensis polysaccharide with low molecular weight and its lipid-lowering effect on nonalcoholic fatty liver disease. Food Sci. Nutr. 2020, 8, 3212–3224. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Q.; Zhang, C.; Jia, W.; Han, L.; Yu, Q. Chicken leg bone as a source of chondroitin sulfate. Carbohydr. Polym. 2019, 207, 191–199. [Google Scholar] [CrossRef]
- Bougatef, H.; Krichen, F.; Capitani, F.; Ben Amor, I.; Gargouri, J.; Maccari, F.; Mantovani, V.; Galeotti, F.; Volpi, N.; Bougatef, A.; et al. Purification, compositional analysis, and anticoagulant capacity of chondroitin sulfate/dermatan sulfate from bone of corb (Sciaena umbra). Int. J. Biol. Macromol. 2019, 134, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.M.; Ashraf, M.; Hashmi, A.S.; Ahmad, M.-u.-D.; Anjum, A.A. Extraction and Biochemical Characterization of Sulphated Glycosaminoglycans from Chicken Keel Cartilage. Pak. Vet. J. 2013, 33, 471–475. [Google Scholar] [CrossRef]
- Vázquez, J.; Fraguas, J.; Novoa-Carballal, R.; Reis, R.L.; Pérez-Martín, R.I.; Valcarcel, J.J.C.P. Optimal isolation and characterisation of chondroitin sulfate from rabbit fish (Chimaera monstrosa). Carbohydr. Polym. 2019, 210, 302–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novoa-Carballal, R.; Pérez-Martín, R.; Blanco, M.; Sotelo, C.G.; Fassini, D.; Nunes, C.; Coimbra, M.A.; Silva, T.H.; Reis, R.L.; Vázquez, J.A.J.C.P. By-products of Scyliorhinus canicula, Prionace glauca and Raja clavata: A valuable source of predominantly 6S sulfated chondroitin sulfate. Carbohydr. Polym. 2017, 157, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.-H.; Hashimoto, N.; Shimada, K.-I.; Sekikawa, M.; Noda, T.; Yamauchi, H.; Hashimoto, M.; Chiji, H.; Topping, D.L.; Fukushima, M. Hepatoprotective effects of purple potato extract against D-galactosamine-induced liver injury in rats. Biosci. Biotechnol. Biochem. 2006, 70, 1432–1437. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Fan, R.; Yin, S.; Zhao, X.; Liu, J.; Li, L.; Zhang, W.; Ge, L. Protective effects of Ginkgo biloba leaf polysaccharide on nonalcoholic fatty liver disease and its mechanisms. Int. J. Biol. Macromol. 2015, 80, 573–580. [Google Scholar] [CrossRef]
- Feldstein, A.E.; Werneburg, N.W.; Canbay, A.; Guicciardi, M.E.; Bronk, S.F.; Rydzewski, R.; Burgart, L.J.; Gores, G.J. Free fatty acids promote hepatic lipotoxicity by stimulating TNF-alpha expression via a lysosomal pathway. Hepatology 2004, 40, 185–194. [Google Scholar] [CrossRef]
- Heeba, G.H.; Morsy, M.A. Fucoidan ameliorates steatohepatitis and insulin resistance by suppressing oxidative stress and inflammatory cytokines in experimental non-alcoholic fatty liver disease. Environ. Toxicol. Pharmacol. 2015, 40, 907–914. [Google Scholar] [CrossRef]
- Ding, X.; Jian, T.; Li, J.; Lv, H.; Tong, B.; Li, J.; Meng, X.; Ren, B.; Chen, J. Chicoric Acid Ameliorates Nonalcoholic Fatty Liver Disease via the AMPK/Nrf2/NF kappa B Signaling Pathway and Restores Gut Microbiota in High-Fat-Diet-Fed Mice. Oxid. Med. Cell. Longev. 2020, 2020, 9734560. [Google Scholar] [CrossRef]
- Yu, Y.; Shen, M.; Song, Q.; Xie, J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydr. Polym. 2018, 183, 91–101. [Google Scholar] [CrossRef]
- Li, W.; Huang, D.; Gao, A.; Yang, X. Stachyose increases absorption and hepatoprotective effect of tea polyphenols in high fructose-fed mice. Mol. Nutr. Food Res. 2016, 60, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, A.M.; German, J.B.; Sanyal, A.J. Comparative review of diets for the metabolic syndrome: Implications for nonalcoholic fatty liver disease. Am. J. Clin. Nutr. 2007, 86, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuniga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Cen, L.; Zhou, T.; Yu, M.; Chen, X.; Jiang, W.; Li, Y.; Yu, C.; Shen, Z. Insulin-like growth factor binding protein 1 ameliorates lipid accumulation and inflammation in nonalcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2021. [Google Scholar] [CrossRef]
- Kim, K.-J.; Yoon, K.-Y.; Lee, B.-Y. Fucoidan regulate blood glucose homeostasis in C57BL/KSJ m+/+db and C57BL/KSJ db/db mice. Fitoterapia 2012, 83, 1105–1109. [Google Scholar] [CrossRef]
- Seol, B.G.; Kim, J.H.; Woo, M.; Song, Y.O.; Choi, Y.H.; Noh, J.S.; Cho, E.J. Skate cartilage extracts containing chondroitin sulfate ameliorates hyperlipidemia-induced inflammation and oxidative stress in high cholesterol diet-fed LDL receptor knockout mice in comparison with shark chondroitin sulfate. Nutr. Res. Pract. 2020, 14, 175–187. [Google Scholar] [CrossRef]
- Wu, S.; Liu, Y.L.; Jiang, P.R.; Xu, Y.X.; Zheng, W.Y.; Song, S.; Ai, C.Q. Effect of sulfate group on sulfated polysaccharides-induced improvement of metabolic syndrome and gut microbiota dysbiosis in high fat diet-fed mice. Int. J. Biol. Macromol. 2020, 164, 2062–2072. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Jiang, H.; Cai, C.; Li, G.; Hao, J.; Yu, G. Marine polysaccharides attenuate metabolic syndrome by fermentation products and altering gut microbiota: An overview. Carbohydr. Polym. 2018, 195, 601–612. [Google Scholar] [CrossRef]
- Kindt, A.; Liebisch, G.; Clavel, T.; Haller, D.; Hoermannsperger, G.; Yoon, H.; Kolmeder, D.; Sigruener, A.; Krautbauer, S.; Seeliger, C.; et al. The gut microbiota promotes hepatic fatty acid desaturation and elongation in mice. Nat. Commun. 2018, 9, 3760. [Google Scholar] [CrossRef] [Green Version]
- Dahlhoff, C.; Worsch, S.; Sailer, M.; Hummel, B.A.; Fiamoncini, J.; Uebel, K.; Obeid, R.; Scherling, C.; Geisel, J.; Bader, B.L.; et al. Methyl-donor supplementation in obese mice prevents the progression of NAFLD, activates AMPK and decreases acyl-carnitine levels. Mol. Metab. 2014, 3, 565–580. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Mao, G.; Hu, Y.; Ye, X.; Tian, D.; Linhardt, R.J.; Chen, S. Fucosylated chondroitin sulfate oligosaccharides from Isostichopus badionotus regulates lipid disorder in C57BL/6 mice fed a high-fat diet. Carbohydr. Polym. 2018, 201, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Sikder, K.; Shukla, S.K.; Patel, N.; Singh, H.; Rafiq, K. High Fat Diet Upregulates Fatty Acid Oxidation and Ketogenesis via Intervention of PPAR-gamma. Cell. Physiol. Biochem. 2018, 48, 1317–1331. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GPR41 | ACCACTATTTACCTCACCTCCCTCTTC | CGATGCCAGGAACCAACAGACTAC |
| GPR43 | TGCACCATCGTCATCATCGTTCAG | AGTTCTCGTAGCAGGTTATTTGGTTCTC |
| AMPK | TCCACAGAGATCGGGATCAGTTAGC | GAGTTAGGTCAACAGGAGAAGAGTCAAG |
| PPARγ | AGCCCTTTACCACAGTTGATTTCTCC | GCAGGTTCTACTTTGATCGCACTTTG |
| SREBP-1c | CCTGCTTGGCTCTTCTCTTTGTCTAC | AGGTCAGCTTGTTTGCGATGTCTC |
| FAS | GTTTAAAGCTGAGGAGGCGGGTTC | GTTTTCAGGTTGGCATGGTTGACAG |
| GAPDH | AAGAAGGTGGTGAAGCAGGCATC | CGGCATCGAAGGTGGAAGAGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Yi, X.; Gao, X.; Ji, J.; Liu, Z.; Xia, G.; Li, C.; Zhang, X.; Shen, X. Tilapia-Head Chondroitin Sulfate Protects against Nonalcoholic Fatty Liver Disease via Modulating the Gut–Liver Axis in High-Fat-Diet-Fed C57BL/6 Mice. Foods 2022, 11, 922. https://doi.org/10.3390/foods11070922
Yu H, Yi X, Gao X, Ji J, Liu Z, Xia G, Li C, Zhang X, Shen X. Tilapia-Head Chondroitin Sulfate Protects against Nonalcoholic Fatty Liver Disease via Modulating the Gut–Liver Axis in High-Fat-Diet-Fed C57BL/6 Mice. Foods. 2022; 11(7):922. https://doi.org/10.3390/foods11070922
Chicago/Turabian StyleYu, Hui, Xiangzhou Yi, Xia Gao, Jun Ji, Zhongyuan Liu, Guanghua Xia, Chuan Li, Xueying Zhang, and Xuanri Shen. 2022. "Tilapia-Head Chondroitin Sulfate Protects against Nonalcoholic Fatty Liver Disease via Modulating the Gut–Liver Axis in High-Fat-Diet-Fed C57BL/6 Mice" Foods 11, no. 7: 922. https://doi.org/10.3390/foods11070922
APA StyleYu, H., Yi, X., Gao, X., Ji, J., Liu, Z., Xia, G., Li, C., Zhang, X., & Shen, X. (2022). Tilapia-Head Chondroitin Sulfate Protects against Nonalcoholic Fatty Liver Disease via Modulating the Gut–Liver Axis in High-Fat-Diet-Fed C57BL/6 Mice. Foods, 11(7), 922. https://doi.org/10.3390/foods11070922