
Characterization of Microbiological Quality of Whole and Gutted Baltic Herring


Abstract
:1. Introduction
2. Materials and Methods
2.1. Herring Samples
2.2. Culture-Based Microbiological Analyses
2.3. Identification of Bacterial Isolates
2.3.1. MALDI-TOF MS Analysis
2.3.2. 16S rRNA Gene Sequencing of Bacterial Isolates
2.3.3. DNA-Isolation and Amplicon Library Preparation
2.3.4. Sequence Processing and Data Analyses
2.4. Statistical Analysis and Visualization
3. Results
3.1. Viable Counts
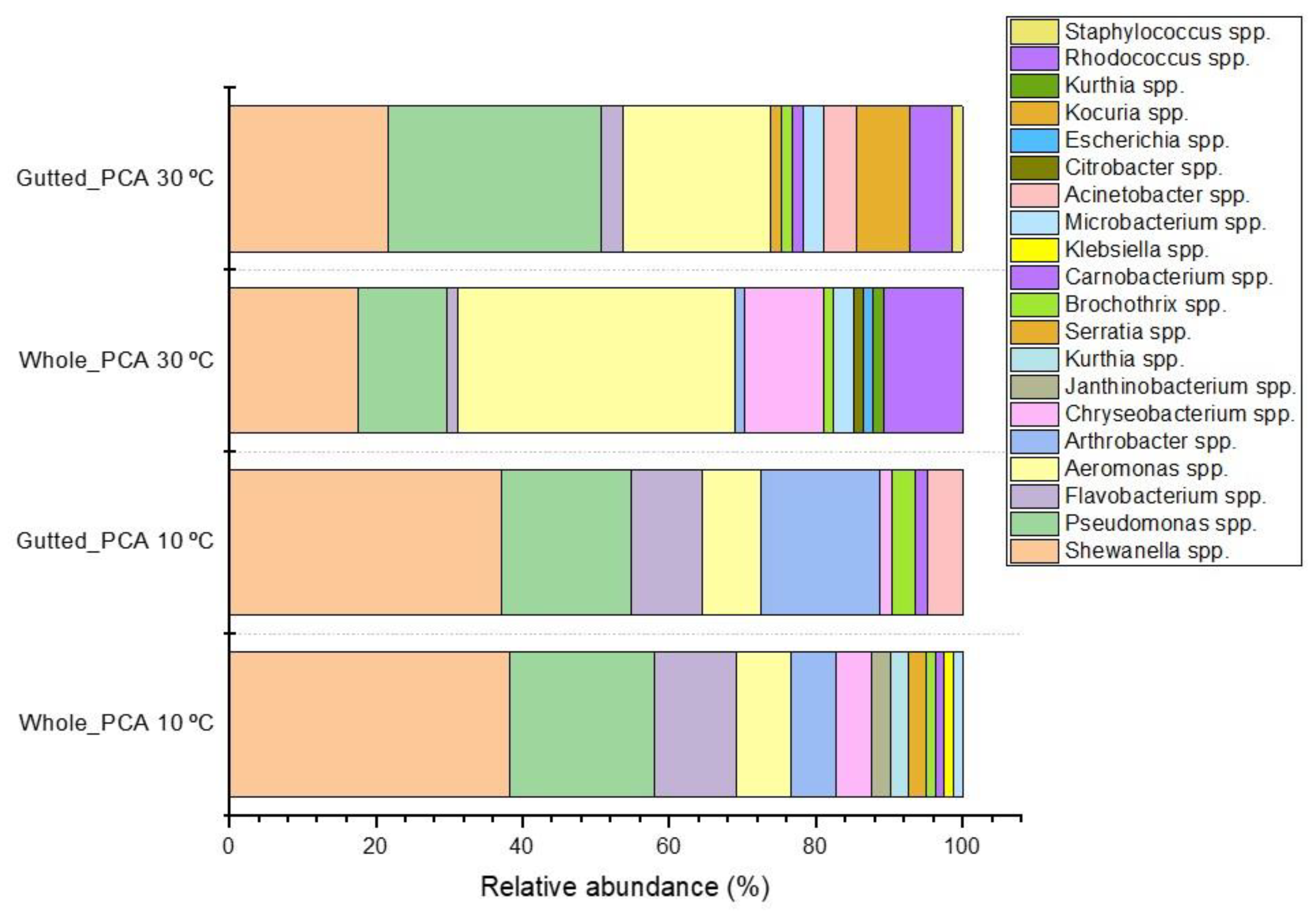
3.2. Identification of Bacterial Isolates
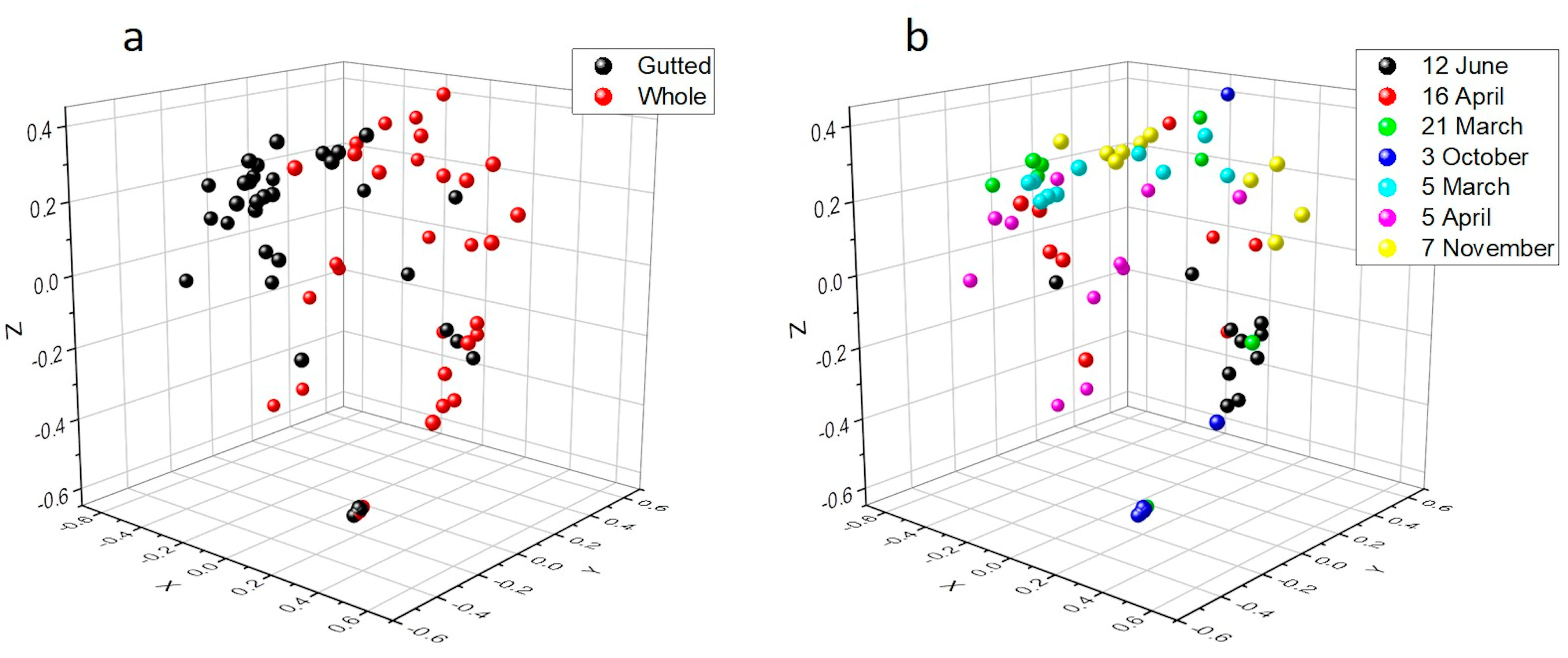
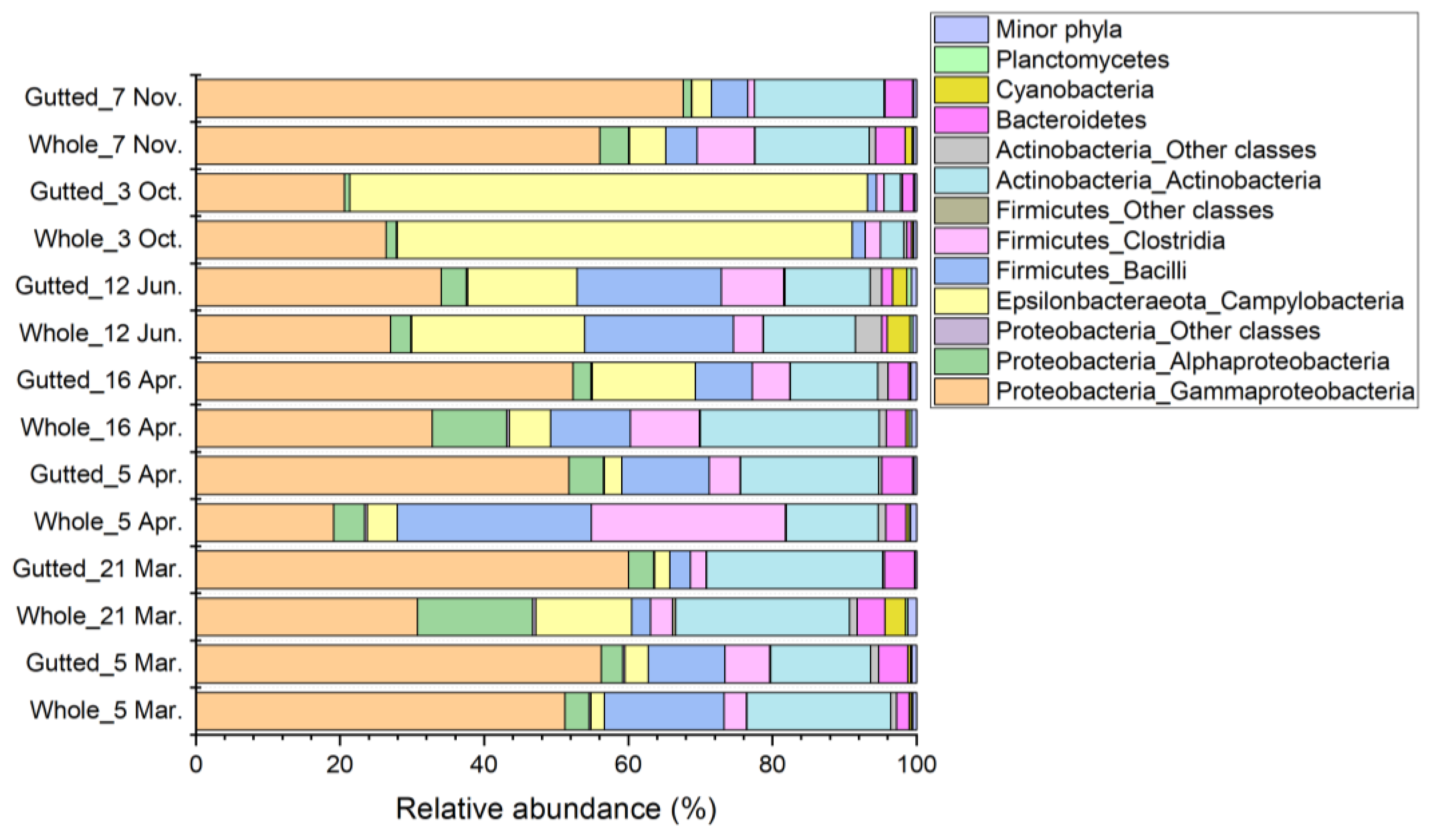
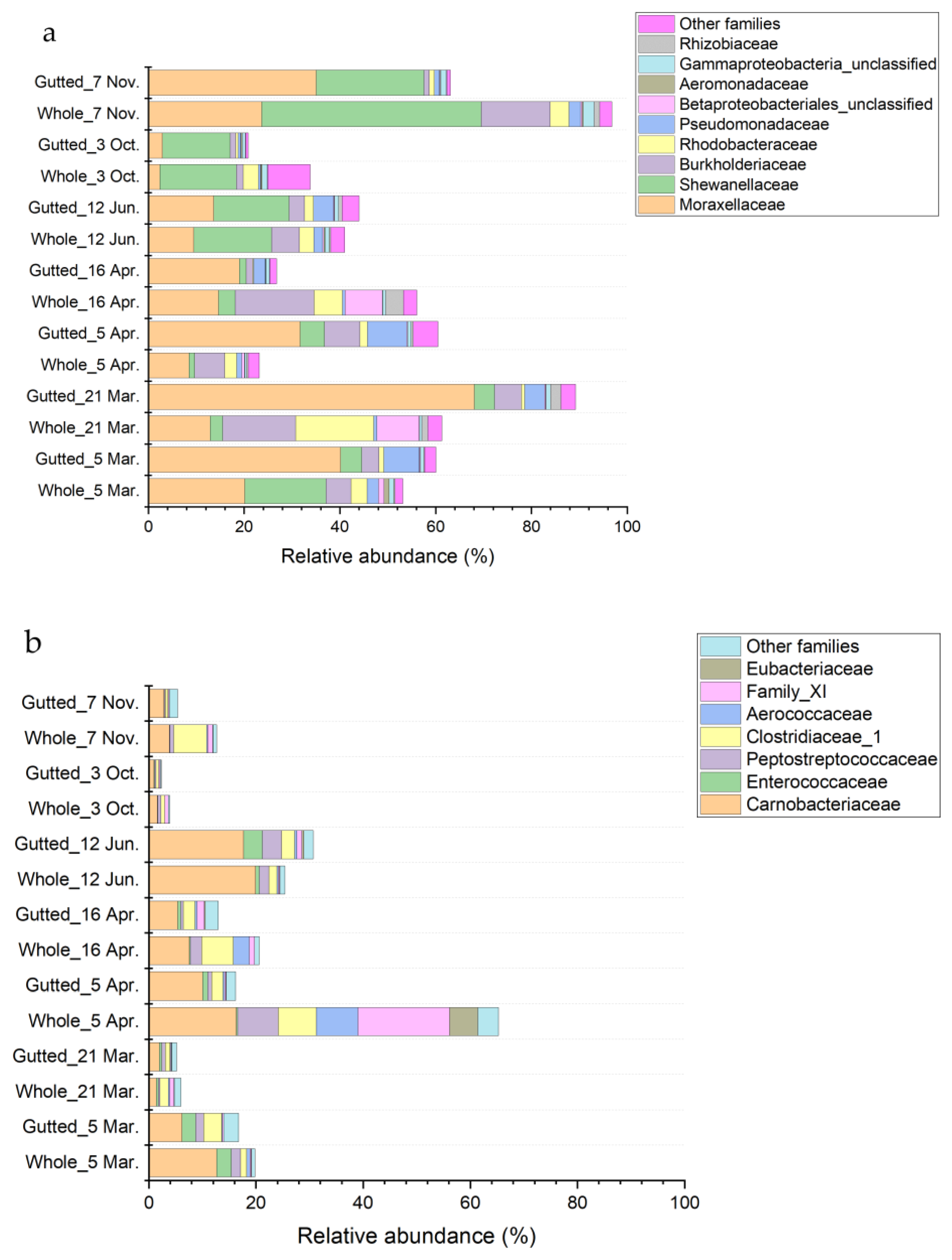
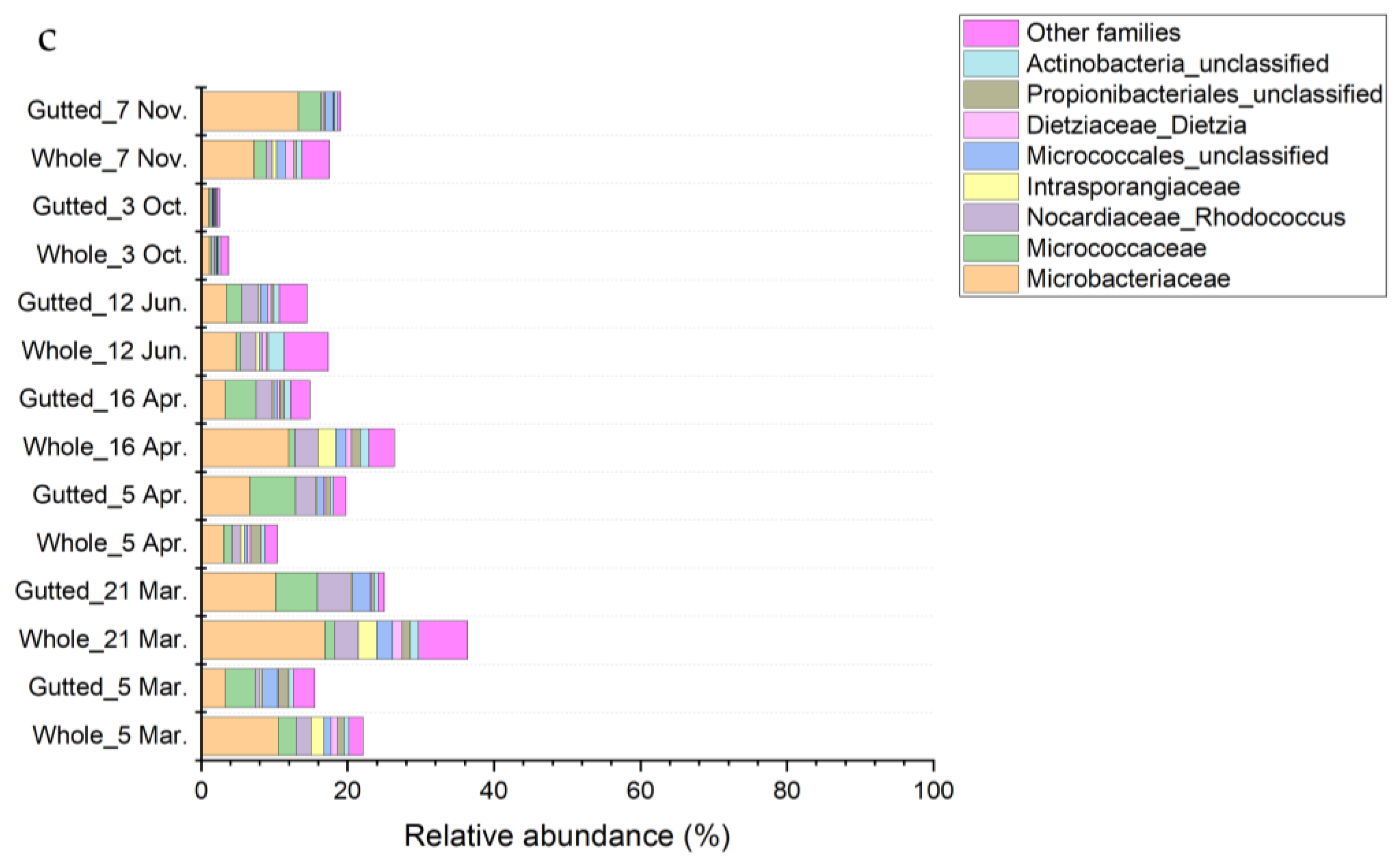
3.3. Culture-Independent Population Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Natural Resources Institute Finland. Statistics Database. Available online: http://statdb.luke.fi/PXWeb/pxweb/en/LUKE/ (accessed on 22 December 2021).
- Setälä, J.; Saarni, K.; Niukko, J. Fish Market Review 2017. Available online: https://www.luke.fi/wp-content/uploads/2019/05/Fish-market-review-2017.pdf (accessed on 3 January 2022).
- Richie, H.; Roser, M. Biodiversity. Our World in Data. Available online: https://ourworldindata.org/biodiversity (accessed on 20 December 2021).
- Nisov, A.; Aisala, H.; Holopainen-Mantila, U.; Alakomi, H.-L.; Nordlund, E.; Honkapää, K. Comparison of Whole and Gutted Baltic Herring as a Raw Material for Restructured Fish Product Produced by High-Moisture Extrusion Cooking. Foods 2020, 9, 1541. [Google Scholar] [CrossRef] [PubMed]
- Boziaris, I.S.; Parlapani, F.F. Specific Spoilage Organisms (SSOs) in Fish; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Gram, L.; Huss, H.H. Microbiological spoilage of fish and fish products. Int. J. Food Microbiol. 1996, 33, 121–137. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Michailidou, S.; Pasentsis, K.; Argiriou, A.; Krey, G.; Boziaris, I.S. A meta-barcoding approach to assess and compare the storage temperature-dependent bacterial diversity of gilt-head sea bream (Sparus aurata) originating from fish farms from two geographically distinct areas of Greece. Int. J. Food Microbiol. 2018, 278, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Wedell-Neergaard, C.; Huss, H.H. The bacteriology of fresh and spoiling Lake Victorian Nile perch (Lates niloticus). Int. J. Food Microbiol. 1990, 10, 303–316. [Google Scholar] [CrossRef]
- Gram, L.; Dalgaard, P. Fish spoilage bacteria—Problems and solutions. Curr. Opin. Biotechnol. 2002, 13, 262–266. [Google Scholar] [CrossRef]
- Papadopoulos, V.; Chouliara, I.; Badeka, A.; Savvaidis, I.; Kontominas, M. Effect of gutting on microbiological, chemical, and sensory properties of aquacultured sea bass (Dicentrarchus labrax) stored in ice. Food Microbiol. 2003, 20, 411–420. [Google Scholar] [CrossRef]
- Fogarty, C.; Whyte, P.; Brunton, N.; Lyng, J.; Smyth, C.; Fagan, J.; Bolton, D. Spoilage indicator bacteria in farmed Atlantic salmon (Salmo salar) stored on ice for 10 days. Food Microbiol. 2018, 77, 38–42. [Google Scholar] [CrossRef]
- Höll, L.; Behr, J.; Vogel, R.F. Identification and growth dynamics of meat spoilage microorganisms in modified atmosphere packaged poultry meat by MALDI-TOF MS. Food Microbiol. 2016, 60, 84–91. [Google Scholar] [CrossRef]
- Chaillou, S.; Chaulot-Talmon, A.; Caekebeke, H.; Cardinal, M.; Christieans, S.; Denis, C.; Desmonts, M.H.; Dousset, X.; Feurer, C.; Hamon, E.; et al. Origin and ecological selection of core and food-specific bacterial communities associated with meat and seafood spoilage. ISME J. 2015, 9, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Peruzy, M.F.; Dumolin, C.; Joossens, M.; Houf, K. Assessment of food microbiological indicators applied on poultry carcasses by culture combined MALDI-TOF MS identification and 16S rRNA amplicon sequencing. Food Microbiol. 2019, 82, 53–61. [Google Scholar] [CrossRef]
- Peruzy, M.; Murru, N.; Yu, Z.; Cnockaert, M.; Joossens, M.; Proroga, Y.; Houf, K. Determination of the microbiological contamination in minced pork by culture dependent and 16S amplicon sequencing analysis. Int. J. Food Microbiol. 2018, 290, 27–35. [Google Scholar] [CrossRef]
- Peruzy, M.; Murru, N.; Yu, Z.; Kerkhof, P.-J.; Neola, B.; Joossens, M.; Proroga, Y.; Houf, K. Assessment of microbial communities on freshly killed wild boar meat by MALDI-TOF MS and 16S rRNA amplicon sequencing. Int. J. Food Microbiol. 2019, 301, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Lyhs, U.; Lahtinen, J.; Schelvissmit, R. Microbiological quality of maatjes herring stored in air and under modified atmosphere at 4 and 10 °C. Food Microbiol. 2007, 24, 508–516. [Google Scholar] [CrossRef]
- Özogul, F.; Taylor, K.D.A.; Quantick, P.; Özogul, Y. Chemical, microbiological and sensory evaluation of Atlantic herring (Clupea harengus) stored in ice, modified atmosphere and vacuum pack. Food Chem. 2000, 71, 267–273. [Google Scholar] [CrossRef]
- Karim, N.U.; Kennedy, T.; Linton, M.; Watson, S.; Gault, N.; Patterson, M.F. Effect of high pressure processing on the quality of herring (Clupea harengus) and haddock (Melanogrammus aeglefinus) stored on ice. Food Control 2011, 22, 476–484. [Google Scholar] [CrossRef]
- Albertos, I.; Martin-Diana, A.B.; Cullen, P.; Tiwari, B.K.; Ojha, K.S.; Bourke, P.; Rico, D. Shelf-life extension of herring (Clupea harengus) using in-package atmospheric plasma technology. Innov. Food Sci. Emerg. Technol. 2019, 53, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.; Zhou, X.; Xiao, H.; Miao, J.; Zhao, L. Characterization of Bacterial Microbiota in Tilapia Fillets Under Different Storage Temperatures. J. Food Sci. 2019, 84, 1487–1493. [Google Scholar] [CrossRef]
- Nordic Committee on Food Analysis. Enterobacteriaceae. Determination in Foods and Feeds. Method 144, 2nd ed.; NMKL Secretariat: Oslo, Norway, 2000. [Google Scholar]
- Jeong, S.; Hong, J.S.; Kim, J.O.; Kim, K.-H.; Lee, W.; Bae, I.K.; Lee, K.; Jeong, S.H. Identification of Acinetobacter Species Using Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. Ann. Lab. Med. 2016, 36, 325–334. [Google Scholar] [CrossRef] [Green Version]
- BLAST: Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 1 November 2021).
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westcott, S.L.; Schloss, P.D. De novo clustering methods outperform reference-based methods for assigning 16S rRNA gene sequences to operational taxonomic units. PeerJ 2015, 12, e1487. [Google Scholar] [CrossRef]
- Finnish Food and Drink Industries’ Federation. Elintarvikkeiden Mikrobiologisia Ohjausarvoja Viimeisenä Käyttöpäivänä; Finnish Food and Drink Industries’ Federation: Helsinki, Finland, 2017. [Google Scholar]
- Vogel, B.F.; Venkateswaran, K.; Satomi, M.; Gram, L. Identification of Shewanella baltica as the Most Important H2S-Producing Species during Iced Storage of Danish Marine Fish. Appl. Environ. Microbiol. 2005, 71, 6689–6697. [Google Scholar] [CrossRef] [Green Version]
- Erkan, N.; Özden, Ö. Quality assessment of whole and gutted sardines (Sardina pilchardus) stored in ice. Int. J. Food Sci. Technol. 2008, 43, 1549–1559. [Google Scholar] [CrossRef]
- Syropoulou, F.; Parlapani, F.; Kakasis, S.; Nychas, G.-J.; Boziaris, I. Primary Processing and Storage Affect the Dominant Microbiota of Fresh and Chill-Stored Sea Bass Products. Foods 2021, 10, 671. [Google Scholar] [CrossRef]
- Cakli, S.; Kilinc, B.; Cadun, A.; Dincer, T.; Tolasa, S. Effects of Gutting and Ungutting on Microbiological, Chemical, and Sensory Properties of Aquacultured Sea Bream (Sparus aurata) and Sea Bass (Dicentrarchus labrax) Stored in Ice. Crit. Rev. Food Sci. Nutr. 2006, 46, 519–527. [Google Scholar] [CrossRef]
- Tzikas, Z.; Amvrosiadis, I.; Soultos, N.; Georgakis, S. Seasonal variation in the chemical composition and microbiological condition of Mediterranean horse mackerel (Trachurus mediterraneus) muscle from the North Aegean Sea (Greece). Food Control 2007, 18, 251–257. [Google Scholar] [CrossRef]
- Chiarello, M.; Auguet, J.-C.; Bettarel, Y.; Bouvier, C.; Claverie, T.; Graham, N.; Rieuvilleneuve, F.; Sucré, E.; Bouvier, T.; Villéger, S. Skin microbiome of coral reef fish is highly variable and driven by host phylogeny and diet. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.H.; Lin, G.; Fu, G.H.; Wan, Z.Y.; Lee, M.; Wang, L.; Liu, X.J.; Yue, G.H. The intestinal microbiome of fish under starvation. BMC Genom. 2014, 15, 266. [Google Scholar] [CrossRef] [Green Version]
- Lyons, P.P.; Turnbull, J.; Dawson, K.A.; Crumlish, M. Exploring the microbial diversity of the distal intestinal lumen and mucosa of farmed rainbow trout Oncorhynchus mykiss (Walbaum) using next generation sequencing (NGS). Aquac. Res. 2015, 48, 77–91. [Google Scholar] [CrossRef]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svanevik, C.S.; Lunestad, B.T. Characterisation of the microbiota of Atlantic mackerel (Scomber scombrus). Int. J. Food Microbiol. 2011, 151, 164–170. [Google Scholar] [CrossRef] [PubMed]
- LPSN. Class Epsilonproteobacteria. Available online: https://lpsn.dsmz.de/class/epsilonproteobacteria (accessed on 13 October 2021).
- Waite, D.W.; Vanwonterghem, I.; Rinke, C.; Parks, D.H.; Zhang, Y.; Takai, K.; Sievert, S.M.; Simon, J.; Campbell, B.J.; Hanson, T.E.; et al. Comparative Genomic Analysis of the Class Epsilonproteobacteria and Proposed Reclassification to Epsilonbacteraeota (phyl. nov.). Front. Microbiol. 2017, 8, 682. [Google Scholar] [CrossRef] [PubMed]
- Patyal, A.; Rathore, R.S.; Mohan, H.V.; Dhama, K.; Kumar, A. Prevalence of Arcobacter spp. in Humans, Animals and Foods of Animal Origin Including Sea Food from India. Transbound. Emerg. Dis. 2011, 58, 402–410. [Google Scholar] [CrossRef]
- Collado, L.; Figueras, M.J. Taxonomy, Epidemiology, and Clinical Relevance of the Genus Arcobacter. Clin. Microbiol. Rev. 2011, 24, 174–192. [Google Scholar] [CrossRef] [Green Version]
- Rathlavath, S.; Kumar, S.; Nayak, B.B. Comparative isolation and genetic diversity of Arcobacter sp. from fish and the coastal environment. Lett. Appl. Microbiol. 2017, 65, 42–49. [Google Scholar] [CrossRef]
- Di Noto, A.M.; Sciortino, S.; Cardamone, C.; Ciravolo, C.; Napoli, C.; Alio, V.; Arculeo, P.; Oliveri, G.; Costa, A. Detection of Arcobacter spp. in food products collected from Sicilia region: A preliminary study. Ital. J. Food Saf. 2018, 7, 72–75. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Group | Culture Medium | Incubation Conditions |
|---|---|---|
| Aerobic mesophilic bacteria | Plate Count Agar (PCA) (BD Difco, Franklin Lakes, NJ, USA) | Aerobic, 30 °C, 3 days |
| Aerobic psychrotrophic bacteria | Plate Count Agar (PCA) (BD Difco, Franklin Lakes, NJ, USA) | Aerobic, 10 °C, 7 days |
| Spores 1 | Plate Count Agar (PCA) (BD Difco, Franklin Lakes, NJ, USA) | Aerobic, 30 °C, 3 days |
| Bacillus cereus | Mannitol Egg Yolk Polymyxin agar (Oxoid, Hampshire, UK) | Aerobic, 37 °C, 24 h |
| Hydrogen sulphide-producing bacteria | Lyngby iron agar 2 (Oxoid, Hampshire, UK) | Aerobic, 25 °C, 2 days |
| Anaerobic sulphide-reducing clostridia | Sulphite iron agar (BioLab, Tampere, Finland) | Anaerobic 3, 37 °C, 2 days |
| Lactic acid bacteria (LAB) | De Man Rogosa Sharpe agar (MRS) (Oxoid, Hampshire, UK) | Anaerobic 3, 25 °C, 5 days |
| Enterococci | mEnterococcus agar (BD Difco, Franklin Lakes, NJ, USA) | Aerobic, 37 °C, 2 days |
| Pseudomonas spp. and Aeromonas spp. | GSP selective agar (Merck, Darmstadt, Germany) | Aerobic, 28 °C, 3 days |
| Enterobacteria 4 | Violet Red Bile Glucose Agar (LabM, Lancashire, UK) | Aerobic, 37 °C, 24 h |
| Coliforms | Chromocult coliform agar (Merck, Darmstadt, Germany) | Aerobic, 37 °C, 24 h |
| Microbiological Sampling Date | Remaining Shelf Life | Aerobic Mesophiles | Aerobic Psychrotrophs | H2S-Producers | Enterobacteria | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Gutted | Whole | Gutted | Whole | Gutted | Whole | Gutted | Whole | Gutted | |
| 6 March | 1 | 3 | 4.9 ± 0.3 | 3.8 ± 0.1 | 5.3 ± 0.2 a | 4.4 ± 0.7 a | 5.5 ± 0.4 | 4.0 ± 0.1 | <1 b | 1.0 ± 1.0 b |
| 21 March | 2 | 4 | 3.5 ± 0.1 c | 3.5 ± 0.1 c | 4.0 ± 0.2 d | 3.9 ± 0.1 d | 2.4 ± 0.1 e | 2.2 ± 0.4 e | 2.2 ± 0.9 f | 1.0 ± 0.0 f |
| 5 April | 1 | 4 | 4.1 ± 0.4 | 3.3 ± 0.1 | 4.6 ± 0.2 | 3.7 ± 0.2 | 3.4 ± 0.2 g | 3.4 ± 0.4 g | <1 h | <1 h |
| 16 April | Nd 1 | Nd 1 | 3.3 ± 0.2 | 2.8 ± 0.2 | 4.3 ± 0.3 | 3.3 ± 0.2 | 4.0 ± 0.5 | 3.4 ± 0.2 | <1 | 1.7 ± 0.1 |
| 14 May | 2 | 5 | 3.3 ± 1.8 i | 3.0 ± 0.1 i | 4.3 ± 0.1 | 3.1 ± 0.3 | 4.1 ± 0.1 | 2.9 ± 0.3 | 1.1 ± 0.7 | <1 |
| 12 June | 1 | 4 | 4.6 ± 0.4 | 3.5 ± 0.2 | 4.9 ± 0.3 | 3.9 ± 0.2 | 5.0 ± 0.3 | 4.0 ± 0.3 | 1.9 ± 0.6 j | 1.7 ± 0.1 j |
| 3 October | 3 | Na 2 | 4.2 ± 0.1 | 3.8 ± 0.1 | 4.4 ± 0.2 | 4.0 ± 0.2 | 4.6 ± 0.4 | 3.8 ± 0.2 | 2.0 ± 0.4 k | 1.0 ± 1.0 k |
| 7 November | 1 | 3 | 4.3 ± 0.1 l | 4.0 ± 0.4 l | 4.5 ± 0.1 m | 4.5 ± 0.3 m | 5.5 ± 0.2 o | 5.6 ± 0.2 o | <1 p | <1 p |
| 29 May | 2 | 5 | 4.0 ± 0.2 | 3.2 ± 0.2 | 4.2 ± 0.1 | 3.5 ± 0.2 | 4.0 ± 0.1 | 3.5 ± 0.1 | <1 q | <1 q |
| 20 November | 3 | Na 2 | 4.6 ± 0.2 | 4.1 ± 0.1 | 4.7 ± 0.3 r | 4.5 ± 0.2 r | 4.6 ± 0.5 s | 4.6 ± 0.2 s | 1.1 ± 0.7 t | 1.1 ± 0.2 t |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huotari, J.; Tsitko, I.; Honkapää, K.; Alakomi, H.-L. Characterization of Microbiological Quality of Whole and Gutted Baltic Herring. Foods 2022, 11, 492. https://doi.org/10.3390/foods11040492
Huotari J, Tsitko I, Honkapää K, Alakomi H-L. Characterization of Microbiological Quality of Whole and Gutted Baltic Herring. Foods. 2022; 11(4):492. https://doi.org/10.3390/foods11040492
Chicago/Turabian StyleHuotari, Jaana, Irina Tsitko, Kaisu Honkapää, and Hanna-Leena Alakomi. 2022. "Characterization of Microbiological Quality of Whole and Gutted Baltic Herring" Foods 11, no. 4: 492. https://doi.org/10.3390/foods11040492
APA StyleHuotari, J., Tsitko, I., Honkapää, K., & Alakomi, H.-L. (2022). Characterization of Microbiological Quality of Whole and Gutted Baltic Herring. Foods, 11(4), 492. https://doi.org/10.3390/foods11040492