
Influence of Different Light Sources on the Biochemical Composition of Arthrospira spp. Grown in Model Systems



 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. Chemicals
2.3. Chemical Physical Analysis
2.3.1. pH
2.3.2. Dry Biomass Weight (DW)
2.4. Quantification of “Chlorophyll a” and Total Carotenoids
2.5. Total Lipids
2.6. Analysis of Protein Fraction
2.6.1. Total Protein
2.6.2. Phycocyanin and Allophycocyanin
2.7. Total Carbohydrates
2.8. Statistical Analysis
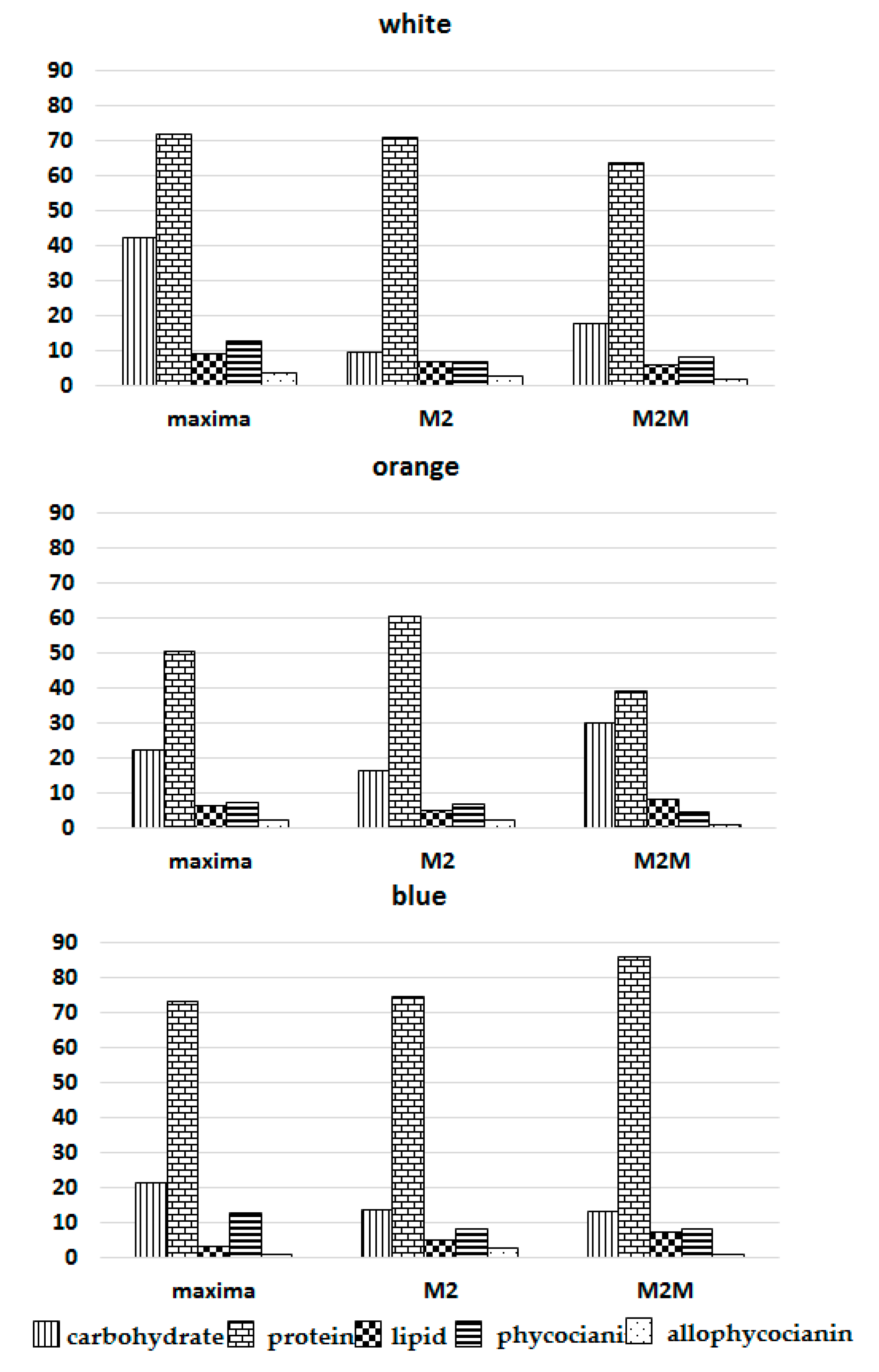
3. Results
3.1. Chemical Physical Analysis
3.2. Chlorophyll a and Total Carotenoids
3.3. Total Lipids
3.4. Analysis of Protein Fraction
3.4.1. Total Protein
3.4.2. Phycocyanin and Allophycocyanin
3.5. Total Carbohydrates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Costa, J.A.C.; Morais, M.G. Microalgae for food production. In Fermentation Process Engineering in the Food Industry; Soccol, C.R., Pandey, A., Larroche, C., Eds.; Taylor &Francis: Abingdon, UK, 2013. [Google Scholar]
- Sili, C.; Torzillo, G.; Vonshak, A. Arthrospira (Spirulina). In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer Science+Business Media B.V.: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- de Morais, M.G.; Vaz, B.D.S.; de Morais, E.G.; Costa, J.A.V. Biological effects of Spirulina (Arthrospira) biopolymers and biomass in the development of nanostructured scaffolds. BioMed Res. Int. 2014, 2014, 762705. [Google Scholar] [CrossRef] [PubMed]
- Yadav, G.; Sen, R. Microalgal green refinery concept for biosequestration of carbon dioxide vis-à-vis wastewater remediation and bioenergy production: Recent technological advances in climate research. J. CO2 Util. 2017, 17, 188–206. [Google Scholar] [CrossRef]
- Da Rosa, G.M.; Moraes, L.; Cardias, B.B.; Souza, M.D.R.A.Z.D.; Costa, J.A.V. Chemical absorption and CO2 biofixation via the cultivation of Spirulina in semicontinuous mode with nutrient recycle. Bioresour. Technol. 2015, 192, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Matos, J.; Cardoso, C.L.; Bandarra, N.M.; Afonso, C.M. Microalgae as a healthy ingredient for functional food: A review. Food Funct. 2017, 8, 2672–2685. [Google Scholar] [CrossRef]
- Mata, T.; Martins, A.; Oliveira, O.; Oliveira, S.; Mendes, A.; Caetano, N. Lipid content and productivity of Arthrospira platensis and Chlorella vulgaris under mixotrophic conditions and salinity stress. Chem. Eng. Trans. 2016, 49, 187–192. [Google Scholar]
- Sotiroudis, T.G.; Sotiroudis, G.T. Health aspects of Spirulina (Arthrospira) microalga food supplement. J. Serb. Chem. Soc. 2016, 78, 395–405. [Google Scholar] [CrossRef]
- Muhling, M.; Belay, A.; Whitton, B.A. Variation in fatty acid composition of Arthrospira (Spirulina) strains. J. Appl. Phychol. 2005, 17, 137–146. [Google Scholar] [CrossRef]
- Junga, F.; Kruger-Genge, A.; Waldeckc, P.; Kupper, J.H. Spirulina platensis, a super food? J. Cell. Biotechnol. 2019, 5, 43–54. [Google Scholar] [CrossRef]
- Léa Vernès, L.; Philippe Granvillain, P.; Farid Chemat, F.; Maryline Vian, M. Phycocyanin from Arthrospira platensis. Production, Extraction and Analysis. Curr. Biotechnol. 2015, 4, 1–11. [Google Scholar] [CrossRef]
- Liu, H.; Blankenship, R.E. On the interface of light-harvesting antenna complexes and reaction centers in oxygenic photosynthesis. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 148079. [Google Scholar] [CrossRef]
- Global Phycobilliuiprotein Market Research Report 2018 Global, China; Get Market Research: Munich, Germany, 2018.
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef] [PubMed]
- Matos, J.; Cardoso, C.L.; Falè, P.; Afonso, C.M.; Bandarra, N.M. Investigation of nutraceutical potential of the microalgae Chlorella vulgaris and Arthrospira platensis. Int. J. Food Sci. Technol. 2020, 55, 303–312. [Google Scholar] [CrossRef]
- Hsieh-Lo, M.; Castillo, G.; Ochoa-Becerra, M.A.; Mojica, L. Phycocyanin and phycoerythrin: Strategies to improve production yield and chemical stability. Algal Res. 2019, 42, 101600. [Google Scholar] [CrossRef]
- Babu, T.S.; Kumar, A.; Varma, A.K. Effect of light quality on phycobilisome components of the cyanobacterium Spirulina platensis. Plant Physiol. 1991, 95, 492–497. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, C.Y.; Fu, C.C.; Liu, Y.C. Effects of using light-emitting diodes on the cultivation of Spirulina platensis. Biochem. Eng. J. 2007, 37, 21–25. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, J.E.; Kim, Y.; Lee, S.Y. The production of high purity phycocyanin by Spirulina platensis using light-emitting diodes based two-stage cultivation. Appl. Biochem. Biotechnol. 2016, 178, 382–395. [Google Scholar] [CrossRef]
- Lima, G.M.; Teixeira, P.C.N.; Teixeira, C.M.L.L.; Filócomo, D.; Lage, C.L.S. Influence of spectral light quality on the pigment concentrations and biomass productivity of Arthrospira platensis. Algal Res. 2018, 31, 157–166. [Google Scholar] [CrossRef]
- Zarrouk, C. Contribution a l’etude d’une cyanophycee. Influence de divers facteurs physiques et chimiques sur la croissance et la photosynthese de Spirulina maxima (Setch. et Gardner) Geitler. Ph.D. Thesis, University of Paris, Paris, France, 1966. [Google Scholar]
- Chini Zittelli, G.; Mugnai, G.; Milia, M.; Cicchi, B.; Silva Benavides, A.M.; Angioni, A.; Addis, P.; Torzillo, G. Effects of blue, orange and white lights on growth, chlorophyll fluorescence, and phycocyanin production of Arthrospira platensis cultures. Algal Res. 2022, 61, 102583. [Google Scholar] [CrossRef]
- Singh, M.; Singh, M.P. Chlorophyll content and its relation to productivity of crops and weeds in a tropical agroecosystem at Gujar Lake Margin, India. Ecoprint 2016, 23, 39–44. [Google Scholar] [CrossRef]
- Chamizo, S.; Adessi, A.; Torzillo, G.; De Philippis, R. Exopolysaccharide Features Influence Growth Success in Biocrust-forming Cyanobacteria, Moving from Liquid Culture to Sand Microcosms. Front. Microbiol. 2020, 11, 2719. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Chen, Y.; Vaidyanathan, S. A simple, reproducible and sensitive spectrophotometric method to estimate microalgal lipids. Anal. Chim. Acta 2012, 724, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Siegelman, H.W.; Kycia, J.H. Biliproteins d’alghe. In Manuale di Metodi USURY; Hellebust, J.A., Craigie, J.S., Eds.; Cambridge University Press: Cambridge, UK, 1978; pp. 72–78. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Aguilera, J.; Niell, F.X. Red and blue light regulation of growth and photosynthetic metabolism in Porphyra umbilicalis (Bangiales, Rhodophyta). Eur. J. Phycol. 1995, 30, 11–18. [Google Scholar] [CrossRef]
- Chen, H.B.; Wu, J.Y.; Wang, C.F.; Fu, C.C.; Shieh, C.J.; Chen, C.I.; Wang, C.Y.; Liu, Y.C. Modelling on chlorophyll a and phycocyanin production by Spirulina platensis under various light-emitting diodes. Biochem. Eng. J. 2010, 53, 52–56. [Google Scholar] [CrossRef]
- Wicaksono, H.A.; Satyantini, W.H.; Masithah, E.D. The spectrum of light and nutrients required to increase the production of phycocyanin Spirulina platensis. IOP Conf. Ser. Earth Environ. Sci. 2019, 236. [Google Scholar] [CrossRef]
- Roessler, P.G. Environmental control of glycerolipid metabolism in microalgae: Commercial implications and future research directions. J. Phycol. 1990, 26, 393–399. [Google Scholar] [CrossRef]
- Kurat, C.F.; Natter, K.; Petschnigg, J.; Wolinski, H.; Scheuringer, K.; Scholz, H.; Zimmermann, R.; Leber, R.; Zechner, R.; Kohlwein, S.D. Obese yeast: Triglyceride lipolysis is functionally conserved from mammals to yeast. J. Biol. Chem. 2006, 281, 491–500. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Dhar, D. Microalgae as second generation biofuel. A review. Agron. Sustain. Dev. 2011, 31, 605–629. [Google Scholar] [CrossRef]
- Zaparoli, M.; Ziemniczak, F.; Mantovani, L.; Costa, J.A.; Colla, L. Cellular Stress Conditions as a Strategy to Increase Carbohydrate Productivity in Spirulina platensis. BioEnergy Res. 2020, 13, 1221–1234. [Google Scholar] [CrossRef]
- Vieira Salla, A.C.; Margarites, A.C.; Seibel, F.I.; Holz, L.C.; Barbosa Brião, V.; Bertolin, T.E.; Colla, L.M.; Vieira Costa, J.A. Increase in the carbohydrate content of the microalgae Spirulina in culture by nutrient starvation and the addition of residues of whey protein concentrate. Bioresour. Technol. 2016, 209, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Torzillo, G.; Sacchi, A.; Materassi, R.; Richmond, A. Effect of temperature on yield and night biomass loss in Spirulina platensis grown outdoors in tubular photobioreactors. J. Appl. Phycol. 1991, 3, 103–109. [Google Scholar] [CrossRef]
- Anvar, A.A.; Nowruzi, B. Bioactive properties of spirulina: A review. Microb. Bioact. 2021, 134–142. [Google Scholar] [CrossRef]
- Saharan, V.; Jood, S. Nutritional composition of Spirulina platensis and its acceptability in food products. Int. J. Adv. Res. (IJAR) 2017, 5, 2295–2300. [Google Scholar] [CrossRef]
- Vargas, M.A.; Rodriguez, H.; Moreno, J.; Olivares, H.; Del Campo, J.A.; Rivas, J.; Guerrero, M.G. Biochemical composition and fatty acid content of filamentous nitrogen-fixing cyanobacteria. J. Phycol. 1998, 34, 812–817. [Google Scholar] [CrossRef]
- Park, J.; Dinh, T.B. Contrasting effects of monochromatic LED lighting on growth, pigments and photosynthesis in the commercially important cyanobacterium Arthrospira maxima. Bioresour. Technol. 2019, 291, 121846. [Google Scholar] [CrossRef]
- Masojídek, J.; Koblížek, M.; Torzillo, G. Photosynthesis in microalgae. In Handbook of Microalgal Mass Cultures; Richmond, A., Ed.; Blackwell Science: Oxford, UK, 2004; pp. 20–39. [Google Scholar]
- de Mooij, T.; Janssen, M.; Cerezo-Chinarro, O.; Mussgnug, J.H.; Kruse, O.; Ballottari, M.; Bassi, R.; Bujaldon, S.; Wollman, F.A.; Wijffels, R.H. Antenna size reduction as a strategy to increase biomass productivity: A great potential not yet realized. J. Appl. Phycol. 2015, 27, 1063–1077. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light | M2 | M2M | A. maxima |
|---|---|---|---|
| White | 7.07 a(a) * | 6.11 a(b) | 9.36 a(c) |
| Orange | 5.01 b(a) | 8.25 b(b) | 6.21 b(c) |
| Blue | 5.23 b(a) | 7.32 b(b) | 3.38 c(c) |
| Light | M2 | M2M | A. maxima |
|---|---|---|---|
| White | 71.18 a(a) * | 63.71 a(b) | 72.14 a(a) |
| Orange | 60.81 b(a) | 39.09 b(b) | 50.63 b(c) |
| Blue | 74.88 a(a) | 85.89 c(b) | 73.53 a(a) |
| Light | M2 | M2M | A. maxima |
|---|---|---|---|
| White | 9.81 a(a) * | 17.75 a(b) | 42.36 a(c) |
| Orange | 16.41 b(a) | 30.17 b(c) | 22.18 b(b) |
| Blue | 13.75 c(a) | 13.33 c(a) | 21.35 b(b) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milia, M.; Corrias, F.; Addis, P.; Chini Zitelli, G.; Cicchi, B.; Torzillo, G.; Andreotti, V.; Angioni, A. Influence of Different Light Sources on the Biochemical Composition of Arthrospira spp. Grown in Model Systems. Foods 2022, 11, 399. https://doi.org/10.3390/foods11030399
Milia M, Corrias F, Addis P, Chini Zitelli G, Cicchi B, Torzillo G, Andreotti V, Angioni A. Influence of Different Light Sources on the Biochemical Composition of Arthrospira spp. Grown in Model Systems. Foods. 2022; 11(3):399. https://doi.org/10.3390/foods11030399
Chicago/Turabian StyleMilia, Massimo, Francesco Corrias, Piero Addis, Graziella Chini Zitelli, Bernardo Cicchi, Giuseppe Torzillo, Valeria Andreotti, and Alberto Angioni. 2022. "Influence of Different Light Sources on the Biochemical Composition of Arthrospira spp. Grown in Model Systems" Foods 11, no. 3: 399. https://doi.org/10.3390/foods11030399
APA StyleMilia, M., Corrias, F., Addis, P., Chini Zitelli, G., Cicchi, B., Torzillo, G., Andreotti, V., & Angioni, A. (2022). Influence of Different Light Sources on the Biochemical Composition of Arthrospira spp. Grown in Model Systems. Foods, 11(3), 399. https://doi.org/10.3390/foods11030399