

Hypoglycemic Effects and Mechanisms of Buckwheat–Oat–Pea Composite Flour in Diabetic Rats

Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Animal Diets
2.2. Animal and Housing Environment
2.3. Establishment of the Diabetic Rat Model and Experimental Treatments
2.4. Oral Glucose Tolerance Test
2.5. Serum Biochemical Analysis
2.6. Histological Analysis
2.7. Gut Microbiota Analysis with 16S rDNA Gene
2.8. Statistical Analysis
3. Results
3.1. Establishment of the Diabetic Model and Hypoglycemic Effects of BOP
3.2. Effects of BOP on Blood Lipids
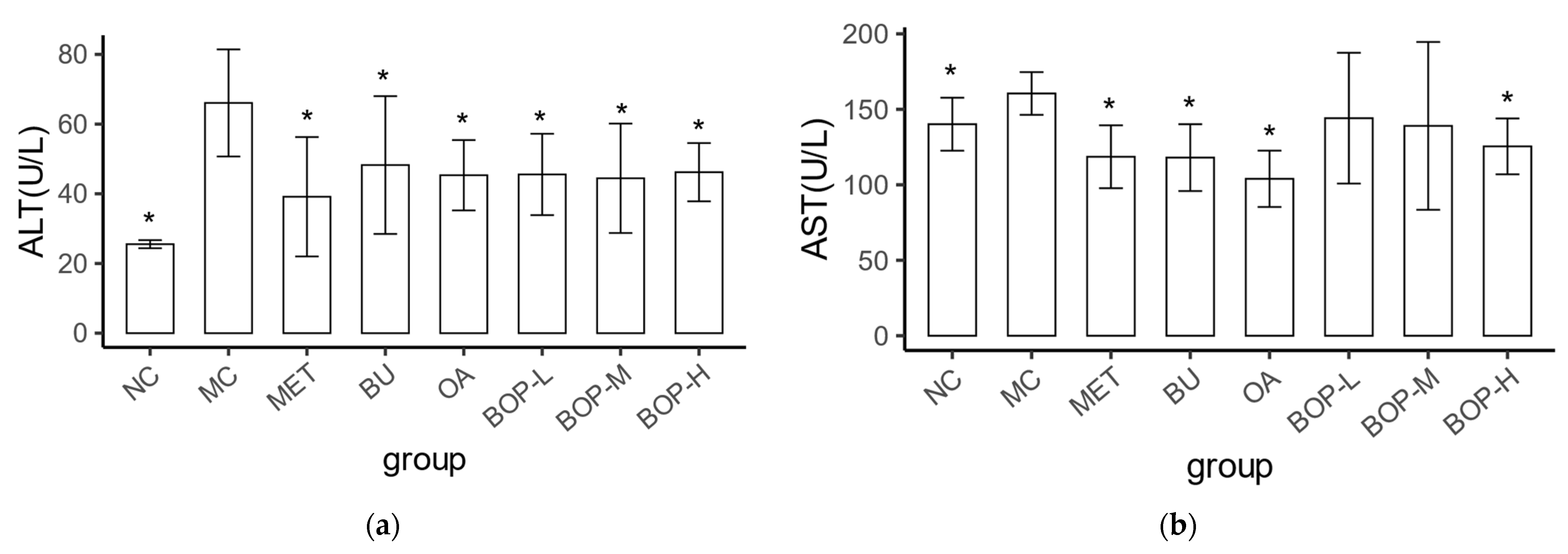
3.3. Effects of BOP on Liver Injury
3.4. Effects of BOP on Gut Microbiota
3.4.1. Alpha and Beta Diversity Analysis
3.4.2. Composition of Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Diabetes. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 3 February 2022).
- International Diabetes Federation. IDF Atlas-10th Edition. Available online: https://diabetesatlas.org/ (accessed on 10 January 2022).
- American Diabetes Association Professional Practice Committee. Facilitating Behavior Change and Well-Being to Improve Health Outcomes: Standards of Medical Care in Diabetes-2022. Diabetes Care 2021, 45, S60–S82. [Google Scholar] [CrossRef]
- Stringer, D.M.; Taylor, C.G.; Appah, P.; Blewett, H.; Zahradka, P. Consumption of Buckwheat Modulates the Post-Prandial Response of Selected Gastrointestinal Satiety Hormones in Individuals with Type 2 Diabetes Mellitus. Metab. Clin. Exp. 2013, 62, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-C.; Lee, B.-H.; Lai, Y.-J. Antioxidation and Antiglycation of Fagopyrum Tataricum Ethanol Extract. J. Food Sci. Technol. 2015, 52, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Gimenez-Bastida, J.A.; Zielinski, H.; Piskula, M.; Zielinska, D.; Szawara-Nowak, D. Buckwheat Bioactive Compounds, Their Derived Phenolic Metabolites and Theirhealth Benefits. Mol. Nutr. Food Res. 2017, 61, 1600475. [Google Scholar] [CrossRef]
- He, L.X.; Zhao, J.; Huang, Y.S.; Li, Y. The Difference between Oats and Beta-Glucan Extract Intake in the Management of HbA1c, Fasting Glucose and Insulin Sensitivity: A Meta-Analysis of Randomized Controlled Trials. Food Funct. 2016, 7, 1413–1428. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, Y.; Zhang, H.; Hu, B.; Wang, L.; Qian, H.; Qi, X. The Anti-Diabetic Activity of Oat β-d-Glucan in Streptozotocin-Nicotinamide Induced Diabetic Mice. Int. J. Biol. Macromol. 2016, 91, 1170–1176. [Google Scholar] [CrossRef] [PubMed]
- Paudel, D.; Dhungana, B.; Caffe, M.; Krishnan, P. A Review of Health-Beneficial Properties of Oats. Foods 2021, 10, 2591. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, Z.; Fouhse, J.; Im, H.S.; Chan, C.B.; Willing, B.P. Dietary Pea Fiber Supplementation Improves Glycemia and Induces Changes in TheComposition of Gut Microbiota, Serum Short Chain Fatty Acid Profile and Expression of Mucins in Glucose Intolerant Rats. Nutrients 2017, 9, 1236. [Google Scholar] [CrossRef] [PubMed]
- Beitane, I.; Krumina-Zemture, G.; Sabovics, M. Technological Properties of Pea and Buckwheat Flours and Their Blends. In Research for Rural Development 2015; Volume 1, Treija, S., Skujeniece, S., Eds.; Latvia Univ Life Sciences & Technologies: Jelgava, Latvia, 2016; pp. 137–142. [Google Scholar]
- Angioloni, A.; Collar, C. Nutritional and Functional Added Value of Oat, Kamut, Spelt, Rye and Buckwheat versus Common Wheat in Breadmaking. J. Sci. Food Agric. 2011, 91, 1283–1292. [Google Scholar] [CrossRef]
- Yin, X.Q.; Zhang, X.X.; Wen, J.; Liu, S.Q.; Liu, X.R.; Zhou, R.Y.; Wang, J.B. Effects of the composite of buckwheat-oat-pea on blood glucose in diabetic rats. Beijing Da Xue Xue Bao Yi Xue Ban 2021, 53, 447–452. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Mouse Tissue Fixation. Cold Spring Harb. Protoc. 2014, 2014, pdb-prot073403. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and Validation of a Histological Scoring System for Nonalcoholic Fatty Liver Disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Micha, R.; Khatibzadeh, S.; Shi, P.; Andrews, K.G.; Engell, R.E.; Mozaffarian, D. Global Burden of Diseases Nutrition and Chronic Diseases Expert Group (NutriCoDE) Global, Regional and National Consumption of Major Food Groups in 1990 and 2010: A Systematic Analysis Including 266 Country-Specific Nutrition Surveys Worldwide. BMJ Open 2015, 5, e008705. [Google Scholar] [CrossRef]
- Ruggiero, E.; Bonaccio, M.; Di Castelnuovo, A.; Bonanni, A.; Costanzo, S.; Persichillo, M.; Bracone, F.; Cerletti, C.; Donati, M.B.; de Gaetano, G.; et al. Consumption of Whole Grain Food and Its Determinants in a General Italian Population: Results from the INHES Study. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lietz, G.; Seal, C. Buckwheat and CVD Risk Markers: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 619. [Google Scholar] [CrossRef]
- Ren, G.; Fan, X.; Teng, C.; Li, Y.; Everaert, N.; Blecker, C. The Beneficial Effect of Coarse Cereals on Chronic Diseases through Regulating Gut Microbiota. Foods 2021, 10, 2891. [Google Scholar] [CrossRef] [PubMed]
- Perez-Ramirez, I.F.; Becerril-Ocampo, L.J.; Reynoso-Camacho, R.; Herrera, M.D.; Guzman-Maldonado, S.H.; Cruz-Bravo, R.K. Cookies Elaborated with Oat and Common Bean Flours Improved Serum Markers in Diabetic Rats. J. Sci. Food Agric. 2018, 98, 998–1007. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.T.; Ha, J.; Onuzuruike, A.U.; Kasturi, K.; Galvan-De La Cruz, M.; Bingham, B.A.; Baker, R.L.; Utumatwishima, J.N.; Mabundo, L.S.; Ricks, M.; et al. Time to Glucose Peak during an Oral Glucose Tolerance Test Identifies Prediabetes Risk. Clin. Endocrinol. 2017, 87, 484–491. [Google Scholar] [CrossRef]
- Guo, X.-X.; Wang, Y.; Wang, K.; Ji, B.-P.; Zhou, F. Stability of a Type 2 Diabetes Rat Model Induced by High-Fat Diet Feeding Withlow-Dose Streptozotocin Injection. J. Zhejiang Univ. Sci. B 2018, 19, 559–569. [Google Scholar] [CrossRef]
- Gheibi, S.; Kashfi, K.; Ghasemi, A. A Practical Guide for Induction of Type-2 Diabetes in Rat: Incorporating Ahigh-Fat Diet and Streptozotocin. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 95, 605–613. [Google Scholar] [CrossRef] [PubMed]
- De Silva, N.M.G.; Borges, M.C.; Hingorani, A.D.; Engmann, J.; Shah, T.; Zhang, X.; Luan, J.; Langenberg, C.; Wong, A.; Kuh, D.; et al. Liver Function and Risk of Type 2 Diabetes: Bidirectional Mendelian Randomization Study. Diabetes 2019, 68, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Bäckhed, F. Functional Interactions between the Gut Microbiota and Host Metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Hills, R.D.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, E1613. [Google Scholar] [CrossRef] [PubMed]
- Ballan, R.; Saad, S.M.I. Characteristics of the Gut Microbiota and Potential Effects of Probiotic Supplements in Individuals with Type 2 Diabetes Mellitus. Foods 2021, 10, 2528. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-C.; Fan, Z.-Y.; Wang, H.-Y.; Wen, D.-C.; Zhang, S.-Y. Effect of Polysaccharides from Adlay Seed on Anti-Diabetic and Gut Microbiota. Food Funct. 2019, 10, 4372–4380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human Gut Microbiota Changes Reveal the Progression of Glucose Intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef]
- Ma, D.; Wang, A.C.; Parikh, I.; Green, S.J.; Hoffman, J.D.; Chlipala, G.; Murphy, M.P.; Sokola, B.S.; Bauer, B.; Hartz, A.M.S.; et al. Ketogenic Diet Enhances Neurovascular Function with Altered Gut Microbiome in Young Healthy Mice. Sci. Rep. 2018, 8, 6670. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Diet | HFD | Buckwheat Diet | Oats Diet | Low Dose BOP Diet | Medium Dose BOP Diet | High Dose BOP Diet | |
|---|---|---|---|---|---|---|---|
| BOP | 0.0 | 0.0 | 0.0 | 0.0 | 33.0 | 100.0 | 300.0 |
| Buckwheat flour | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Oats flour | 0.0 | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 |
| Lactic casein | 140.0 | 234.9 | 220.1 | 218.3 | 229.5 | 218.5 | 185.4 |
| L-Cystine | 1.8 | 2.1 | 2.1 | 2.1 | 2.1 | 2.1 | 2.1 |
| Corn starch | 465.7 | 85.5 | 12.0 | 18.9 | 62.0 | 14.3 | 0.0 |
| Maltodextrin | 155.0 | 117.5 | 117.5 | 117.5 | 117.5 | 117.5 | 0.0 |
| Sucrose | 100.0 | 207.7 | 207.6 | 207.7 | 207.7 | 207.7 | 197.1 |
| Cellulose | 50.0 | 58.7 | 51.1 | 51.4 | 56.0 | 50.7 | 34.9 |
| Soybean Oil | 40.0 | 29.4 | 29.4 | 29.4 | 29.4 | 29.4 | 29.4 |
| Lard | 0.0 | 208.5 | 204.5 | 199.0 | 207.1 | 204.1 | 195.4 |
| Mineral Mix | 35.0 | 41.1 | 41.1 | 41.1 | 41.1 | 41.1 | 41.1 |
| Choline Bitartrate | 2.5 | 2.9 | 2.9 | 2.9 | 2.9 | 2.9 | 2.9 |
| Vitamin Mix | 10.0 | 11.7 | 11.7 | 11.7 | 11.7 | 11.7 | 11.7 |
| Tert-butylhydroquinone | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| Overall carbohydrate | 770.7 | 469.4 | 468.7 | 468.7 | 468.8 | 468.2 | 466 |
| Overall protein | 141.8 | 237.0 | 236.2 | 236.4 | 236.9 | 236.6 | 235.5 |
| Overall fat | 40.0 | 237.9 | 237.2 | 237.5 | 237.7 | 237.3 | 236.2 |
| Groups | TC Median (25% IQR, 75% IQR) | TG Median (25% IQR, 75% IQR) | HDL-C Mean ± SD | LDL-C Median (25% IQR, 75% IQR) |
|---|---|---|---|---|
| NC | 1.22 (1.12, 1.39) * | 0.82 (0.60, 0.95) * | 0.43 ± 0.05 * | 0.27 (0.24, 0.28) * |
| MC | 1.70 (1.43, 1.94) | 1.68 (1.21, 2.35) | 0.86 ± 0.17 | 0.32 (0.30, 0.46) |
| MET | 1.41 (1.32, 1.46) | 0.40 (0.36, 0.54) * | 0.60 ± 0.06 * | 0.28 (0.27, 0.32) |
| BU | 1.17 (1.14, 1.49) * | 0.26 (0.22, 1.29) * | 0.65 ± 0.14 * | 0.23 (0.18, 0.26) |
| OA | 1.54 (1.44, 1.73) | 0.70 (0.37, 1.14) | 0.67 ± 0.15 | 0.33 (0.30, 0.47) |
| BOP-L | 1.29 (1.15, 1.51) * | 0.77 (0.51, 0.99) | 0.65 ± 0.21 * | 0.31 (0.26, 0.33) |
| BOP-M | 1.73 (1.62, 1.82) | 0.78 (0.48, 1.67) | 0.75 ± 0.19 | 0.37 (0.31, 0.53) |
| BOP-H | 1.38 (1.34, 1.51) | 0.74 (0.64, 1.02) | 0.68 ± 0.09 | 0.24 (0.23, 0.27) |
| Group | Chao1 Median (25% IQR, 75% IQR) | Shannon Median (25% IQR, 75% IQR) | Faith’s Phylogenetics Diversity Median (25% IQR, 75% IQR) | Simpson Median (25% IQR, 75% IQR) |
|---|---|---|---|---|
| NC | 255.0 (241.0, 270.0) | 5.22 (4.85, 5.74) | 17.87 (17.49, 18.07) | 0.94 (0.91, 0.95) |
| MC | 237.0 (176.0, 347.0) | 5.68 (4.79, 6.34) | 18.03 (12.02, 21.88) | 0.96 (0.88, 0.96) |
| MET | 187.0 (134.2, 292.0) | 3.91 (3.07, 4.94) | 11.46 (11.43, 13.52) | 0.83 (0.74, 0.90) |
| BU | 313.0 (217.0, 334.0) | 4.83 (4.49, 4.96) | 17.63 (11.76, 21.43) | 0.92 (0.89, 0.93) |
| OA | 165.0 (123.0, 225.0) | 3.53 (3.47, 4.48) | 13.51 (10.62, 18.27) | 0.83 (0.72, 0.84) |
| BOP-L | 147.0 (146.0, 153.0) | 4.21 (3.97, 4.70) | 11.93 (11.10, 12.22) | 0.87 (0.85, 0.92) |
| BOP-M | 184.0 (182.0, 204.1) | 4.82 (4.37, 5.37) | 14.51 (14.13, 16.23) | 0.93 (0.90, 0.95) |
| BOP-H | 288.2 (186.3, 453.0) | 5.62 (5.05, 5.84) | 14.07 (13.02, 18.57) | 0.95 (0.94, 0.96) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, X.; Liu, S.; Zhang, X.; Jian, Y.; Wen, J.; Zhou, R.; Yin, N.; Liu, X.; Hou, C.; Wang, J. Hypoglycemic Effects and Mechanisms of Buckwheat–Oat–Pea Composite Flour in Diabetic Rats. Foods 2022, 11, 3938. https://doi.org/10.3390/foods11233938
Yin X, Liu S, Zhang X, Jian Y, Wen J, Zhou R, Yin N, Liu X, Hou C, Wang J. Hypoglycemic Effects and Mechanisms of Buckwheat–Oat–Pea Composite Flour in Diabetic Rats. Foods. 2022; 11(23):3938. https://doi.org/10.3390/foods11233938
Chicago/Turabian StyleYin, Xueqian, Siqi Liu, Xiaoxuan Zhang, Yuanzhi Jian, Jing Wen, Ruoyu Zhou, Ning Yin, Xinran Liu, Chao Hou, and Junbo Wang. 2022. "Hypoglycemic Effects and Mechanisms of Buckwheat–Oat–Pea Composite Flour in Diabetic Rats" Foods 11, no. 23: 3938. https://doi.org/10.3390/foods11233938
APA StyleYin, X., Liu, S., Zhang, X., Jian, Y., Wen, J., Zhou, R., Yin, N., Liu, X., Hou, C., & Wang, J. (2022). Hypoglycemic Effects and Mechanisms of Buckwheat–Oat–Pea Composite Flour in Diabetic Rats. Foods, 11(23), 3938. https://doi.org/10.3390/foods11233938