
Screening and Identification of Novel Soluble Epoxide Hydrolase Inhibitors from Corn Gluten Peptides

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of Corn Gluten Peptides
2.3. Virtual Screening of Potential sEH Inhibitory Peptides
2.4. Validation of the Reaction Conditions between the sEH and Substrate
2.5. Determination of the sEH Inhibitory Activities of Peptides
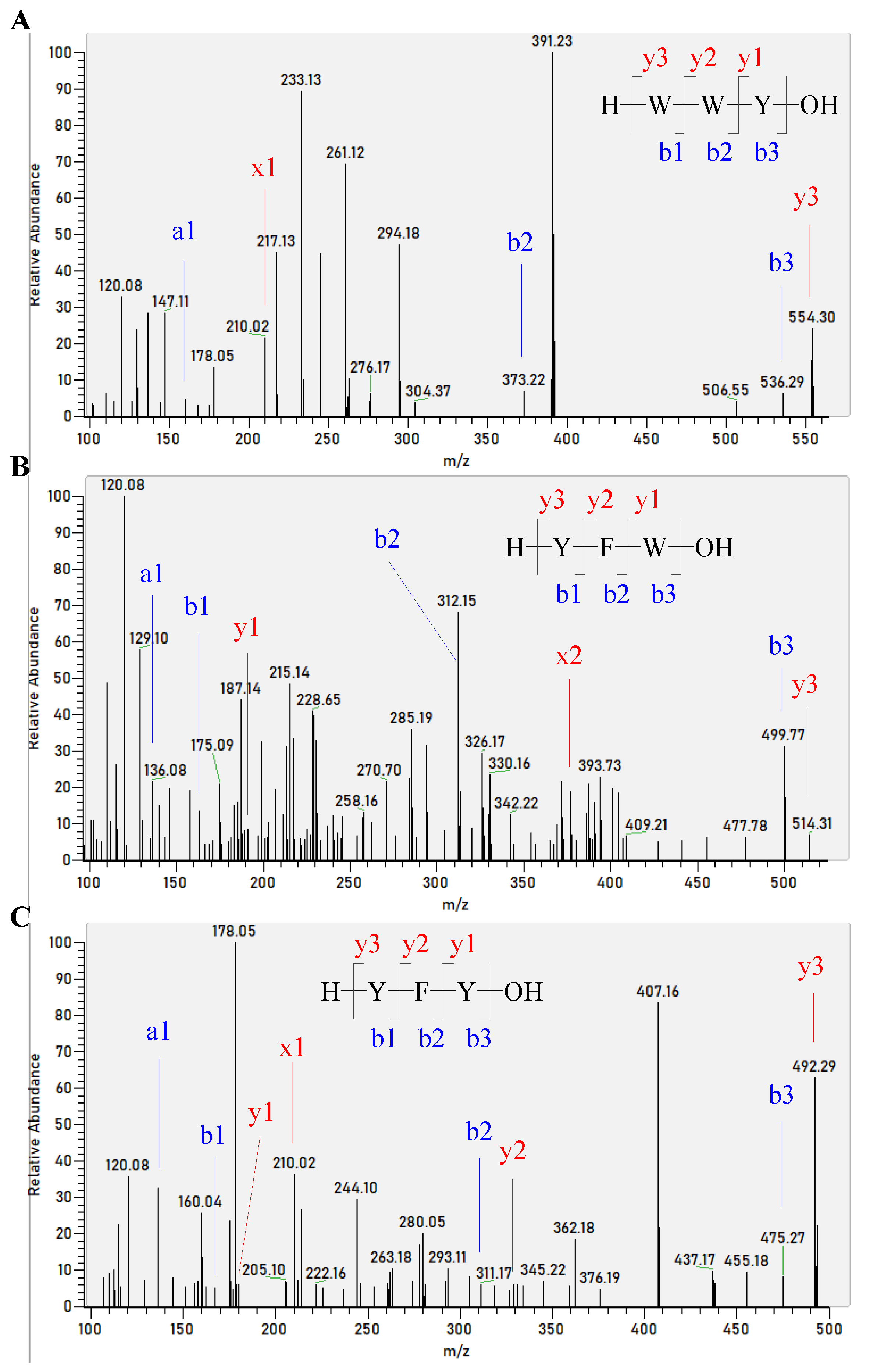
2.6. Identification of Corn Gluten Peptides Using Nano LC-MS/MS
2.7. Physicochemical Property and Toxicity Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. The Optimal Reaction Conditions between the sEH and Substrate
3.2. Virtual Screening of sEH Inhibitory Peptides by Molecular Docking
3.3. Inhibition Mode of sEH Inhibitory Peptides
3.4. Interactions between Peptides and sEH Enzyme
3.5. Physicochemical Property and Toxicity Analysis of sEH Inhibitory Peptides
3.6. The sEH Inhibitory Activities of Corn Gluten Peptides
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Harris, T.R.; Hammock, B.D. Soluble epoxide hydrolase: Gene structure, expression and deletion. Gene 2013, 526, 61–74. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wang, C.; Zhu, Y.; Ai, D. Soluble epoxide hydrolase: A potential target for metabolic diseases. J. Diabetes 2016, 8, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Morisseau, C.; Hammock, B.D. Impact of soluble epoxide hydrolase and epoxyeicosanoids on human health. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Swardfager, W.; Hennebelle, M.; Yu, D.; Hammock, B.D.; Levitt, A.J.; Hashimoto, K.; Taha, A.Y. Metabolic/inflammatory/vascular comorbidity in psychiatric disorders; soluble epoxide hydrolase (sEH) as a possible new target. Neurosci. Biobehav. Rev. 2018, 87, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Waltenberger, B.; Garscha, U.; Temml, V.; Liers, J.; Werz, O.; Schuster, D.; Stuppner, H. Discovery of potent soluble epoxide hydrolase (sEH) inhibitors by pharmacophore-based virtual screening. J. Chem. Inf. Model. 2016, 56, 747–762. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.C.; Hammock, B.D. Discovery of inhibitors of soluble epoxide hydrolase: A target with multiple potential therapeutic indications. J. Med. Chem. 2012, 55, 1789–1808. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.P.; Zhang, X.Y.; Morisseau, C.; Hwang, S.H.; Zhang, Z.J.; Hammock, B.D.; Ma, X.C. Discovery of soluble epoxide hydrolase inhibitors from chemical synthesis and natural products. J. Med. Chem. 2021, 64, 184–215. [Google Scholar] [CrossRef]
- Zhao, W.Y.; Zhang, X.Y.; Zhou, M.R.; Tian, X.G.; Lv, X.; Zhang, H.L.; Deng, S.; Zhang, B.J.; Sun, C.P.; Ma, X.C. Natural soluble epoxide hydrolase inhibitors from Alisma orientale and their potential mechanism with soluble epoxide hydrolase. Int. J. Biol. Macromol. 2021, 183, 811–817. [Google Scholar] [CrossRef]
- Liu, Z.B.; Sun, C.P.; Xu, J.X.; Morisseau, C.; Hammock, B.D.; Qiu, F. Phytochemical constituents from Scutellaria baicalensis in soluble epoxide hydrolase inhibition: Kinetics and interaction mechanism merged with simulations. Int. J. Biol. Macromol. 2019, 133, 1187–1193. [Google Scholar] [CrossRef]
- Leem, H.H.; Lee, G.Y.; Lee, J.S.; Lee, H.; Kim, J.H.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of components from Leonurus japonicus. Int. J. Biol. Macromol. 2017, 103, 451–457. [Google Scholar] [CrossRef]
- Bai, M.M.; Shi, W.; Tian, J.M.; Lei, M.; Kim, J.H.; Sun, Y.N.; Kim, Y.H.; Gao, J.M. Soluble epoxide hydrolase inhibitory and anti-inflammatory components from the leaves of Eucommia ulmoides Oliver (duzhong). J. Agric. Food Chem. 2015, 63, 2198–2205. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.S.; Ng, J.C.; Yang, J.; Hwang, S.H.; Morisseau, C.; Wagner, K.; Hammock, B.D. Preparation and evaluation of soluble epoxide hydrolase inhibitors with improved physical properties and potencies for treating diabetic neuropathic pain. Bioorganic Med. Chem. 2020, 28, 115735. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, M.; Singh, J.; Gani, A. Exploration of bioactive peptides from various origin as promising nutraceutical treasures: In vitro, in silico and in vivo studies. Food Chem. 2022, 373, 131395. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; He, H.; Hou, T. A comprehensive review of corn protein-derived bioactive peptides: Production, characterization, bioactivities, and transport pathways. Compr. Rev. Food Sci. Food Saf. 2019, 18, 329–345. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Wang, Y.; Zhang, Y.; Liu, J. Isolation and characterisation of in vitro and cellular free radical scavenging peptides from corn peptide fractions. Molecules 2015, 20, 3221–3237. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Yu, Z.; Zhang, T.; Ma, S.; Liu, J. Intracellular ROS scavenging and antioxidant enzyme regulating capacities of corn gluten meal-derived antioxidant peptides in HepG2 cells. Food Res. Int. 2016, 90, 33–41. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Xue, C.; Ma, S.; Du, Z.; Zhang, T.; Liu, J. Corn gluten hydrolysate regulates the expressions of antioxidant defense and ROS metabolism relevant genes in H2O2-induced HepG2 cells. J. Funct. Foods 2018, 42, 362–370. [Google Scholar] [CrossRef]
- Zhao, W.Z.; Li, X.; Yu, Z.P.; Wu, S.J.; Ding, L.; Liu, J.B. Identification of lactoferrin-derived peptides as potential inhibitors against the main protease of SARS-CoV-2. Leb. Wiss Technol. 2022, 154, 112684. [Google Scholar] [CrossRef]
- Yu, Z.P.; Cao, Y.X.; Kan, R.T.; Ji, H.Z.; Zhao, W.Z.; Wu, S.J.; Liu, J.B.; Shiuan, D. Identification of egg protein-derived peptides as xanthine oxidase inhibitors: Virtual hydrolysis, molecular docking, and in vitro activity evaluation. Food Sci. Hum. Wellness 2022, 6, 1591–1597. [Google Scholar] [CrossRef]
- You, H.; Wu, T.; Wang, W.; Li, Y.; Liu, X.; Ding, L. Preparation and identification of dipeptidyl peptidase IV inhibitory peptides from quinoa protein. Food Res. Int. 2022, 156, 111176. [Google Scholar] [CrossRef]
- Gomez, G.A.; Morisseau, C.; Hammock, B.D.; Christianson, D.W. Human soluble epoxide hydrolase: Structural basis of inhibition by 4-(3-cyclohexylureido)-carboxylic acids. Protein Sci. 2006, 15, 58–64. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. The PyMOL Molecular Graphics System. 2002. Available online: http://www.pymol.org (accessed on 10 October 2022).
- Li, H.X.; Heo, M.; Go, Y.; Kim, Y.S.; Kim, Y.H.; Yang, S.Y.; Li, W. Coumarin and moracin derivatives from mulberry leaves (Morus alba L.) with soluble epoxide hydrolase inhibitory activity. Molecules 2020, 25, 3967. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.D.; Wolf, N.M.; Morisseau, C.; Whetstone, P.; Hock, B.; Hammock, B.D. Fluorescent substrates for soluble epoxide hydrolase and application to inhibition studies. Anal. Biochem. 2005, 343, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Wolf, N.M.; Morisseau, C.; Jones, P.D.; Hock, B.; Hammock, B.D. Development of a high-throughput screen for soluble epoxide hydrolase inhibition. Anal. Biochem. 2006, 355, 71–80. [Google Scholar] [CrossRef]
- Cho, I.S.; Kim, J.H.; Lin, Y.; Su, X.D.; Kang, J.S.; Yang, S.Y.; Kim, Y.H. Inhibitory activity of quercetin 3-O-arabinofuranoside and 2-oxopomolic acid derived from Malus domestica on soluble epoxide hydrolase. Molecules 2020, 25, 4352. [Google Scholar] [CrossRef]
- Gazzillo, E.; Terracciano, S.; Ruggiero, D.; Potenza, M.; Chini, M.G.; Lauro, G.; Fischer, K.; Hofstetter, R.K.; Giordano, A.; Werz, O.; et al. Repositioning of quinazolinedione-based compounds on soluble epoxide hydrolase (sEH) through 3D structure-based pharmacophore model-driven investigation. Molecules 2022, 27, 3866. [Google Scholar] [CrossRef]
- Kitamura, S.; Morisseau, C.; Harris, T.R.; Inceoglu, B.; Hammock, B.D. Occurrence of urea-based soluble epoxide hydrolase inhibitors from the plants in the order Brassicales. PLoS ONE 2017, 12, e0176571. [Google Scholar] [CrossRef]
- Kim, J.H.; Jo, Y.D.; Jin, C.H. Isolation of soluble epoxide hydrolase inhibitor of capsaicin analogs from Capsicum chinense Jacq. cv. Habanero. Int. J. Biol. Macromol. 2019, 135, 1202–1207. [Google Scholar] [CrossRef]
- Fakhar, Z.; Hejazi, L.; Tabatabai, S.A.; Munro, O.Q. Discovery of novel heterocyclic amide-based inhibitors: An integrative in-silico approach to targeting soluble epoxide hydrolase. J. Biomol. Struct. Dyn. 2021, 40, 7114–7128. [Google Scholar] [CrossRef]
- Amano, Y.; Yamaguchi, T.; Tanabe, E. Structural insights into binding of inhibitors to soluble epoxide hydrolase gained by fragment screening and X-ray crystallography. Bioorganic Med. Chem. 2014, 22, 2427–2434. [Google Scholar] [CrossRef]
- Rezaee, E.; Amrolah, S.M.; Nazari, M.; Tabatabai, S.A. Novel amide derivatives of 3-phenylglutaric acid as potent soluble epoxide hydrolase inhibitors. Mol. Divers. 2021, 25, 45–53. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Sequence | Binding Free Energy (kcal/mol) | sEH Inhibitory Activity at 100 µM (%) |
|---|---|---|
| KII | −11.0883 | 37.99 ± 8.23 |
| WKW | −10.9831 | 31.13 ± 1.24 |
| WQW | −10.8840 | 37.19 ± 6.14 |
| WRW | −10.7671 | 38.30 ± 3.66 |
| WLR | −10.5700 | 43.82 ± 1.73 |
| YRW | −10.5607 | 47.58 ± 6.23 |
| RWI | −10.5393 | 19.63 ± 10.26 |
| FKW | −10.5095 | 36.29 ± 10.08 |
| YMW | −10.4981 | 49.96 ± 5.86 |
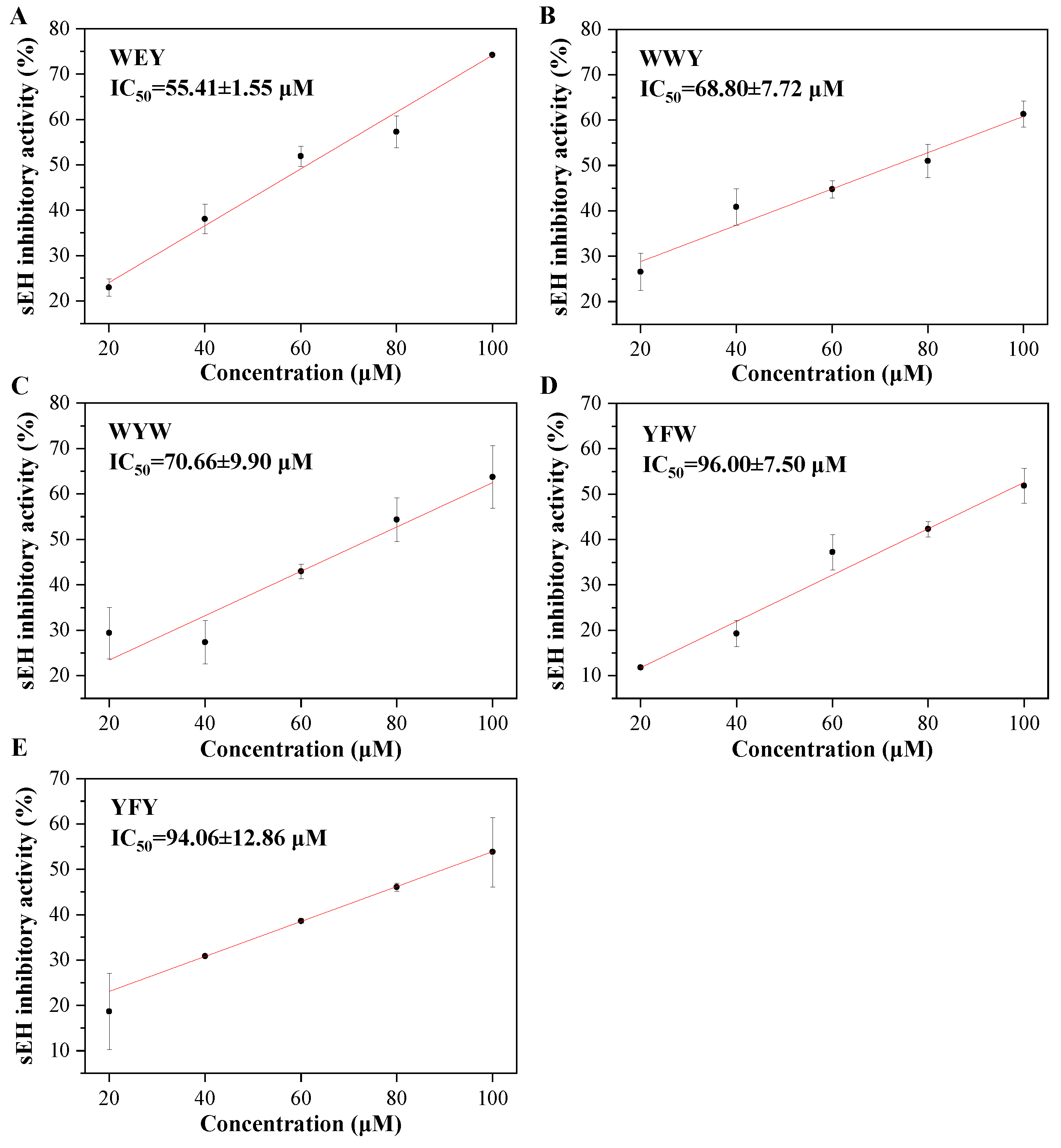
| WYW | −10.4858 | 60.30 ± 5.60 |
| FRY | −10.4562 | 45.83 ± 14.14 |
| WWY | −10.3874 | 55.15 ± 2.38 |
| YFW | −10.3840 | 57.38 ± 7.62 |
| YHW | −10.3641 | 32.06 ± 3.06 |
| YFY | −10.3629 | 52.96 ± 3.09 |
| WTR | −10.3607 | 26.05 ± 9.78 |
| RYW | −10.3486 | 35.89 ± 2.83 |
| YHY | −10.3050 | 41.43 ± 4.19 |
| WEY | −10.3024 | 70.01 ± 6.25 |
| WFW | −10.2877 | 36.24 ± 3.73 |
| WR | −8.4954 | 31.65 ± 4.20 |
| YW | −8.1424 | 32.08 ± 5.06 |
| RV | −8.1371 | 46.76 ± 5.18 |
| HR | −7.8854 | 29.71 ± 2.30 |
| WW | −7.8543 | 39.88 ± 3.12 |
| RW | −7.8351 | 19.84 ± 5.06 |
| YR | −7.8328 | 27.32 ± 5.18 |
| WF | −7.7965 | 43.98 ± 2.30 |
| WY | −7.7922 | 29.16 ± 2.35 |
| RY | −7.7852 | 22.75 ± 7.70 |
| Peptide Sequence | Interactions | Site from Peptide | Site from She | Distance (Å) | ||||
|---|---|---|---|---|---|---|---|---|
| WEY | hydrogen bond | Trp1 | NH3+ | H | Asp333 | CO | O | 2.0 |
| Tyr3 | OH | H | Arg408 | CO | O | 2.3 | ||
| arene–cation stacking | Trp1 | NH3+ | H | Trp334 | pyrrole ring | 3.4 | ||
| arene–arene stacking | Trp1 | pyrrole ring | Trp334 | pyrrole ring | 4.0 | |||
| arene–H stacking | Glu2 | CH | H | His523 | imidazole ring | 3.0 | ||
| WWY | hydrogen bond | Trp1 | NH3+ | H | Ser413 | OH | O | 2.2 |
| Tyr3 | NH | H | Met418 | S | S | 3.0 | ||
| arene–H stacking | Trp1 | benzene ring | His419 | CH | H | 2.8 | ||
| WYW | hydrogen bond | Trp1 | NH3+ | H | Asp333 | CO | O | 2.1 |
| YFW | hydrogen bond | Tyr1 | NH3+ | H | Asp333 | OH | O | 1.9 |
| arene–cation stacking | Tyr1 | NH3+ | H | Trp334 | pyrrole ring | 3.3 | ||
| arene–H stacking | Phe2 | CH | H | His523 | imidazole ring | 2.8 | ||
| YFY | hydrogen bond | Tyr1 | CH | H | Asp333 | OH | O | 2.1 |
| Phe2 | NH | H | Asp333 | OH | O | 2.2 | ||
| Tyr3 | OH | H | Lys494 | CO | O | 2.2 | ||
| Tyr3 | CO | O | Trp524 | NH | H | 2.5 | ||
| arene–arene stacking | Trp1 | benzene ring | Trp334 | pyrrole ring | 3.6 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, J.; Du, S.; Wang, L. Screening and Identification of Novel Soluble Epoxide Hydrolase Inhibitors from Corn Gluten Peptides. Foods 2022, 11, 3695. https://doi.org/10.3390/foods11223695
Dang J, Du S, Wang L. Screening and Identification of Novel Soluble Epoxide Hydrolase Inhibitors from Corn Gluten Peptides. Foods. 2022; 11(22):3695. https://doi.org/10.3390/foods11223695
Chicago/Turabian StyleDang, Jiamin, Shuangkui Du, and Liying Wang. 2022. "Screening and Identification of Novel Soluble Epoxide Hydrolase Inhibitors from Corn Gluten Peptides" Foods 11, no. 22: 3695. https://doi.org/10.3390/foods11223695
APA StyleDang, J., Du, S., & Wang, L. (2022). Screening and Identification of Novel Soluble Epoxide Hydrolase Inhibitors from Corn Gluten Peptides. Foods, 11(22), 3695. https://doi.org/10.3390/foods11223695