
Wasabi (Eutrema japonicum) Reduces Obesity and Blood Pressure in Diet-Induced Metabolic Syndrome in Rats
 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Composition of Wasabi Powder (Stem and Rhizome)
2.3. Rats and Diets
2.4. Body Composition
2.5. Oral Glucose Tolerance and Systolic Blood Pressure Measurement
2.6. Plasma Biomarkers, Tissue Weights and Histology of Liver and Heart
2.7. Statistical Analysis
3. Results and Discussion
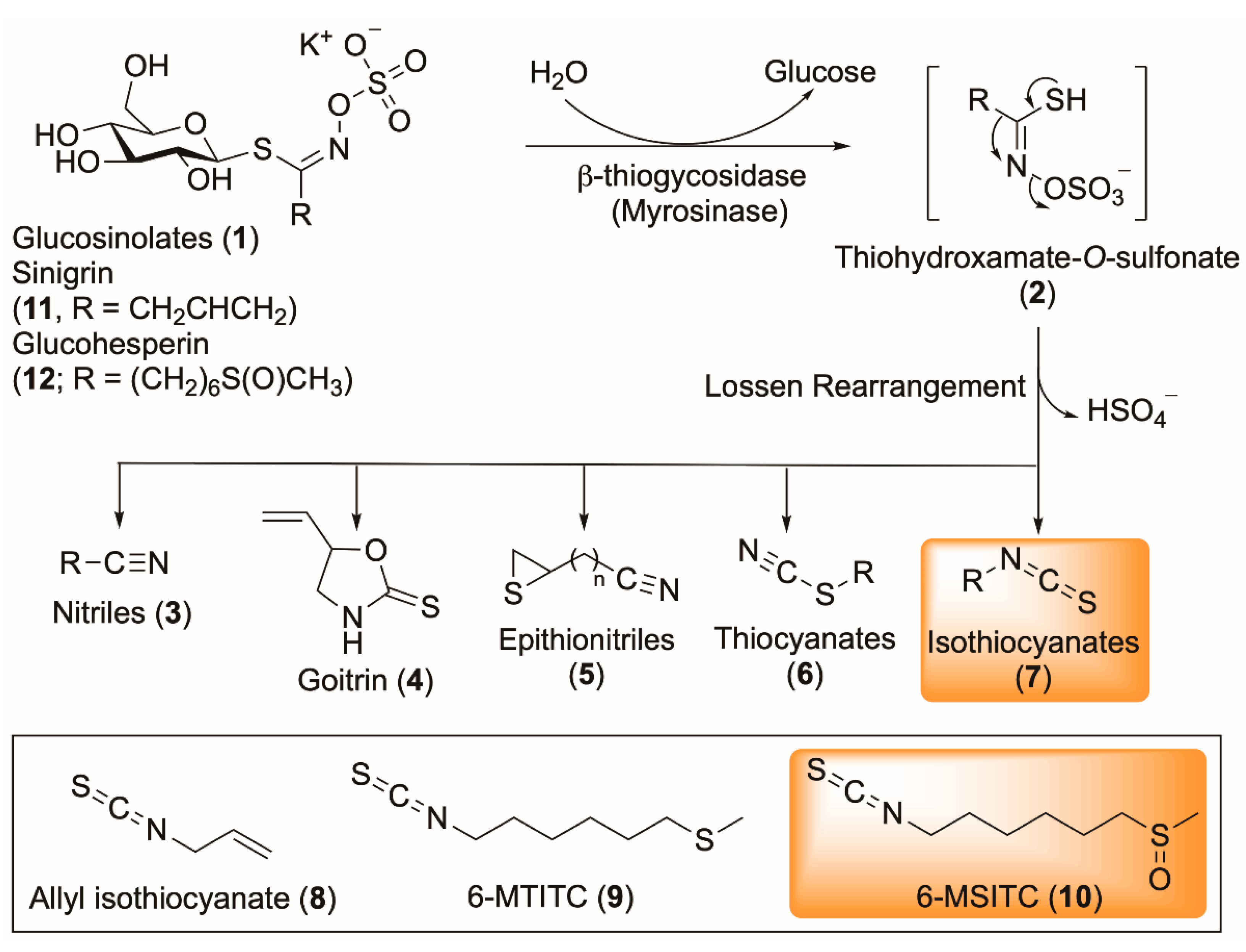
3.1. Composition of Wasabi Powder
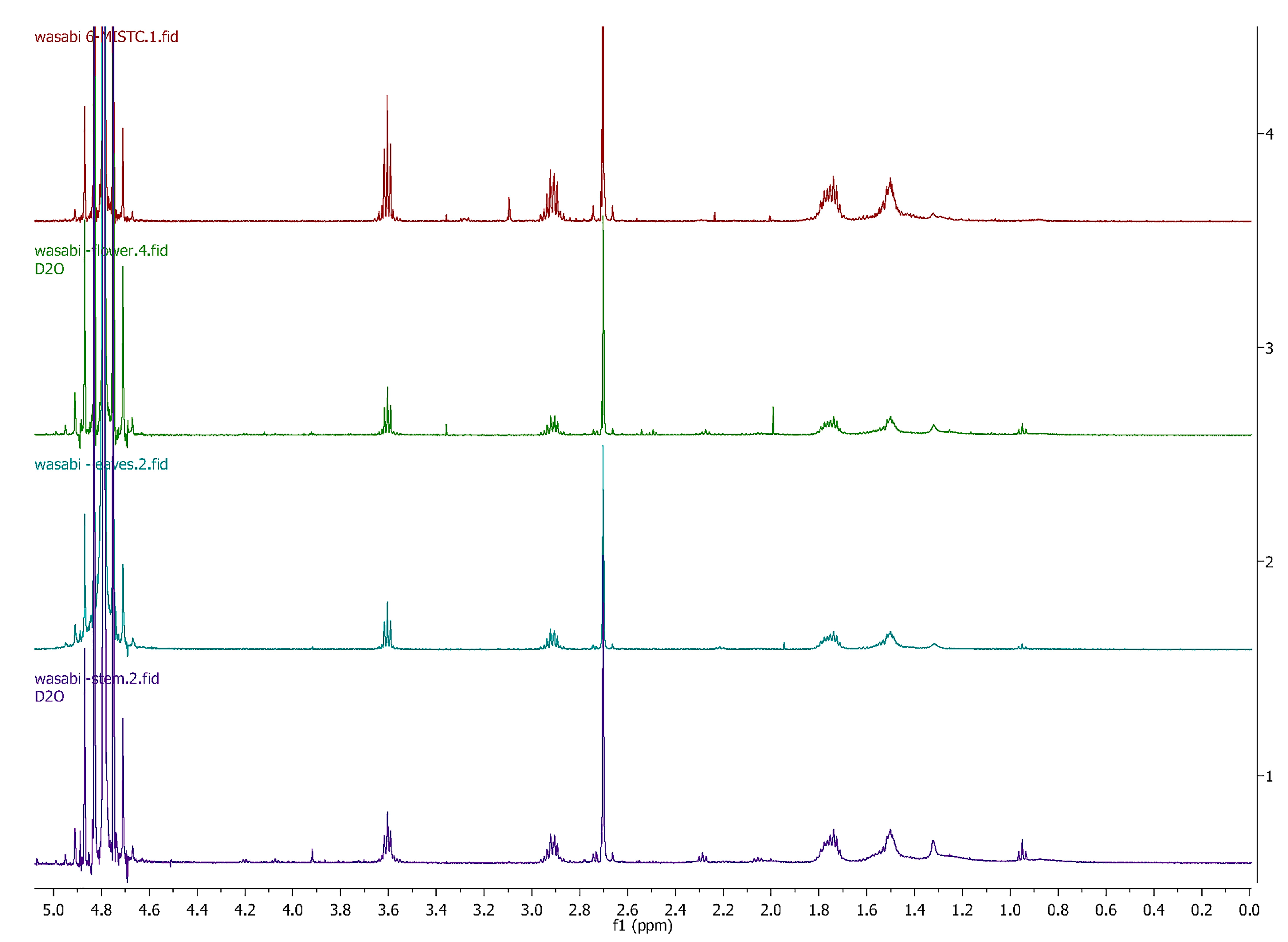
3.2. Comparing Eutrema Japonicum Flower, Leaves and Stem
3.3. Diet Intake, Body Composition and Metabolic Parameters
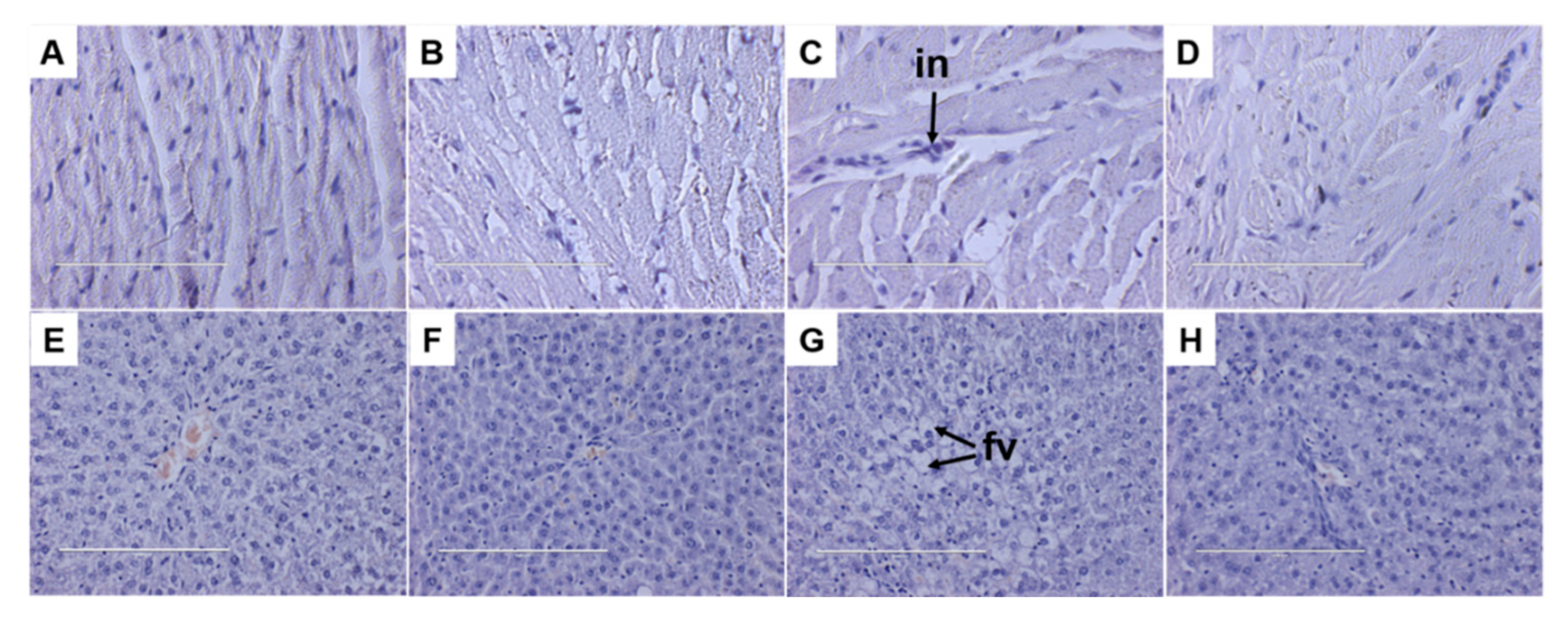
3.4. Liver and Heart Parameters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; Naghavi, M.; et al. Health effects of overweight and obesity in 195 countries over 25 years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Poudyal, H.; Panchal, S.K. Functional foods as potential therapeutic options for metabolic syndrome. Obes. Rev. 2015, 16, 914–941. [Google Scholar] [CrossRef] [PubMed]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef]
- Fenwick, G.R.; Heaney, R.K. Glucosinolates and their breakdown products in cruciferous crops, foods and feedingstuffs. Food Chem. 1983, 11, 249–271. [Google Scholar] [CrossRef]
- Holst, B.; Williamson, G. A critical review of the bioavailability of glucosinolates and related compounds. Nat. Prod. Rep. 2004, 21, 425–447. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef]
- Maina, S.; Misinzo, G.; Bakari, G.; Kim, H.-Y. Human, animal and plant health benefits of glucosinolates and strategies for enhanced bioactivity: A systematic review. Molecules 2020, 25, 3682. [Google Scholar] [CrossRef]
- Etoh, H.; Nishimura, A.; Takasawa, R.; Yagi, A.; Saito, K.; Sakata, K.; Kishima, I.; Ina, K. ω-Methylsulfinylalkyl isothiocyanates in wasabi, Wasabia japonica Matsum. Agric. Biol. Chem. 1990, 54, 1587–1589. [Google Scholar] [CrossRef]
- Ina, K.; Ina, H.; Ueda, M.; Yagi, A.; Kishima, I. ω-Methylthioalkyl isothiocyanates in wasabi. Agric. Biol. Chem. 1989, 53, 537–538. [Google Scholar] [CrossRef]
- Uto, T.; Hou, D.-X.; Morinaga, O.; Shoyama, Y. Molecular mechanisms underlying anti-inflammatory actions of 6-(methylsulfinyl)hexyl isothiocyanate derived from wasabi (Wasabia japonica). Adv. Pharmacol. Sci. 2012, 2012, 614046. [Google Scholar] [CrossRef]
- Nomura, T.; Shinoda, S.; Yamori, T.; Sawaki, S.; Nagata, I.; Ryoyama, K.; Fuke, Y. Selective sensitivity to wasabi-derived 6-(methylsulfinyl)hexyl isothiocyanate of human breast cancer and melanoma cell lines studied in vitro. Cancer Detect. Prev. 2005, 29, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, M.; Ogawa, T.; Wang, L.; Katsube, T.; Yamasaki, Y.; Sun, X.; Shiwaku, K. Anti-obesity effects of hot water extract from wasabi (Wasabia japonica Matsum.) leaves in mice fed high-fat diets. Nutr. Res. Pract. 2013, 7, 267–272. [Google Scholar] [CrossRef]
- Agoreyo, B.; Obansa, E.; Obanor, E. Comparative nutritional and phytochemical analyses of two varieties of Solanum melongena. Sci. World J. 2012, 7, 5–8. [Google Scholar]
- Fahey, G.C.; Novotny, L.; Layton, B.; Mertens, D.R. Critical factors in determining fiber content of feeds and foods and their ingredients. J. AOAC Int. 2019, 102, 52–62. [Google Scholar] [CrossRef]
- Dramur, M.; Bilgin, M.; Şahin, S. Investigation of oleuropein content in olive leaf extract obtained by supercritical fluid extraction and Soxhlet methods. Sep. Sci. Technol. 2011, 46, 1829–1837. [Google Scholar] [CrossRef]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, M.A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate, high-fat diet-induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Dockery, C.R.; Crosby, M.; Chavarria, K.; Patterson, B.; Giedd, M. Antibacterial activities of wasabi against Escherichia coli O157:H7 and Staphylococcus aureus. Front. Microbiol. 2016, 7, 1403. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.C.; Sorensen, J.L. Phytoalexin accumulation and antifungal compounds from the crucifer wasabi. Phytochemistry 1998, 49, 1959–1965. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Noshita, T.; Kidachi, Y.; Umetsu, H.; Fuke, Y.; Ryoyama, K. Detection of 6-(methylsulfinyl)hexyl isothiocyanate (6-MITC) and its conjugate with N-acetyl-L-cysteine (NAC) by high performance liquid chromatograpy-atmospheric pressure chemical ionization mass spectrometry (HPLC-MS/APCI). Chem. Pharm. Bull. 2008, 56, 715–719. [Google Scholar] [CrossRef][Green Version]
- Kumagai, H.; Kashima, N.; Seki, T.; Sakurai, H.; Ishii, K.; Ariga, T. Analysis of volatile components in essential oil of upland wasabi and their inhibitory effects on platelet aggregation. Biosci. Biotechnol. Biochem. 1994, 58, 2131–2135. [Google Scholar] [CrossRef]
- Depree, J.A.; Howard, T.M.; Savage, G.P. Flavour and pharmaceutical properties of the volatile sulphur compounds of wasabi (Wasabia japonica). Food Res. Int. 1998, 31, 329–337. [Google Scholar] [CrossRef]
- Jae-Joon, L.; Hyun, A.K.; Joomin, L. The effects of Brassica juncea L. leaf extract on obesity and lipid profiles of rats fed a high-fat/high-cholesterol diet. Nutr. Res. Pract. 2018, 12, 298–306. [Google Scholar] [CrossRef]
- Oowatari, Y.; Ogawa, T.; Katsube, T.; Iinuma, K.; Yoshitomi, H.; Gao, M. Wasabi leaf extracts attenuate adipocyte hypertrophy through PPARγ and AMPK. Biosci. Biotechnol. Biochem. 2016, 80, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Astell, K.; Mathai, M.; McAinch, A.; Stathis, C.; Su, X. A pilot study investigating the effect of Caralluma fimbriata extract on the risk factors of metabolic syndrome in overweight and obese subjects: A randomised controlled clinical trial. Complement. Ther. Med. 2013, 21, 180–189. [Google Scholar] [CrossRef]
- Bishnoi, M.; Khare, P.; Brown, L.; Panchal, S.K. Transient receptor potential (TRP) channels: A metabolic TR(i)P to obesity prevention and therapy. Obes. Rev. 2018, 19, 1269–1292. [Google Scholar] [CrossRef]
- Gees, M.; Alpizar, Y.A.; Boonen, B.; Sanchez, A.; Everaerts, W.; Segal, A.; Xue, F.; Janssens, A.; Owsianik, G.; Nilius, B.; et al. Mechanisms of transient receptor potential vanilloid 1 activation and sensitization by allyl isothiocyanate. Mol. Pharmacol. 2013, 84, 325–334. [Google Scholar] [CrossRef]
- Panchal, S.K.; Bliss, E.; Brown, L. Capsaicin in metabolic syndrome. Nutrients 2018, 10, 630. [Google Scholar] [CrossRef]
- Derbenev, A.V.; Zsombok, A. Potential therapeutic value of TRPV1 and TRPA1 in diabetes mellitus and obesity. Semin. Immunopathol. 2016, 38, 397–406. [Google Scholar] [CrossRef]
- Mao, Y.; Li, Z.; Chen, K.; Yu, H.; Zhang, S.; Jiang, M.; Ma, Y.; Liang, C.; Liu, H.; Li, H.; et al. Antinociceptive effect of ghrelin in a rat model of irritable bowel syndrome involves TRPV1/opioid systems. Cell. Physiol. Biochem. 2017, 43, 518–530. [Google Scholar] [CrossRef]
- Kawabata, F.; Kawabata, Y.; Liang, R.; Nishimura, S.; Tabata, S. Comparative analysis of allyl isothiocyanate (AITC)-induced carbohydrate oxidation changes via TRPV1 between mice and chickens. Biomed. Res. 2017, 38, 149–155. [Google Scholar] [CrossRef]
- Mori, N.; Kurata, M.; Yamazaki, H.; Matsumura, S.; Hashimoto, T.; Kanazawa, K.; Nadamoto, T.; Inoue, K.; Fushiki, T. Allyl isothiocyanate increases carbohydrate oxidation through enhancing insulin secretion by TRPV1. Biosci. Biotechnol. Biochem. 2018, 82, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Katsumura, T.; Motoi, M.; Oota, H.; Watanuki, S. Experimental evidence reveals the UCP1 genotype changes the oxygen consumption attributed to non-shivering thermogenesis in humans. Sci. Rep. 2017, 7, 5570. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N.; Hosoya, T.; Yoshida, S.; Sugimoto, O.; Yamada-Kato, T.; Kumazawa, S. 5-Hydroxyferulic acid methyl ester isolated from wasabi leaves inhibits 3T3-L1 adipocyte differentiation. Phytother. Res. 2018, 32, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.W.; Chen, C.S.; Chen, Y.C.; Hsu, Y.A.; Huang, C.C.; Chang, C.Y.; Lin, C.J.; Lin, C.W.; Lin, H.J.; Liu, F.T.; et al. Allyl isothiocyanate ameliorates obesity by inhibiting galectin-12. Mol. Nutr. Food Res. 2018, 62, e1700616. [Google Scholar] [CrossRef]
- Loke, W.M.; Hodgson, J.M.; Proudfoot, J.M.; McKinley, A.J.; Puddey, I.B.; Croft, K.D. Pure dietary flavonoids quercetin and (−)-epicatechin augment nitric oxide products and reduce endothelin-1 acutely in healthy men. Am. J. Clin. Nutr. 2008, 88, 1018–1025. [Google Scholar] [CrossRef]
- Quaschning, T.; Voss, F.; Relle, K.; Kalk, P.; Vignon-Zellweger, N.; Pfab, T.; Bauer, C.; Theilig, F.; Bachmann, S.; Kraemer-Guth, A.; et al. Lack of endothelial nitric oxide synthase promotes endothelin-induced hypertension: Lessons from endothelin-1 transgenic/endothelial nitric oxide synthase knockout mice. J. Am. Soc. Nephrol. 2007, 18, 730–740. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, J.; Kang, D.G.; Paek, Y.W.; Chung, D.J.; Chung, M.Y. Increased vascular endothelin-1 gene expression with unaltered nitric oxide synthase levels in fructose-induced hypertensive rats. Metabolism 2001, 50, 74–78. [Google Scholar] [CrossRef]
- Kalk, P.; Westermann, D.; Herzfeld, S.; Relle, K.; Pfab, T.; Bauer, C.; Tschope, C.; Stasch, J.P.; Hocher, B. Additional lack of iNOS attenuates diastolic dysfunction in aged ET-1 transgenic mice. Can. J. Physiol. Pharmacol. 2008, 86, 353–357. [Google Scholar] [CrossRef]
- Korenori, Y.; Tanigawa, S.; Kumamoto, T.; Qin, S.; Daikoku, Y.; Miyamori, K.; Nagai, M.; Hou, D.X. Modulation of Nrf2/Keap1 system by wasabi 6-methylthiohexyl isothiocyanate in ARE-mediated NQO1 expression. Mol. Nutr. Food Res. 2013, 57, 854–864. [Google Scholar] [CrossRef]
- Okamoto, T.; Akita, N.; Nagai, M.; Hayashi, T.; Suzuki, K. 6-Methylsulfinylhexyl isothiocyanate modulates endothelial cell function and suppresses leukocyte adhesion. J. Nat. Med. 2014, 68, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Subedi, L.; Venkatesan, R.; Kim, S.Y. Neuroprotective and anti-inflammatory activities of allyl isothiocyanate through attenuation of JNK/NF-κB/TNF-α signaling. Int. J. Mol. Sci. 2017, 18, 1423. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Fujii, M.; Hou, D.X. Inhibition of lipopolysaccharide-induced cyclooxygenase-2 transcription by 6-(methylsulfinyl) hexyl isothiocyanate, a chemopreventive compound from Wasabia japonica (Miq.) Matsumura, in mouse macrophages. Biochem. Pharmacol. 2005, 70, 1772–1784. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, M. Stress-sensing mechanisms and the physiological roles of the Keap1-Nrf2 system during cellular stress. J. Biol. Chem. 2017, 292, 16817–16824. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Uto, T.; Tanigawa, S.; Yamada-Kato, T.; Fujii, M.; Hou, D.E.X. Microarray-based determination of anti-inflammatory genes targeted by 6-(methylsulfinyl)hexyl isothiocyanate in macrophages. Exp. Ther. Med. 2010, 1, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, K.; Sato, S.; Sakakibara, Y.; Mori, S.; Ohkuma, M.; Kawashima, Y.; Ban, T.; Sasaki, H.; Tachiyashiki, K. Allyl isothiocyanate-induced changes in the distribution of white blood cells in rats. J. Toxicol. Sci. 2010, 35, 583–589. [Google Scholar] [CrossRef][Green Version]
- Kang, J.H.; Choi, S.; Jang, J.E.; Ramalingam, P.; Ko, Y.T.; Kim, S.Y.; Oh, S.H. Wasabia japonica is a potential functional food to prevent colitis via inhibiting the NF-κB signaling pathway. Food Funct. 2017, 8, 2865–2874. [Google Scholar] [CrossRef]
- Kim, M.W.; Choi, S.; Kim, S.Y.; Yoon, Y.S.; Kang, J.H.; Oh, S.H. Allyl isothiocyanate ameliorates dextran sodium sulfate-induced colitis in mouse by enhancing tight junction and mucin expression. Int. J. Mol. Sci. 2018, 19, 2025. [Google Scholar] [CrossRef]
- Park, B.K.; Chun, E.; Choi, J.J.; Shin, Y.; Kho, Y.T.; Oh, S.H.; Kim, S.Y.; Lee, T.H.; Kim, T.W.; Shin, E.; et al. Administration of Wasabia koreana ameliorates irritable bowel syndrome-like symptoms in a zymosan-induced mouse model. J. Med. Food 2017, 20, 474–484. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Kirpotina, L.N.; Khlebnikov, A.I.; Balasubramanian, N.; Quinn, M.T. Neutrophil immunomodulatory activity of natural organosulfur compounds. Molecules 2019, 24, 1809. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, D.H.; Ahn, J.; Chung, W.J.; Jang, Y.J.; Seong, K.S.; Moon, J.H.; Ha, T.Y.; Jung, C.H. Pharmacokinetics, tissue distribution, and anti-lipogenic/adipogenic effects of allyl-isothiocyanate metabolites. PLoS ONE 2015, 10, e0132151. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Composition | Value |
|---|---|
| Ash (%) | 9.25 ± 0.04 |
| Fat (%) | 0.96 ± 0.01 |
| Moisture (%) | 11.30 ± 0.00 |
| Carbohydrate (%) | 71.5% |
| Dietary fibre (%) | 5.1% ± 0.00 |
| Protein (%) | 1.78 ± 0.03 |
| Elements (mg/g) | |
| Copper | 0.03 ± 0.50 |
| Calcium | 5.76 ± 0.05 |
| Iron | 0.02 ± 0.00 |
| Magnesium | 1.33 ± 0.01 |
| Manganese | 0.01 ± 0.50 |
| Phosphorus | 3.09 ± 0.03 |
| Potassium | 32.02 ± 0.59 |
| Sodium | 0.08 ± 0.01 |
| Sulphur | 7.66 ± 0.08 |
| Zinc | 0.01 ± 0.00 |
| Variables | C | CW | H | HW | p-Value | ||
|---|---|---|---|---|---|---|---|
| Diet | Wasabi | Interaction | |||||
| Water intake (mL/day) | 40.6 ± 3.8 a | 40.0 ± 1.7 a | 33.3 ± 2.5 b | 29.5 ± 1.4 c | 0.74 | 0.34 | 0.4900 |
| Food intake (g/day) | 36.9 ± 1.3 a | 36.5 ± 2.6 a | 27.1 ± 2.1 b | 24.4 ± 0.9 c | 0.47 | 0.01 | <0.0001 |
| Energy intake (kJ/day) | 413 ± 33 c | 410 ± 33 c | 553 ± 82 a | 484 ± 48 b | 0.47 | 0.0007 | <0.0001 |
| Feed efficiency (kJ/g) | 0.15 ± 0.01 bc | 0.10 ± 0.01 c | 0.36 ± 0.03 a | 0.23 ± 0.02 b | 0.43 | 0.16 | <0.0001 |
| Body weight (g) | 368 ± 5.3 a | 350 ± 4.2 b | 460 ± 9.5 c | 416 ± 3.6 d | 0.03 | 0.04 | <0.0001 |
| Abdominal circumference (cm) | 18.3 ± 0.3 c | 17.8 ± 0.2 bc | 21.3 ± 1.0 a | 19.7 ± 0.3 b | 0.005 | <0.0001 | <0.0001 |
| Whole-body lean mass (g) | 289 ± 7 b | 289 ± 8 b | 318 ± 14 a | 315 ± 15 a | 0.88 | 0.25 | 0.0001 |
| Whole-body fat mass (g) | 89 ± 9 ab | 60 ± 6 c | 178 ± 14 a | 120 ± 23 b | <0.0001 | 0.02 | <0.0001 |
| Wasabi intake (g/day) | - | 1.7 ± 0.1 a | - | 1.0 ± 0.1 b | - | - | - |
| Blood glucose area under the curve (mmol/L × min) | 681 ± 24 b | 541 ± 27 c | 729 ± 34 a | 663 ± 20 b | 0.19 | 0.004 | 0.0006 |
| Systolic blood pressure (mmHg) | 131 ± 4 b | 128 ± 2 c | 146 ± 2 a | 132 ± 2 b | <0.0001 | 0.0005 | <0.0001 |
| Plasma aspartate transaminase (U/L) | 66.3 ± 3.7 b | 64.0 ± 4.0 c | 94.9 ± 10.4 a | 67.0 ± 5.0 b | 0.32 | 0.01 | <0.0001 |
| Plasma alanine transaminase (U/L) | 26.1 ± 5.2 b | 24.0 ± 2.0 c | 37.9 ± 4.0 a | 37.0 ± 4.0 a | 0.39 | 0.43 | <0.0001 |
| Triglycerides (mmol/L) | 0.5 ± 0.1 c | 0.5 ± 0.04 c | 1.7 ± 0.3 a | 0.9 ± 0.3 b | 0.37 | 0.02 | <0.0001 |
| Total cholesterol (mmol/L) | 1.4 ± 0.1 a | 1.0 ± 0.1 b | 1.5 ± 0.1 a | 1.0 ± 0.04 b | 0.11 | 0.26 | <0.0001 |
| Retroperitoneal fat (mg/mm tibial length) | 169 ± 19 b | 121 ± 6 c | 398 ± 67 a | 246 ± 24 b | <0.0001 | <0.0001 | <0.0001 |
| Epididymal fat (mg/mm tibial length) | 90 ± 5 b | 75 ± 13 c | 172 ± 20 a | 123 ± 18 b | <0.0001 | <0.0001 | <0.0001 |
| Omental fat (mg/mm tibial length) | 131 ± 10 b | 85 ± 6 c | 220 ± 13 a | 161 ± 20 b | <0.0001 | <0.0001 | <0.0001 |
| Total abdominal fat (mg/mm tibial length) | 390 ± 20 ab | 281 ± 20 c | 790 ± 97 a | 530 ± 58 b | <0.0001 | <0.0001 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomaz, F.S.; Tan, Y.P.; Williams, C.M.; Ward, L.C.; Worrall, S.; Panchal, S.K. Wasabi (Eutrema japonicum) Reduces Obesity and Blood Pressure in Diet-Induced Metabolic Syndrome in Rats. Foods 2022, 11, 3435. https://doi.org/10.3390/foods11213435
Thomaz FS, Tan YP, Williams CM, Ward LC, Worrall S, Panchal SK. Wasabi (Eutrema japonicum) Reduces Obesity and Blood Pressure in Diet-Induced Metabolic Syndrome in Rats. Foods. 2022; 11(21):3435. https://doi.org/10.3390/foods11213435
Chicago/Turabian StyleThomaz, Fernanda Santos, Yuen P. Tan, Craig M. Williams, Leigh C. Ward, Simon Worrall, and Sunil K. Panchal. 2022. "Wasabi (Eutrema japonicum) Reduces Obesity and Blood Pressure in Diet-Induced Metabolic Syndrome in Rats" Foods 11, no. 21: 3435. https://doi.org/10.3390/foods11213435
APA StyleThomaz, F. S., Tan, Y. P., Williams, C. M., Ward, L. C., Worrall, S., & Panchal, S. K. (2022). Wasabi (Eutrema japonicum) Reduces Obesity and Blood Pressure in Diet-Induced Metabolic Syndrome in Rats. Foods, 11(21), 3435. https://doi.org/10.3390/foods11213435