
Potential Correlation between Microbial Diversity and Volatile Flavor Compounds in Different Types of Korean Dry-Fermented Sausages


Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Sausage Samples
2.2. Estimation of Physicochemical Characteristics of Sausage Samples
2.2.1. Proximate Composition, pH, and Water Activity (aw)
2.2.2. Color Measurement
2.3. HTS of Sausage DNA
2.3.1. DNA Extraction
2.3.2. Illumina HTS
2.3.3. Data Analysis
2.4. Aroma and Taste Profiling
2.4.1. Volatile Compound Profiling Using an Electronic Nose
2.4.2. Taste Profiling Using an Electronic Tongue
2.5. Statistical Analysis
3. Results
3.1. Physicochemical Properties of the Sausage Samples
3.2. Microbial Richness and Diversity
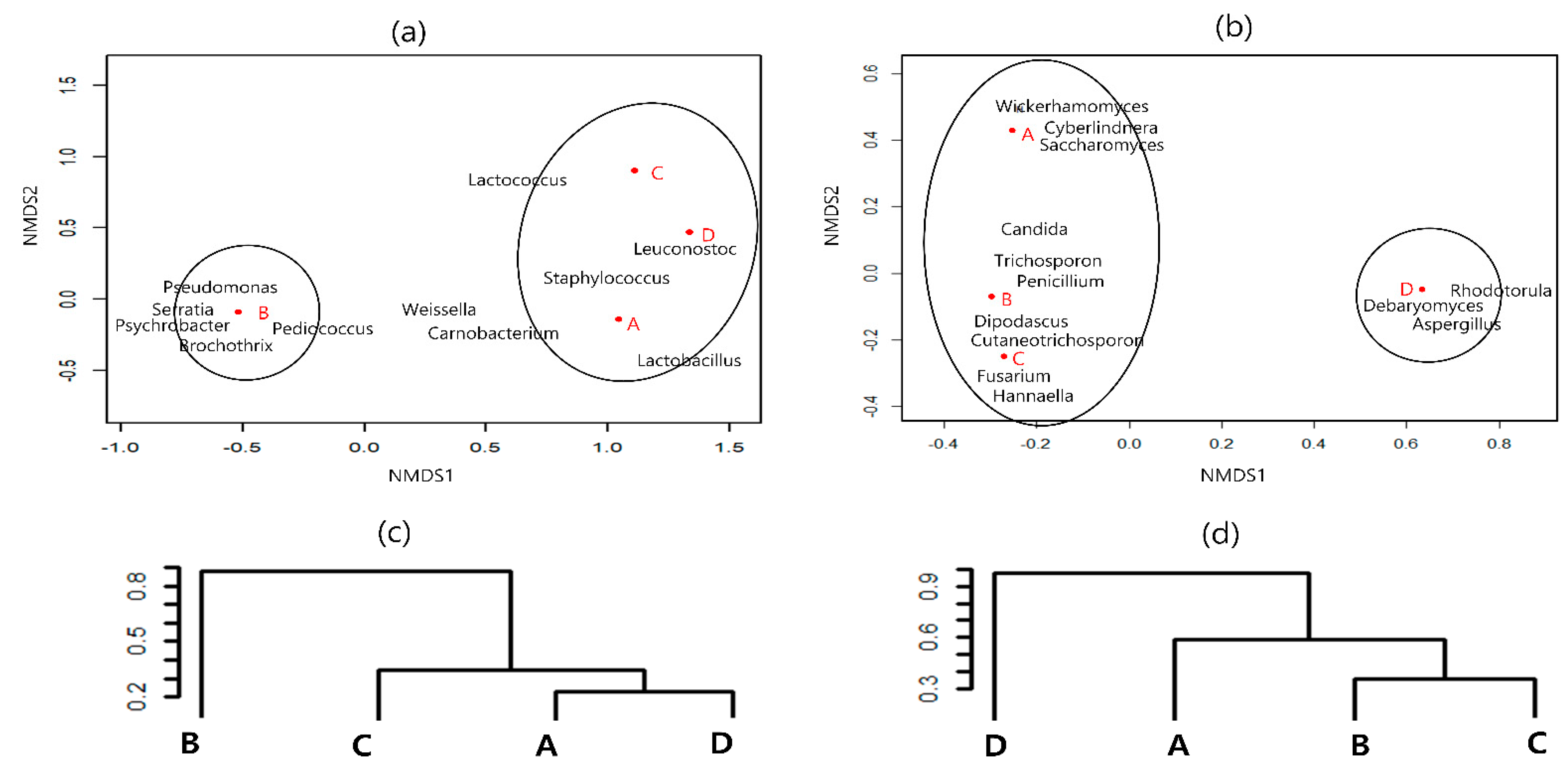
3.3. Similarity of Microbial Communities
3.4. Composition of the Microbial Communities
3.5. E-Nose and E-Tongue Analysis
3.6. Potential Correlation of Microbiota, Flavors, and Tastes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paik, H.D.; Lee, J.Y. Investigation of Reduction and Tolerance Capability of Lactic Acid Bacteria Isolated from Kimchi against Nitrate and Nitrite in Fermented Sausage Condition. Meat Sci. 2014, 97, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, S.; Cachaldora, A.; Gómez, M.; Franco, I.; Carballo, J. Monitoring the Bacterial Population Dynamics During the Ripening of Galician Chorizo, a Traditional Dry Fermented Spanish Sausage. Food Microbiol. 2013, 33, 77–84. [Google Scholar] [CrossRef]
- Sidira, M.; Kandylis, P.; Kanellaki, M.; Kourkoutas, Y. Effect of Immobilized Lactobacillus casei on the Evolution of Flavor Compounds in Probiotic Dry-Fermented Sausages During Ripening. Meat Sci. 2015, 100, 41–51. [Google Scholar] [CrossRef]
- Połka, J.; Rebecchi, A.; Pisacane, V.; Morelli, L.; Puglisi, E. Bacterial Diversity in Typical Italian Salami at Different Ripening Stages as Revealed by High-Throughput Sequencing of 16S RRNA Amplicons. Food Microbiol. 2015, 46, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hou, J.; Zhang, X.; Hu, J.; Yu, Z.; Zhu, Y. Improving the Flavor of Fermented Sausage by Increasing Its Bacterial Quality via Inoculation with Lactobacillus plantarum MSZ2 and Staphylococcus xylosus YCC3. Foods 2022, 11, 736. [Google Scholar] [CrossRef]
- Magistà, D.; Susca, A.; Ferrara, M.; Logrieco, A.F.; Perrone, G. Penicillium species: Crossroad between Quality and Safety of Cured Meat Production. Curr. Opin. Food Sci. 2017, 17, 36–40. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, L.; Liu, Q.; Wang, Y.; Chen, Q.; Kong, B. The Potential Correlation between Bacterial Diversity and the Characteristic Volatile Flavour of Traditional Dry Sausages from Northeast China. Food Microbiol. 2020, 91, 103505. [Google Scholar] [CrossRef]
- Meng, Y.; Chen, X.; Sun, Z.; Li, Y.; Chen, D.; Fang, S.; Chen, J. Exploring Core Microbiota Responsible for the Production of Volatile Flavor Compounds During the Traditional Fermentation of Koumiss. LWT 2021, 135, 110049. [Google Scholar] [CrossRef]
- Prado, N.; Sampayo, M.; González, P.; Lombó, F.; Díaz, J. Physicochemical, Sensory and Microbiological Characterization of Asturian Chorizo, a Traditional Fermented Sausage Manufactured in Northern Spain. Meat Sci. 2019, 156, 118–124. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Kuczynski, J.; Stombaugh, J.; Walters, W.A.; González, A.; Caporaso, J.G.; Knight, R. Using QIIME to Analyze 16S rRNA Gene Sequences from Microbial Communities. Curr. Protoc. Microbiol. 2012, 27, 1E-5. [Google Scholar] [CrossRef] [PubMed]
- Go, H.Y.; Lee, S.H.; Kim, H.Y. The Effect of Hot-Air Dried Lentinula Edodes on the Quality and Oranoleptic Properties of Rolled-Dumplings. Food Sci. Anim. Resour. 2022, 42, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Dalmış, U.; Soyer, A. Effect of Processing Methods and Starter Culture (Staphylococcus xylosus and Pediococcus pentosaceus) on Proteolytic Changes in Turkish Sausages (Sucuk) During Ripening and Storage. Meat Sci. 2008, 80, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Casaburi, A.; Aristoy, M.C.; Cavella, S.; Di Monaco, R.; Ercolini, D.; Toldrá, F.; Villani, F. Biochemical and Sensory Characteristics of Traditional Fermented Sausages of Vallo di Diano (Southern Italy) as Affected by the Use of Starter Cultures. Meat Sci. 2007, 76, 295–307. [Google Scholar] [CrossRef]
- Porto-Fett, A.C.S.; Hwang, C.A.; Call, J.E.; Juneja, V.K.; Ingham, S.C.; Ingham, B.H.; Luchansky, J.B. Viability of Multi-strain Mixtures of Listeria monocytogenes, Salmonella typhimurium, or Escherichia coli O157: H7 Inoculated into the Batter or onto the Surface of a Soudjouk-Style Fermented Semi-dry Sausage. Food Microbiol. 2008, 25, 793–801. [Google Scholar] [CrossRef]
- Wang, X.; Ren, H.; Zhan, Y. Characterization of Microbial Community Composition and Pathogens Risk Assessment in Typical Italian-Style Salami by High-Throughput Sequencing Technology. Food Sci. Biotechnol. 2018, 27, 241–249. [Google Scholar] [CrossRef]
- Aquilanti, L.; Garofalo, C.; Osimani, A.; Clementi, F. Ecology of Lactic Acid Bacteria and Coagulase Negative Cocci in Fermented Dry Sausages Manufactured in Italy and Other Mediterranean Countries: An Overview. Int. Food Res. J. 2016, 23, 429–445. [Google Scholar]
- Charmpi, C.; Van der Veken, D.; Van Reckem, E.; De Vuyst, L.; Leroy, F. Raw Meat Quality and Salt Levels Affect the Bacterial Species Diversity and Community Dynamics During the Fermentation of Pork Mince. Food Microbiol. 2020, 89, 103434. [Google Scholar] [CrossRef]
- Laranjo, M.; Potes, M.E.; Elias, M. Role of Starter Cultures on the Safety of Fermented Meat Products. Front. Microbiol. 2019, 10, 853. [Google Scholar] [CrossRef]
- Cho, J.I.; Joo, I.S.; Choi, J.H.; Jung, K.H.; Choi, E.J.; Son, N.R.; Han, M.K.; Jeong, S.J.; Lee, S.H.; Hwang, I.G. Distribution of Methicillin-Resistant Staphylococcus aureus (MRSA) in RAW Meat and Fish Samples in Korea. Food Sci. Biotechnol. 2014, 23, 999–1003. [Google Scholar] [CrossRef]
- Ravyts, F.; Vuyst, L.D.; Leroy, F. Bacterial Diversity and Functionalities in Food Fermentations. Eng. Life Sci. 2012, 12, 356–367. [Google Scholar] [CrossRef]
- Cappello, M.S.; Zapparoli, G.; Logrieco, A.; Bartowsky, E.J. Linking Wine Lactic Acid Bacteria Diversity with Wine Aroma and Flavour. Int. J. Food Microbiol. 2017, 243, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Kohno, Y.; Makino, T.; Kanakubo, M. Effect of Phase Behavior for Ionic Liquid Catalysts with Reactants/Products on Reactivity of Esterification from Long-Chain Fatty Alcohols and Fatty Acids. Fluid Phase Equilibria 2019, 490, 107–113. [Google Scholar] [CrossRef]
- Casaburi, A.; Piombino, P.; Nychas, G.J.; Villani, F.; Ercolini, D. Bacterial Populations and the Volatilome Associated to Meat Spoilage. Food Microbiol. 2015, 45, 83–102. [Google Scholar] [CrossRef]
- Solina, M.; Baumgartner, P.; Johnson, R.L.; Whitfield, F.B. Volatile Aroma Components of Soy Protein Isolate and Acid-Hydrolysed Vegetable Protein. Food Chem. 2005, 90, 861–873. [Google Scholar] [CrossRef]
- Feng, L.; Tang, N.; Liu, R.; Gong, M.; Wang, Z.; Guo, Y.; Wang, Y.; Zhang, Y.; Chang, M. The Relationship between Flavor Formation, Lipid Metabolism, and Microorganisms in Fermented Fish Products. Food Funct. 2021, 12, 5685–5702. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Cachaldora, A.; Fonseca, S.; Gómez, M.; Franco, I.; Carballo, J. Production of Biogenic Amines “In Vitro” in Relation to the Growth Phase by Enterobacteriaceae species Isolated from Traditional Sausages. Meat Sci. 2010, 86, 684–691. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Martinez, S.; Franco, I.; Carballo, J. Biogenic Amine Content in Relation to Physicochemical Parameters and Microbial Counts in Two Kinds of Spanish Traditional Sausages. Arch. Lebensmittelhyg. 2008, 59, 70–75. [Google Scholar]
- Bodey, G.P.; Bolivar, R.; Fainstein, V.; Jadeja, L. Infections Caused by Pseudomonas aeruginosa. Rev. Infect. Dis. 1983, 5, 279–313. [Google Scholar] [CrossRef]
- Russo, F.; Ercolini, D.; Mauriello, G.; Villani, F. Behaviour of Brochothrix thermosphacta in Presence of Other Meat Spoilage Microbial Groups. Food Microbiol. 2006, 23, 797–802. [Google Scholar] [CrossRef]
- Huang, L.; Wang, Y.; Li, R.; Wang, Q.; Dong, J.; Wang, J.; Lu, S. Thyme Essential Oil and Sausage Diameter Effects on Biogenic Amine Formation and Microbiological Load in Smoked Horse Meat Sausage. Food Biosci. 2021, 40, 100885. [Google Scholar] [CrossRef]
- Lee, E.; Choe, E. Changes in Oxidation-Derived Off-Flavor Compounds of Roasted Sesame Oil During Accelerated Storage in the Dark. Biocatal. Agric. Biotechnol. 2012, 1, 89–93. [Google Scholar] [CrossRef]
- Sánchez-Montero, L.; Córdoba, J.J.; Peromingo, B.; Álvarez, M.; Núñez, F. Effects of Environmental Conditions and Substrate on Growth and Ochratoxin a Production by Penicillium verrucosum and Penicillium nordicum: Relative Risk Assessment of OTA in Dry-Cured Meat Products. Food Res. Int. 2019, 121, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Cao, Y.; Zhang, Y.; Zhang, Z.; Shi, X.; Zhang, W.; Wen, P. Effects of Storage Methods on the Microbial Community and Quality of Sichuan Smoked Bacon. LWT Food Sci. Technol. 2022, 158, 113115. [Google Scholar] [CrossRef]
- Ogunremi, O.R.; Sanni, A.I.; Agrawal, R. Probiotic Potentials of Yeasts Isolated from Some Cereal-Based N Igerian Traditional Fermented Food Products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast Spoilage of Foods and Beverages. In The Yeasts; Elsevier: Amsterdam, The Netherlands, 2011; pp. 53–63. [Google Scholar]
- Andrade, M.J.; Thorsen, L.; Rodríguez, A.; Córdoba, J.J.; Jespersen, L. Inhibition of Ochratoxigenic Moulds by Debaryomyces hansenii Strains for Biopreservation of Dry-Cured Meat Products. Int. J. Food Microbiol. 2014, 170, 70–77. [Google Scholar] [CrossRef]
- Trofa, D.; Gácser, A.; Nosanchuk, J.D. Candida parapsilosis, an Emerging Fungal Pathogen. Clin. Microbiol. Rev. 2008, 21, 606–625. [Google Scholar] [CrossRef]
- Shukla, S.; Park, J.; Park, J.H.; Kim, M.K.; Park, S.; Dubey, A.; Jeon, J.; Khang, Y.; Kim, M. Evaluation of Fungal Microflora for Aflatoxin Producing Possibility in Novel Quality Meju Fermented with Single and/or Multiple Additions of Nelumbo nucifera, Ginkgo biloba, and Allium sativum Extracts. J. Food Saf. 2017, 37, e12368. [Google Scholar] [CrossRef]
- Guo, J.; Qin, D.; Li, W.; Wu, F.; Li, L.; Liu, X. Inactivation of Penicillium italicum on Kumquat via Plasma-Activated Water and Its Effects on Quality Attributes. Int. J. Food Microbiol. 2021, 343, 109090. [Google Scholar] [CrossRef]
- Zhou, C.; Zhan, G.; Pan, D.; Zhou, G.; Wang, Y.; He, J.; Cao, J. Charactering the Spoilage Mechanism of “Three Sticks” of Jinhua Ham. Food Sci. Hum. Wellness 2022, 11, 1322–1330. [Google Scholar] [CrossRef]
- Liu, L.; Chen, X.; Hao, L.; Zhang, G.; Jin, Z.; Li, C.; Yang, Y.; Rao, J.; Chen, B. Traditional Fermented Soybean Products: Processing, Flavor Formation, Nutritional and Biological Activities. Crit. Rev. Food Sci. Nutr. 2022, 62, 1971–1989. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Starter Culture | Ingredient List | Fermentation Period | |
|---|---|---|---|---|
| Lactic Acid Bacteria (LAB) | Mold | |||
| A | Lactiplantibacillus plantarum | Penicillium nalgiovens | Pork shoulders, pork fat, white sugar, refined salt, onion, garlic, ginger, pepper powder, kelp, shiitake mushroom, yeast extract, vegetable mixed powder, barbecue flavor seasoning, collagen casing | 40–45 d |
| B | Pediococcus acidilactici, Latilactobacillus curvatus, Staphylococcus xylosus | Penicillium nalgiovens | Pork shoulders, pork fat, pepper powder, glucose, garlic, sodium nitrite, natural (pig intestine) casing | 25–30 d |
| C | Non-starter | Penicillium nalgiovens | Pork shoulders, pork fat, red wine, salt, black pepper, garlic, nitrite, ascorbic acid | 40–45 d |
| D | Mixed LABs (21 species) | Non-starter | Pork shoulders, pork fat, slat 1%, herb mix (coriander, clove, fennel, oregano), garlic, black pepper, salt, nitrate | 60–65 d |
| Sausage Products | ||||
|---|---|---|---|---|
| Parameter | A (n = 3) | B (n = 3) | C (n = 3) | D (n = 3) |
| pH | 4.70 ± 0.19 c | 5.87 ± 0.03 a | 3.62 ± 0.08 d | 5.05 ± 0.07 b |
| aw | 0.82 ± 0.00 b | 0.78 ± 0.00 a | 0.82 ± 0.01 b | 0.85 ± 0.01 a |
| Moisture (%) | 29.31 ± 0.31 b | 27.39 ± 0.15 c | 28.93 ± 0.16 b | 30.96 ± 0.30 a |
| Fat (%) | 33.22 ± 0.59 b | 29.46 ± 0.44 c | 34.38 ± 0.42 a | 32.71 ± 0.60 b |
| Protein (%) | 35.28 ± 0.15 b | 38.12 ± 0.13 a | 34.38 ± 0.33 c | 34.23 ± 0.11 c |
| Collagen (%) | 2.59 ± 0.03 b | 4.43 ± 0.15 a | 2.15 ± 0.04 c | 2.51 ± 0.06 b |
| L* value | 46.58 ± 1.36 a | 38.22 ± 1.33 c | 40.37 ± 1.51 b | 47.05 ± 1.58 a |
| a* value | 8.62 ± 1.22 c | 11.62 ± 0.69 b | 12.80 ± 0.61 a | 12.77 ± 0.69 a |
| b* value | 9.03 ± 0.63 a | 6.49 ± 0.98 c | 8.04 ± 0.66 b | 8.88 ± 0.26 a |
| Section diameter (mm) | 40 | 26 | 40 | 60 |
| Sample | Reads | Observed OTUs | Chao 1 Index | Good’s Coverage | Shannon Index | Simpson | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | |
| A | 420,848 | 156,310 | 28 | 14 | 28 | 14 | 1.0 | 1.0 | 4.10 | 2.03 | 0.92 | 0.68 |
| B | 131,765 | 193,906 | 36 | 12 | 45 | 12 | 1.0 | 1.0 | 4.43 | 1.27 | 0.95 | 0.40 |
| C | 135,926 | 218,900 | 6 | 24 | 15 | 12 | 1.0 | 1.0 | 2.24 | 1.37 | 0.87 | 0.45 |
| D | 382,850 | 182,988 | 17 | 12 | 19 | 12 | 1.0 | 1.0 | 3.53 | 1.32 | 0.90 | 0.44 |
| Variables | Volatile Flavor | Taste Profile | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2-Propanone | Methanethiol | Ethanol | Pentane | 2-Methylfuran | Ethyl acetate | Thiopherne | Trichloroethylene | 4-Methylnonane | 2,4,5-Trimethylthiazole | Cyclopentane | Limonene | AHS_ Sourness | PKS_ Sweets | CTS_ Saltiness | NMS_ Umami | ANS_ Bitters | CPS | SCS | |
| Bacteria | |||||||||||||||||||
| Lactobacillus | 0.398 | −0.402 | 0.331 | −0.997 ** | 0.564 | 0.805 | 0.363 | 0.604 | 0.256 | 0.173 | −0.022 | −0.002 | −0.909 | −0.855 | 0.735 | −0.971 * | −0.91 | −0.936 | −0.906 |
| Carnobacterium | 0.809 | 0.254 | 0.357 | 0.249 | 0.595 | 0.276 | 0.268 | 0.407 | −0.737 | −0.83 | −0.857 | −0.798 | −0.115 | −0.314 | −0.34 | 0.097 | −0.109 | −0.021 | −0.13 |
| Brochothrix | −0.486 | 0.243 | −0.502 | 0.974 * | −0.591 | −0.906 | −0.525 | −0.748 | −0.061 | 0.02 | 0.21 | 0.196 | 0.973 * | 0.932 | −0.601 | 0.932 | 0.974 * | 0.986 * | 0.971 * |
| Pediococcus | −0.486 | 0.243 | −0.502 | 0.8 | −0.591 | −0.906 | −0.525 | −0.748 | −0.061 | 0.02 | 0.21 | 0.196 | 0.973 * | 0.932 | −0.601 | 0.932 | 0.974 * | 0.986 * | 0.971 * |
| Leuconostoc | 0.958 * | −0.443 | 0.112 | −0.399 | 0.994 ** | 0.959 * | 0.051 | 0.425 | −0.124 | −0.295 | −0.469 | −0.366 | −0.516 | −0.684 | 0.469 | −0.619 | −0.51 | −0.463 | −0.527 |
| Pseudomonas | −0.486 | 0.243 | −0.502 | 0.974 * | −0.591 | −0.906 | −0.525 | −0.748 | −0.061 | 0.02 | 0.21 | 0.196 | 0.973 * | 0.932 | −0.601 | 0.932 | 0.974 * | 0.986 * | 0.971 * |
| Staphylococcus | −0.351 | −0.838 | −0.707 | −0.376 | −0.018 | −0.265 | −0.653 | −0.524 | 0.924 | 0.972 * | 0.898 | 0.925 | 0.061 | 0.146 | 0.816 | −0.434 | 0.059 | −0.016 | 0.07 |
| Lactococcus | 0.946 * | 0.148 | 0.526 | −0.132 | 0.784 | 0.601 | 0.45 | 0.666 | −0.672 | −0.791 | −0.887 | −0.827 | −0.475 | −0.648 | −0.095 | −0.259 | −0.469 | −0.391 | −0.488 |
| Weissella | 0.375 | 0.64 | 0.004 | 0.748 | 0.144 | −0.289 | −0.08 | −0.095 | −0.625 | −0.653 | −0.568 | −0.529 | 0.455 | 0.272 | −0.611 | 0.602 | 0.461 | 0.538 | 0.443 |
| Psychrobacter | −0.486 | 0.243 | −0.502 | 0.974 * | −0.591 | −0.906 | −0.525 | −0.748 | −0.061 | 0.02 | 0.21 | 0.196 | 0.973 * | 0.932 | −0.601 | 0.932 | 0.974 * | 0.986 * | 0.971 * |
| Serratia | −0.486 | 0.243 | −0.502 | 0.974 * | −0.591 | −0.906 | −0.525 | −0.748 | −0.061 | 0.02 | 0.21 | 0.196 | 0.973 * | 0.932 | −0.601 | 0.932 | 0.974 * | 0.986 * | 0.971 * |
| Fungal | |||||||||||||||||||
| Penicillium | −0.518 | 0.528 | 0.506 | 0.394 | −0.756 | −0.086 | 0.53 | 0.151 | −0.56 | −0.408 | −0.243 | −0.352 | 0.199 | 0.303 | −0.852 | 0.646 | 0.194 | 0.201 | 0.203 |
| Debaryomyces | −0.037 | −0.963 * | −0.675 | −0.465 | 0.297 | −0.117 | −0.645 | −0.399 | 0.93 | 0.853 | 0.729 | 0.79 | −0.076 | −0.051 | 0.935 | −0.598 | −0.076 | −0.133 | −0.071 |
| Candida | 0.538 | 0.936 | 0.383 | 0.521 | 0.252 | 0.072 | 0.301 | 0.29 | −0.861 | −0.893 | −0.839 | −0.816 | 0.123 | −0.046 | −0.663 | 0.437 | 0.128 | 0.215 | 0.11 |
| Wickerhamomyces | 0.946 | 0.148 | 0.526 | −0.132 | 0.784 | 0.601 | 0.45 | 0.666 | −0.672 | −0.791 | −0.887 | −0.827 | −0.475 | −0.648 | −0.095 | −0.259 | −0.469 | −0.391 | −0.488 |
| Aspergillus | −0.018 | −0.959 * | −0.641 | −0.505 | 0.316 | −0.072 | −0.609 | −0.358 | 0.92 | 0.842 | 0.711 | 0.771 | −0.121 | −0.094 | 0.949 | −0.633 | −0.121 | −0.179 | −0.117 |
| Fusarium | −0.451 | 0.572 | 0.621 | −0.343 | −0.517 | 0.38 | 0.689 | 0.443 | −0.185 | −0.068 | −0.029 | −0.138 | −0.381 | −0.191 | −0.254 | −0.041 | −0.387 | −0.421 | −0.37 |
| Dipodascus | −0.486 | 0.243 | −0.502 | 0.974 * | −0.591 | −0.906 | −0.525 | −0.748 | −0.061 | 0.02 | 0.21 | 0.196 | 0.973 * | 0.932 | −0.601 | 0.932 | 0.974 * | 0.986 * | 0.971 * |
| Mucor | −0.011 | −0.962 * | −0.647 | −0.498 | 0.322 | −0.077 | −0.616 | −0.363 | 0.918 | 0.839 | 0.708 | 0.77 | −0.115 | −0.091 | 0.948 | −0.629 | −0.115 | −0.172 | −0.111 |
| Hannaella | −0.449 | 0.571 | 0.622 | −0.345 | −0.515 | 0.382 | 0.69 | 0.445 | −0.185 | −0.068 | −0.03 | −0.139 | −0.383 | −0.194 | −0.253 | −0.043 | −0.389 | −0.423 | −0.373 |
| Sporobolomyces | −0.449 | 0.571 | 0.622 | −0.345 | −0.515 | 0.382 | 0.69 | 0.445 | −0.185 | −0.068 | −0.03 | −0.139 | −0.383 | −0.194 | −0.253 | −0.043 | −0.389 | −0.423 | −0.373 |
| Rhodotorula | −0.011 | −0.962 * | −0.647 | −0.498 | 0.322 | −0.077 | −0.616 | −0.363 | 0.918 | 0.839 | 0.708 | 0.77 | −0.115 | −0.091 | 0.948 | −0.629 | −0.115 | −0.172 | −0.111 |
| Cyberlindnera | 0.946 | 0.148 | 0.526 | −0.132 | 0.784 | 0.601 | 0.45 | 0.666 | −0.672 | −0.791 | −0.887 | −0.827 | −0.475 | −0.648 | −0.095 | −0.259 | −0.469 | −0.391 | −0.488 |
| Trichosporon | −0.469 | 0.714 | 0.01 | 0.898 | −0.706 | −0.595 | −0.006 | −0.339 | −0.472 | −0.351 | −0.14 | −0.2 | 0.729 | 0.726 | −0.921 | 0.984 * | 0.728 | 0.754 | 0.728 |
| Saccharomyces | 0.946 | 0.148 | 0.526 | −0.132 | 0.784 | 0.601 | 0.45 | 0.666 | −0.672 | −0.791 | −0.887 | −0.827 | −0.475 | −0.648 | −0.095 | −0.259 | −0.469 | −0.391 | −0.488 |
| Tausonia | 0.946 | 0.148 | 0.526 | −0.132 | 0.784 | 0.601 | 0.45 | 0.666 | −0.672 | −0.791 | −0.887 | −0.827 | −0.475 | −0.648 | −0.095 | −0.259 | −0.469 | −0.391 | −0.488 |
| Cutaneotrichosporon | −0.781 | 0.615 | −0.102 | 0.758 | −0.929 | −0.665 | −0.082 | −0.466 | −0.181 | −0.024 | 0.192 | 0.107 | 0.733 | 0.814 | −0.769 | 0.911 | 0.729 | 0.719 | 0.738 |
| Pichia | −0.486 | 0.243 | −0.502 | 0.974 * | −0.591 | −0.906 | −0.525 | −0.748 | −0.061 | 0.02 | 0.21 | 0.196 | 0.973 * | 0.932 | −0.601 | 0.932 | 0.974 * | 0.986 * | 0.971 * |
| Plectosphaerella | −0.449 | 0.571 | 0.622 | −0.345 | −0.515 | 0.382 | 0.69 | 0.445 | −0.185 | −0.068 | −0.03 | −0.139 | −0.383 | −0.194 | −0.253 | −0.043 | −0.389 | −0.423 | −0.373 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-H.; Lee, E.-S.; Kim, B.-M.; Oh, M.-H. Potential Correlation between Microbial Diversity and Volatile Flavor Compounds in Different Types of Korean Dry-Fermented Sausages. Foods 2022, 11, 3182. https://doi.org/10.3390/foods11203182
Kim J-H, Lee E-S, Kim B-M, Oh M-H. Potential Correlation between Microbial Diversity and Volatile Flavor Compounds in Different Types of Korean Dry-Fermented Sausages. Foods. 2022; 11(20):3182. https://doi.org/10.3390/foods11203182
Chicago/Turabian StyleKim, Jong-Hui, Eun-Seon Lee, Bu-Min Kim, and Mi-Hwa Oh. 2022. "Potential Correlation between Microbial Diversity and Volatile Flavor Compounds in Different Types of Korean Dry-Fermented Sausages" Foods 11, no. 20: 3182. https://doi.org/10.3390/foods11203182
APA StyleKim, J.-H., Lee, E.-S., Kim, B.-M., & Oh, M.-H. (2022). Potential Correlation between Microbial Diversity and Volatile Flavor Compounds in Different Types of Korean Dry-Fermented Sausages. Foods, 11(20), 3182. https://doi.org/10.3390/foods11203182