
Physiological Response of an Oil-Producing Microalgal Strain to Salinity and Light Stress



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Separation
2.2. Algal Species Culture and Identification
2.2.1. Algal Species Culture
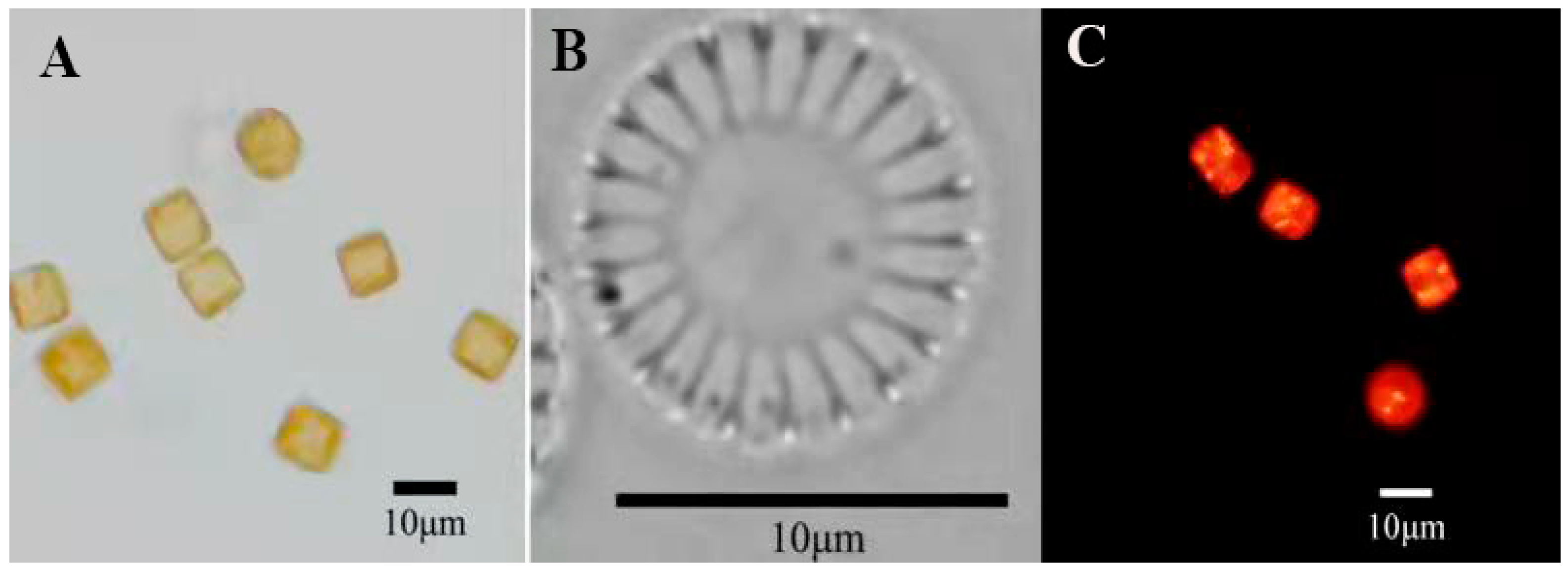
2.2.2. Observation of Algal Cell Morphology and Detection of Oil-Producing Algal Strain
2.2.3. Algal Strain 18S rRNA Gene Amplification and Sequencing
2.3. Determination of Physiological and Biochemical Indicators of Algal Plants
2.3.1. Determination of the Fatty Acid Content
2.3.2. Determination of End Biomass
2.3.3. Determination of Chlorophyll Fluorescence Parameters
2.3.4. Determination of the Chlorophyll Content
2.3.5. Determination of the Total Lipid Content
2.3.6. Statistical Analysis
3. Results and Analysis
3.1. Morphological Identification and Phylogenetic Analysis of Algal Strains
3.2. Fatty Acid Composition Analysis
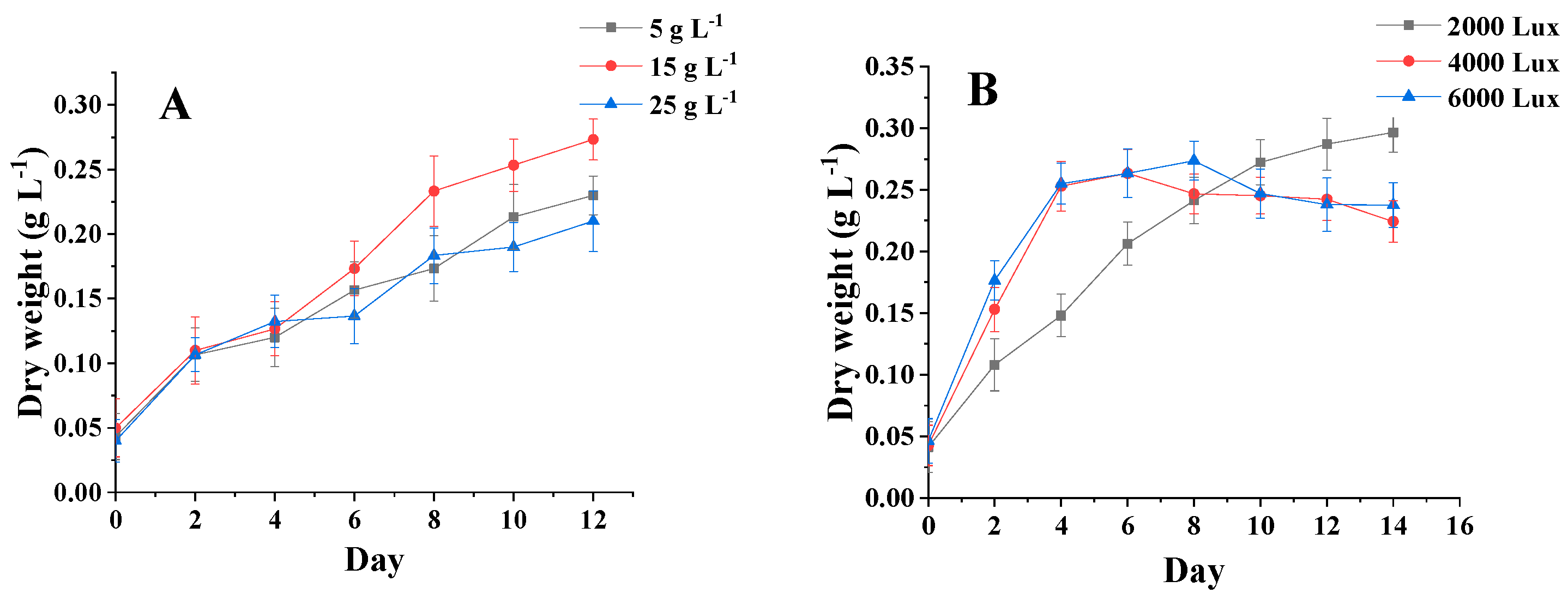
3.3. Analysis of Algal Biomass under Different Salinities and Light Levels
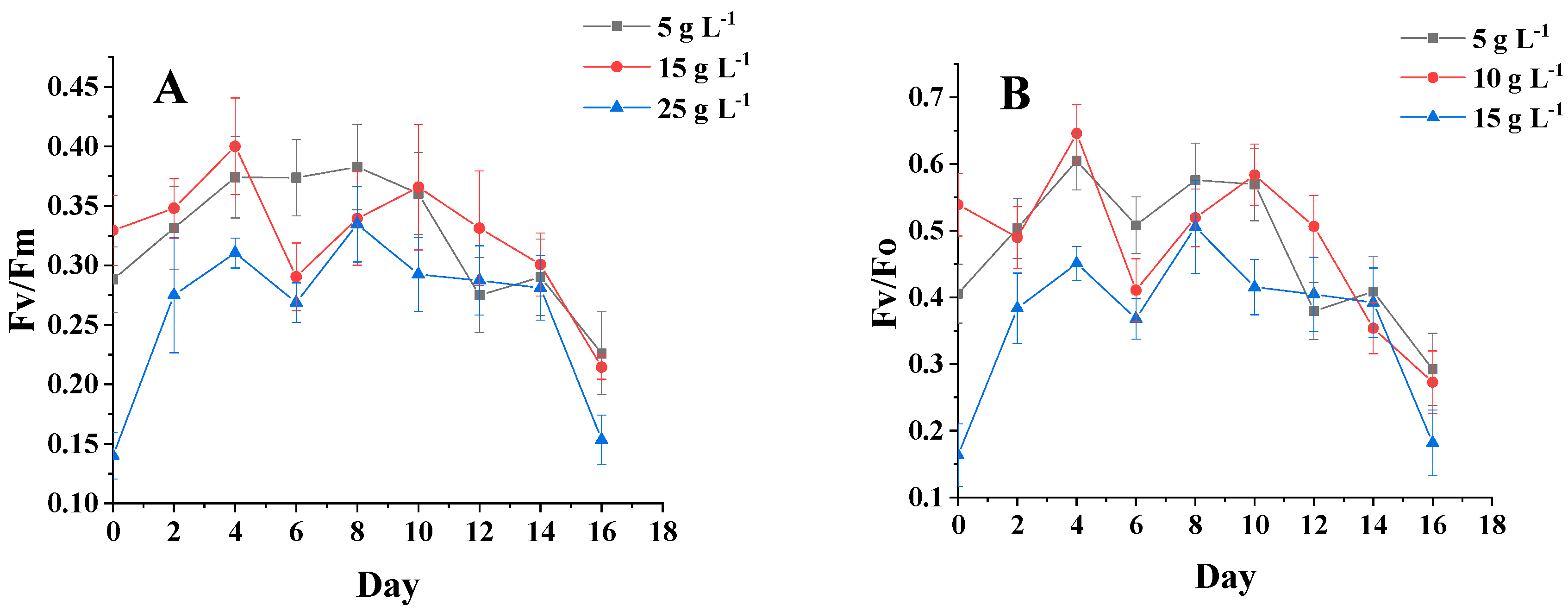
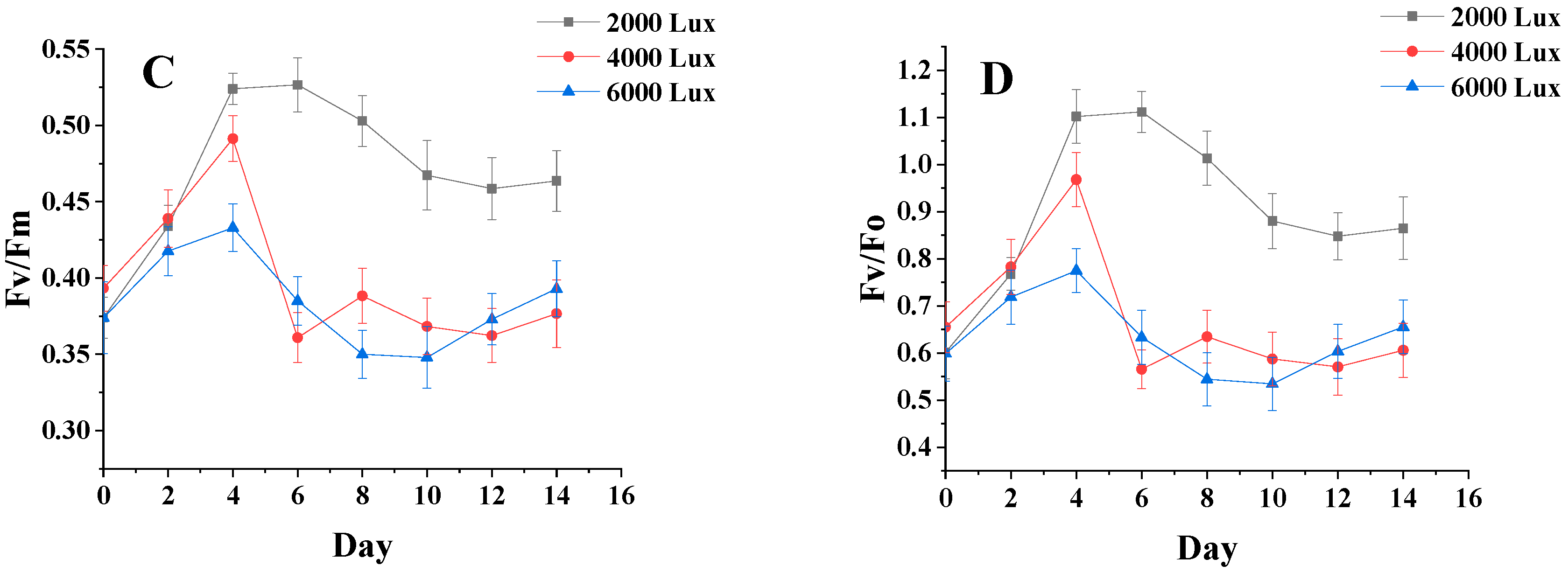
3.4. Analysis of Chlorophyll Fluorescence Parameters of Algal Biomass under Different Salinity and Light Conditions
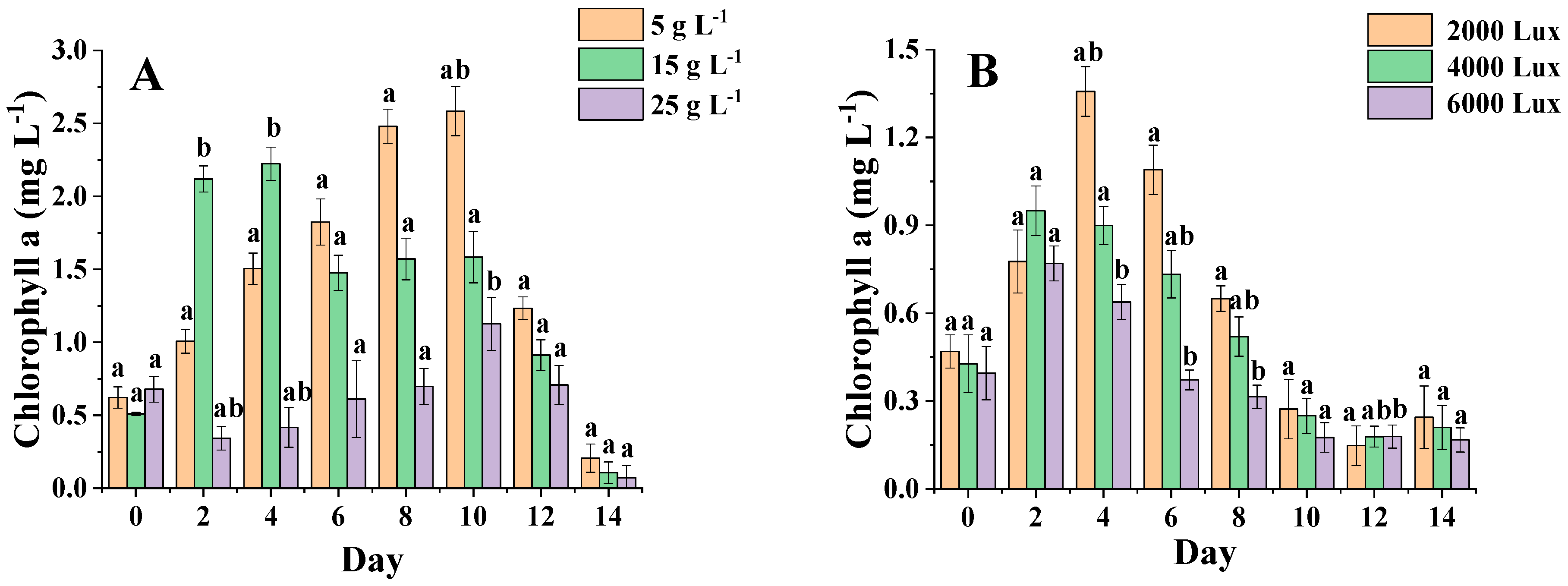
3.5. Analysis of Chlorophyll Content of Algal Biomass under Different Salinities and Light Levels
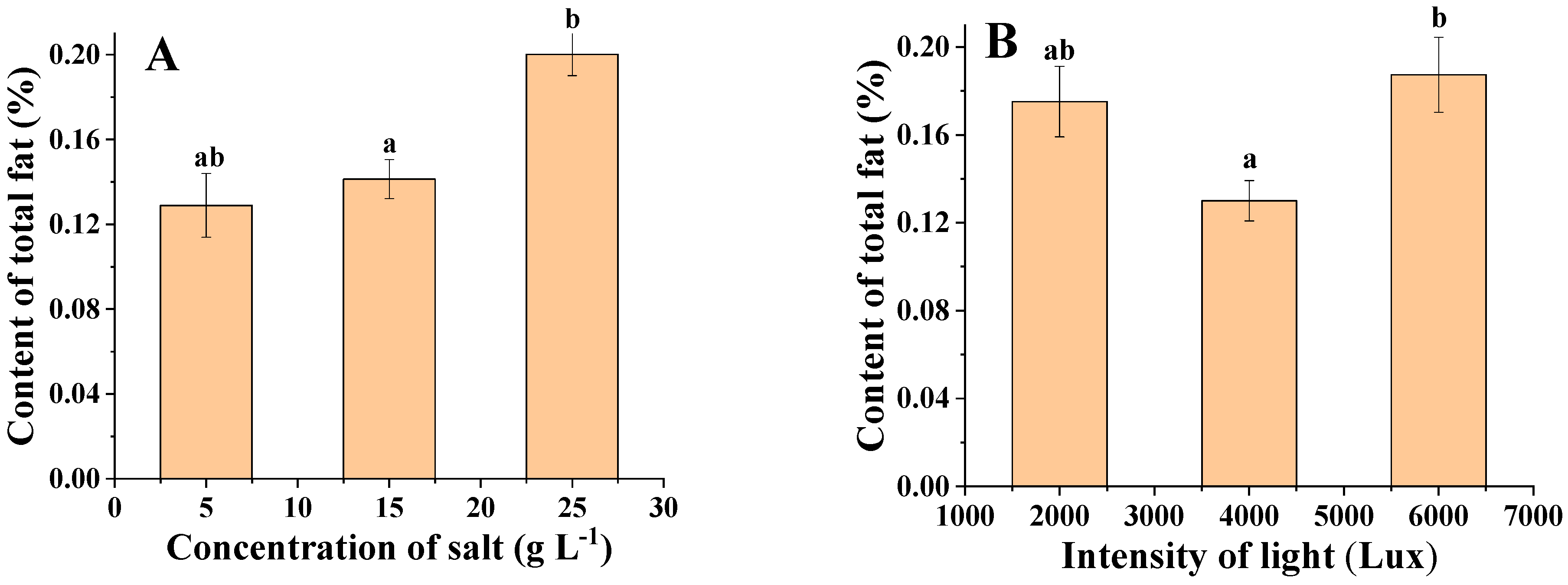
3.6. Analysis of Total Lipid Content of Algal Biomass under Different Salinities and Light Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Yusuf, C. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar]
- Zhang, W.G.; Zhang, D.C.; Chen, X.Y. A novel process for extraction of tea oil from Camellia oleifera seed kernels by combination of microwave puffing and aqueous enzymatic oil extraction. Eur. J. Lipid Sci. Technol. 2012, 114, 352–356. [Google Scholar] [CrossRef]
- Huang, Y.F.; Zhang, D.M.; Xue, S.Z. The Potential of Microalgae Lipids for Edible Oil Production. Appl. Biochem. Biotechnol. 2016, 180, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Lee, J.D.; Hong, I.K. Ultrasonic energy effect on vegetable oil based biodiesel synthetic process. J. Ind. Eng. Chem. 2010, 17, 138–143. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.F.; Lv, J.P.; Feng, J. Morphology, phylogeny and lipid components of an oil-rich microalgal strain. J. Appl. Bot. Food Qual. 2017, 90, 298–305. [Google Scholar]
- Kimura, K.; Yamaoka, M.; Kamisaka, Y. Rapid estimation of lipids in oleaginous fungi and yeasts using Nile red fluorescence. J. Microbiol. Methods 2004, 56, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, P.; Mayer, E.P.; Fowler, S.D. Nile red: A selec-tive fluorescent stain for intracellular lipid droplets. J. Cell Biol. 1985, 100, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Guo, Y.N.; Zhang, X.J. Identification and characterization of a symbiotic alga from soil bryophyte for lipid profiles. Biol. Open 2016, 5, 1317–1323. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Thang, D.V.; Faruq, A.; Thomas-Hall, S.R. High protein and high lipid-producing microalgae from northern Australia as potential feedstock for animal feed and biodiesel. Front. Bioeng. Biotechnol. 2015, 3, 53–60. [Google Scholar]
- Mulbry, W.; Kondrad, S.; Buyer, J. Treatment of dairy and swine manure effluents using freshwater algae: Fatty acid content and composition of algal biomass at different manure loading rates. J. Appl. Phycol. 2008, 20, 1079–1085. [Google Scholar] [CrossRef]
- Liu, J.; Huang, J.; Sun, Z. Differential lipid and fatty acid profiles of photoautotrophic and heterotrophic Chlorella zofingiensis: Assessment of algal oils for biodiesel production. Bioresour. Technol. 2011, 102, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.P.; Guo, J.Y.; Feng, J. Effect of sulfate ions on growth and pollutants removal of self-flocculating microalga Chlorococcum sp. GD in synthetic municipal wastewater. Bioresour. Technol. 2017, 234, 289–296. [Google Scholar] [CrossRef]
- Markou, G.; Depraetere, O.; Muylaert, K. Effect of ammonia on the photosynthetic activity of Arthrospira and Chlorella: A study on chlorophyll fluorescence and electron transport. Algal Res. 2016, 16, 449–457. [Google Scholar] [CrossRef]
- Mera, R.; Torres, E.; Abalde, J. Effects of sodium sulfate on the freshwater microalga Chlamydomonas moewusii: Implications for the optimization of algal culture media. J. Phycol. 2016, 52, 75–88. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W. A rapid method of total lipidextraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Bohnenberger, J.E.; Crossetti, L.O. Influence of temperature and nutrient content on lipid production in freshwater microalgae cultures. An. Acad. Bras. Ciências 2014, 86, 1239–1248. [Google Scholar] [CrossRef] [Green Version]
- Bold, H.C.; Wynne, M.J. Introduction to the algae. Phycologia 1985, 24, 486–487. [Google Scholar]
- Knothe, G. Improving biodiesel fuel properties by modifying fatty ester composition. Energy Environ. Sci. 2009, 2, 759–766. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R. Nutritional evaluation of microalgae oils rich in omega-3 long chain polyunsaturated fatty acids as an alternative for fish oil. Food Chem. 2014, 160, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.M.; Chen, H.; He, C.L. Nitrogen starvation induced oxidative stress in an oil-producing green alga Chlorella sorokiniana C3. PLoS ONE 2013, 8, e69225. [Google Scholar] [CrossRef]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Procssing Technol. 2006, 86, 1059–1070. [Google Scholar] [CrossRef]
- Li, X.L.; Marella, T.K.; Tao, L. Optimization of growth conditions and fatty acid analysis for three freshwater diatom isolates. Phycol. Res. 2017, 65, 177–187. [Google Scholar] [CrossRef]
- Valenzuela-Espinoza, E.; Millán-Núñez, R.; Núñez-Cebrero, F. Protein, carbohydrate, lipid and chlorophyll a content in Isochrysis aff. galbana (clone T-Iso) cultured with a low cost alternative to the f/2 medium. Aquac. Eng. 2002, 25, 207–216. [Google Scholar] [CrossRef]
- Pahl, S.L.; Lewis, D.M.; Chen, F. Heterotrophic growth and nutritional aspects of the diatom Cyclotella cryptica (Bacillariophyceae): Effect of some environmental factors. J. Biosci. Bioeng. 2010, 109, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Bartley, M.L.; Boeing, W.J.; Corcoran, A.A.; Holguin, F.O.; Schaub, T. Effects of salinity on growth and lipid accumulation of biofuel microalga Nannochloropsis salina and invading organisms. Biomass Bioenergy 2013, 54, 83–88. [Google Scholar] [CrossRef]
- Adams, C.; Bugbee, B. Enhancing lipid production of the marine diatom Chaetoceros gracilis: Synergistic interactions of sodium chloride and silicon. J. Appl. Phycol. 2014, 26, 1351–1357. [Google Scholar] [CrossRef]
- Cheirsilp, B.; Torpee, S. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: Effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour. Technol. 2012, 110, 510–516. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Mishra, S. Enhanced biofuel production potential with nutritional stress amelioration through optimization of carbon source and light intensity in Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 179, 565–572. [Google Scholar] [CrossRef]
- Ji, Y.Z.; Liu, Y.P.; Huang, Y.C. Optimization of Cultivation Process for Oil Production of Cyclotella Menni. J. AnHui Agric. Sci. 2012, 40, 6360–6362. [Google Scholar]
- Chen, X.; Goh, Q.Y.; Tan, W. Lumostatic strategy for microalgae cultivation utilizing image analysis and chlorophyll a content as design parameters. Bioresour. Technol. 2011, 102, 6005–6012. [Google Scholar] [CrossRef]
- Wahidin, S.; Idris, A.; Shaleh, S.R. The influence of light intensity and photoperiod on the growth and lipid content of microalgae Nannochloropsis sp. Bioresour. Technol. 2013, 129, 7–11. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Dosage |
|---|---|
| NaNO3 | 0.12 g |
| K2HPO4 | 0.04 g |
| MgSO4·7H2O | 0.07 g |
| KH2PO4 | 0.08 g |
| CaCl2·2H2O | 0.02 g |
| NaSiO3·9H2O | 0.1 g |
| NaCl | 0.01 g |
| MnSO4·4H2O | 0.1 mL |
| Ferric citract | 0.005 g |
| Soil extract | 20 mL |
| A5 solution | 1 mL |
| Distilled water | 979 mL |
| NaNO3 | 0.12 g |
| K2HPO4 | 0.04 g |
| MgSO4·7H2O | 0.07 g |
| Item | Chemical Formula | Content |
|---|---|---|
| C14:0 | C14H28O2 | 7.24% |
| C15:0 | C15H30O2 | 0.37% |
| C16:0 | C16H36O2 | 20.79% |
| cis C16:1 | C16H30O2 | 27.59% |
| cis C16:2 | C16H28O2 | 2.08% |
| cis C16:3 | C16H26O2 | 10.78% |
| cis C18:1 | C18H34O2 | 5.3% |
| cis C20:5 | C20H30O2 | 19.46% |
| Others | - | 6.39% |
| SFA | - | 27.7% |
| MUFA | - | 32.43% |
| PUFA | - | 32.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ju, Z.; Feng, T.; Feng, J.; Lv, J.; Xie, S.; Liu, Q. Physiological Response of an Oil-Producing Microalgal Strain to Salinity and Light Stress. Foods 2022, 11, 215. https://doi.org/10.3390/foods11020215
Ju Z, Feng T, Feng J, Lv J, Xie S, Liu Q. Physiological Response of an Oil-Producing Microalgal Strain to Salinity and Light Stress. Foods. 2022; 11(2):215. https://doi.org/10.3390/foods11020215
Chicago/Turabian StyleJu, Zhihao, Tingting Feng, Jia Feng, Junping Lv, Shulian Xie, and Qi Liu. 2022. "Physiological Response of an Oil-Producing Microalgal Strain to Salinity and Light Stress" Foods 11, no. 2: 215. https://doi.org/10.3390/foods11020215
APA StyleJu, Z., Feng, T., Feng, J., Lv, J., Xie, S., & Liu, Q. (2022). Physiological Response of an Oil-Producing Microalgal Strain to Salinity and Light Stress. Foods, 11(2), 215. https://doi.org/10.3390/foods11020215