
A Comparative Analysis of Lipid Digestion in Human Milk and Infant Formulas Based on Simulated In Vitro Infant Gastrointestinal Digestion




Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemicals
2.3. Methods
2.3.1. Simulated Infant Gastrointestinal Digestion In Vitro
2.3.2. Particle Size Analysis
2.3.3. Lipid Extraction and Analysis
2.3.4. Statistical Analysis
3. Results and Discussion
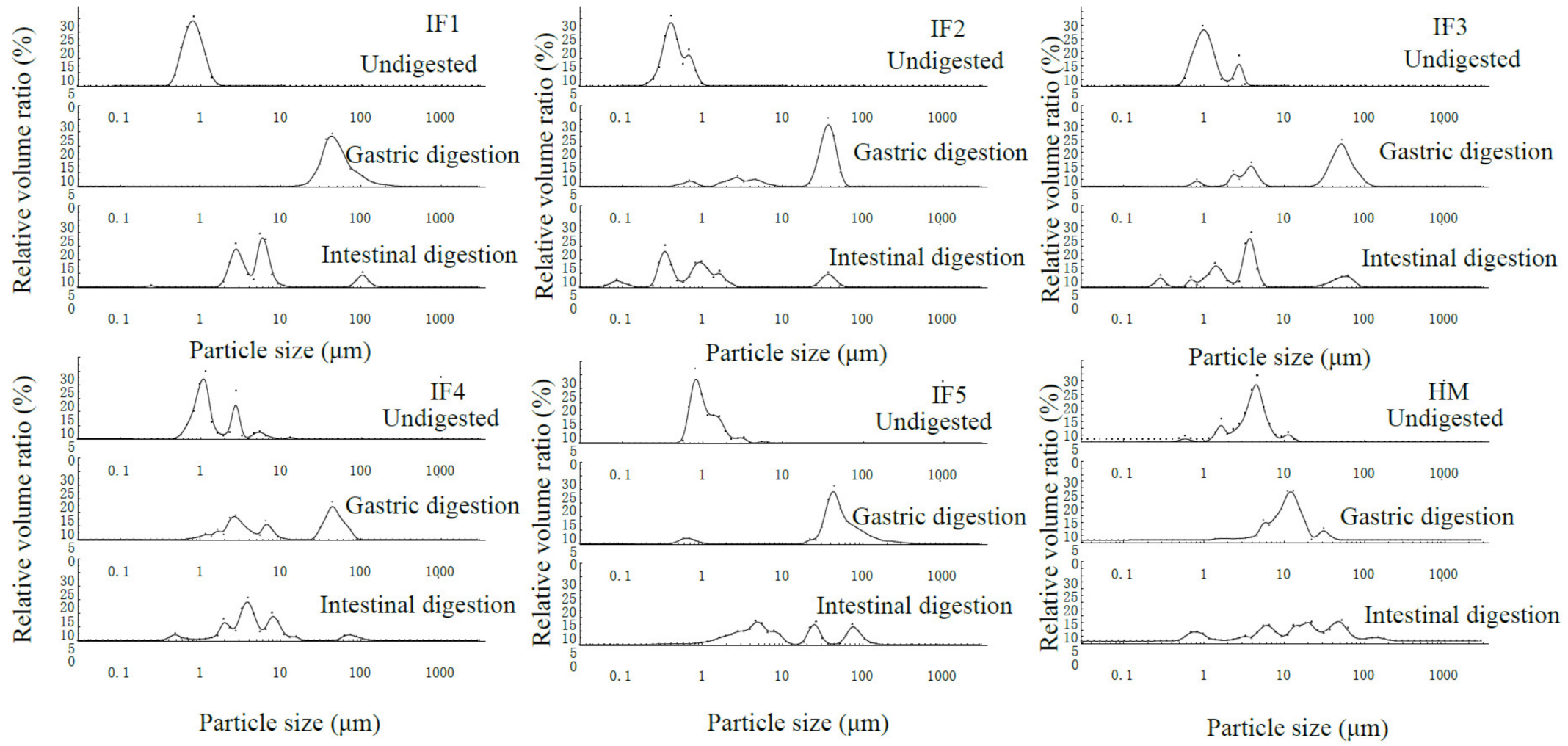
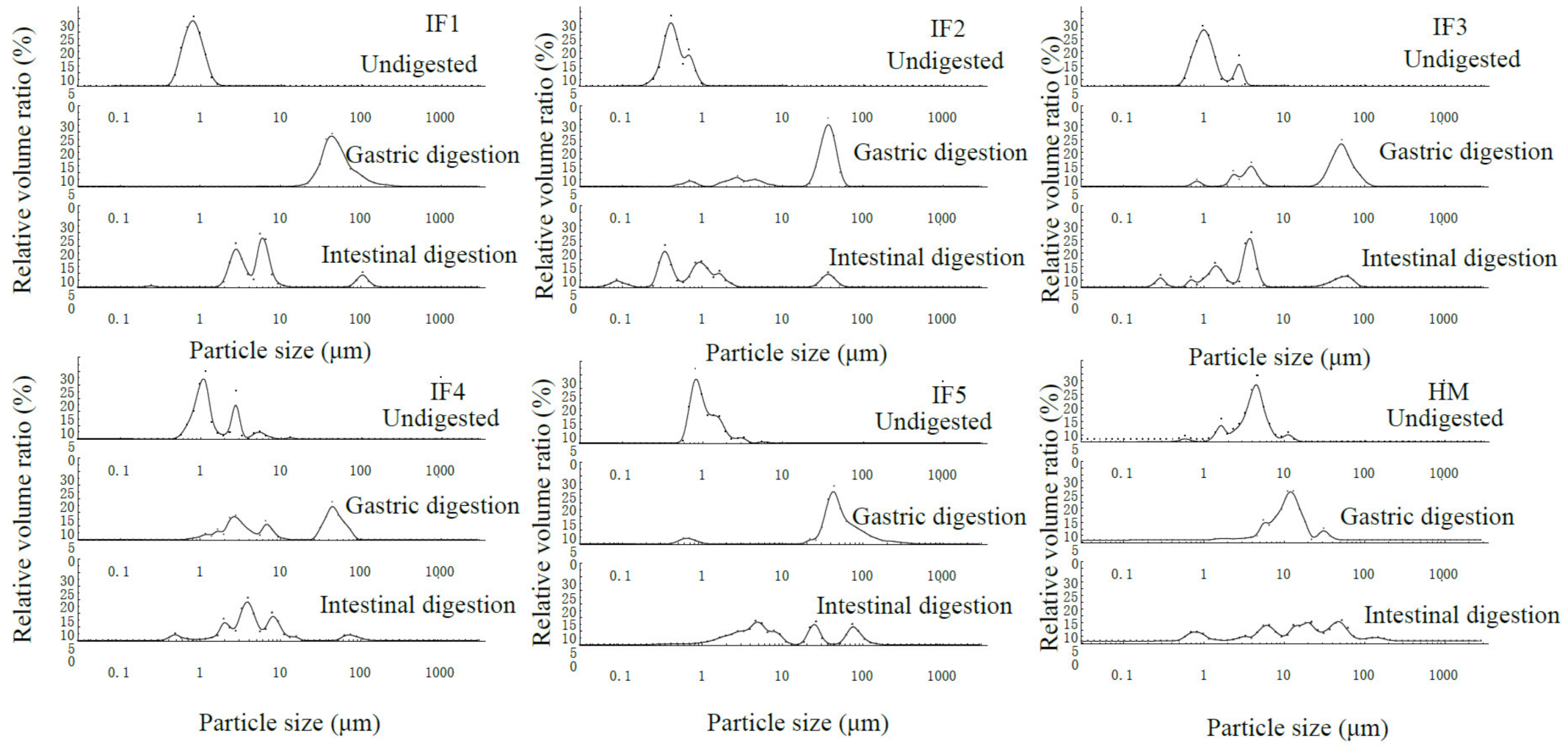
3.1. Initial Fat Globule Particle Size of Infant Formulas and Human Milk
3.2. Fat Globule Particle Size Changes in Infant Formulas and Human Milk during In Vitro Digestion
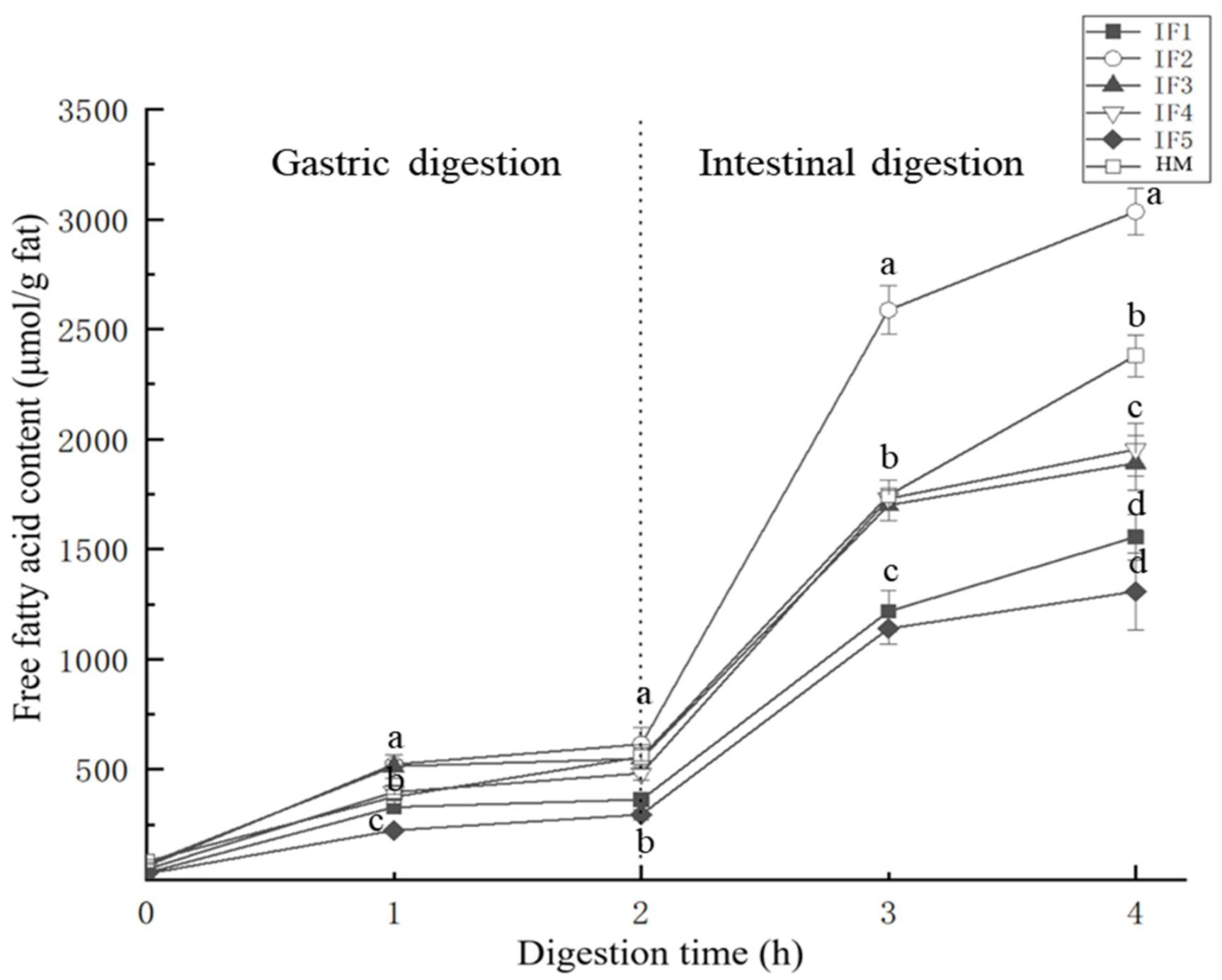
3.3. Fatty Acid Release in Infant Formulas and Human Milk during Digestion
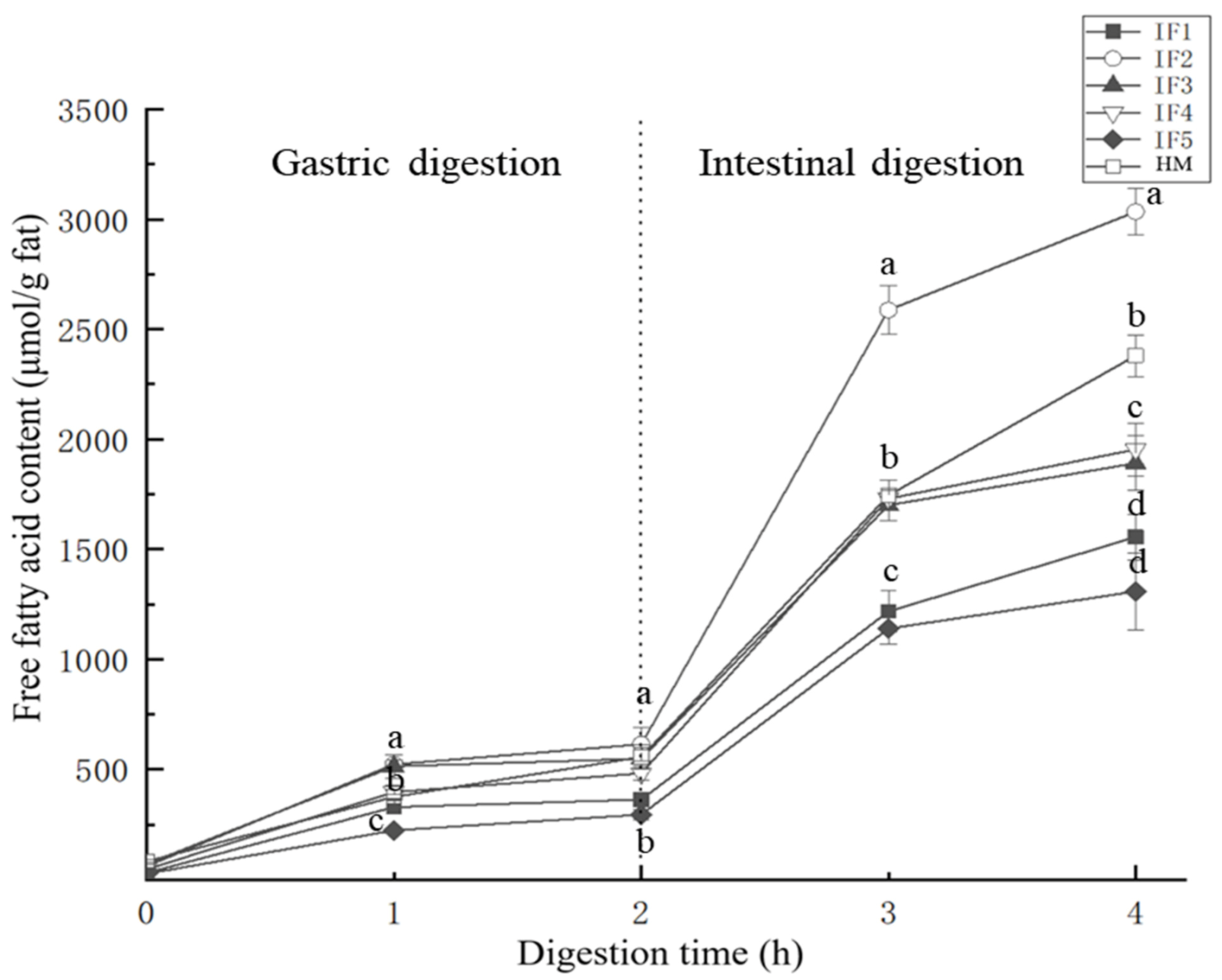
3.3.1. Analysis of Lipolysis Products
3.3.2. Release Rate and Content of Fatty Acids
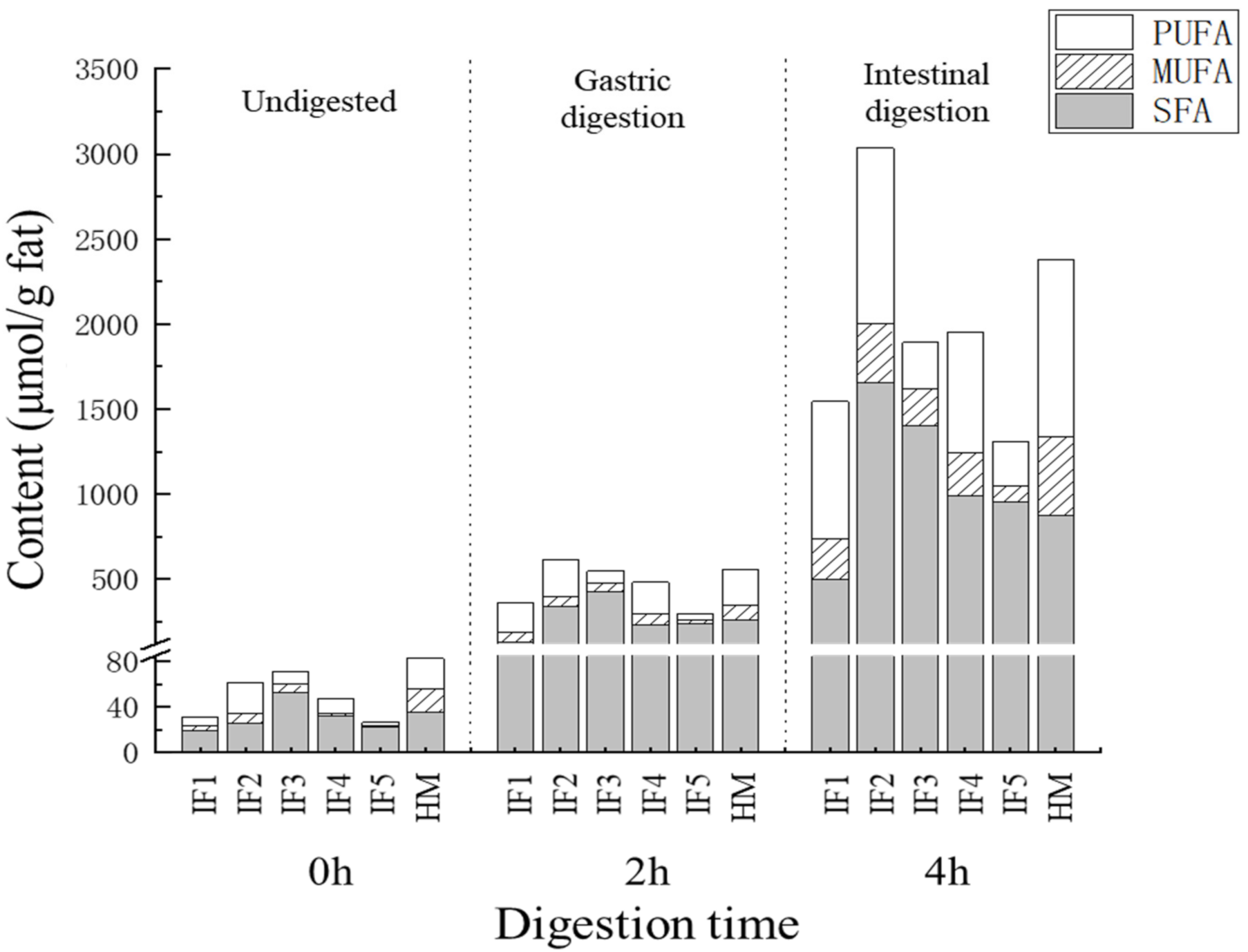
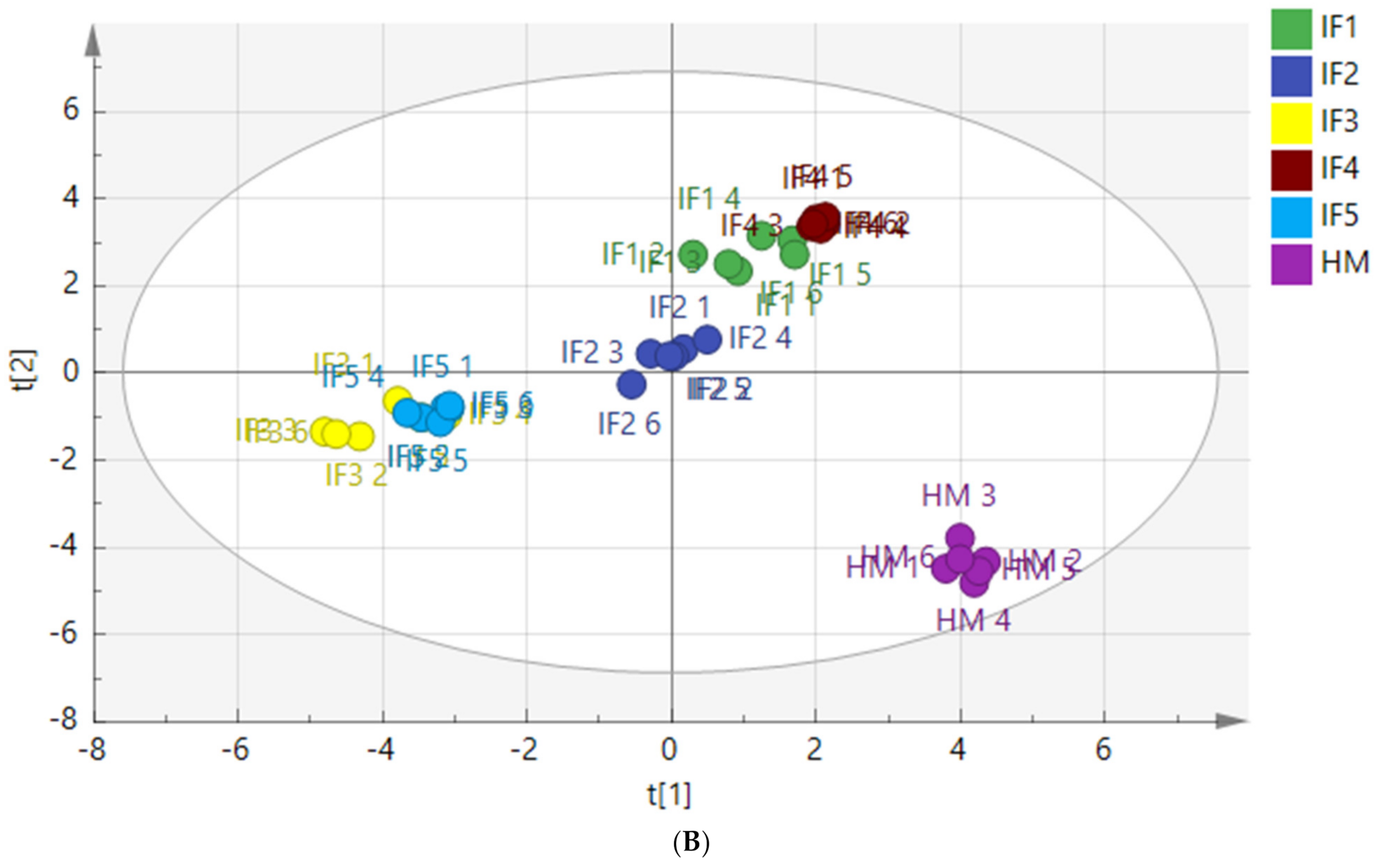
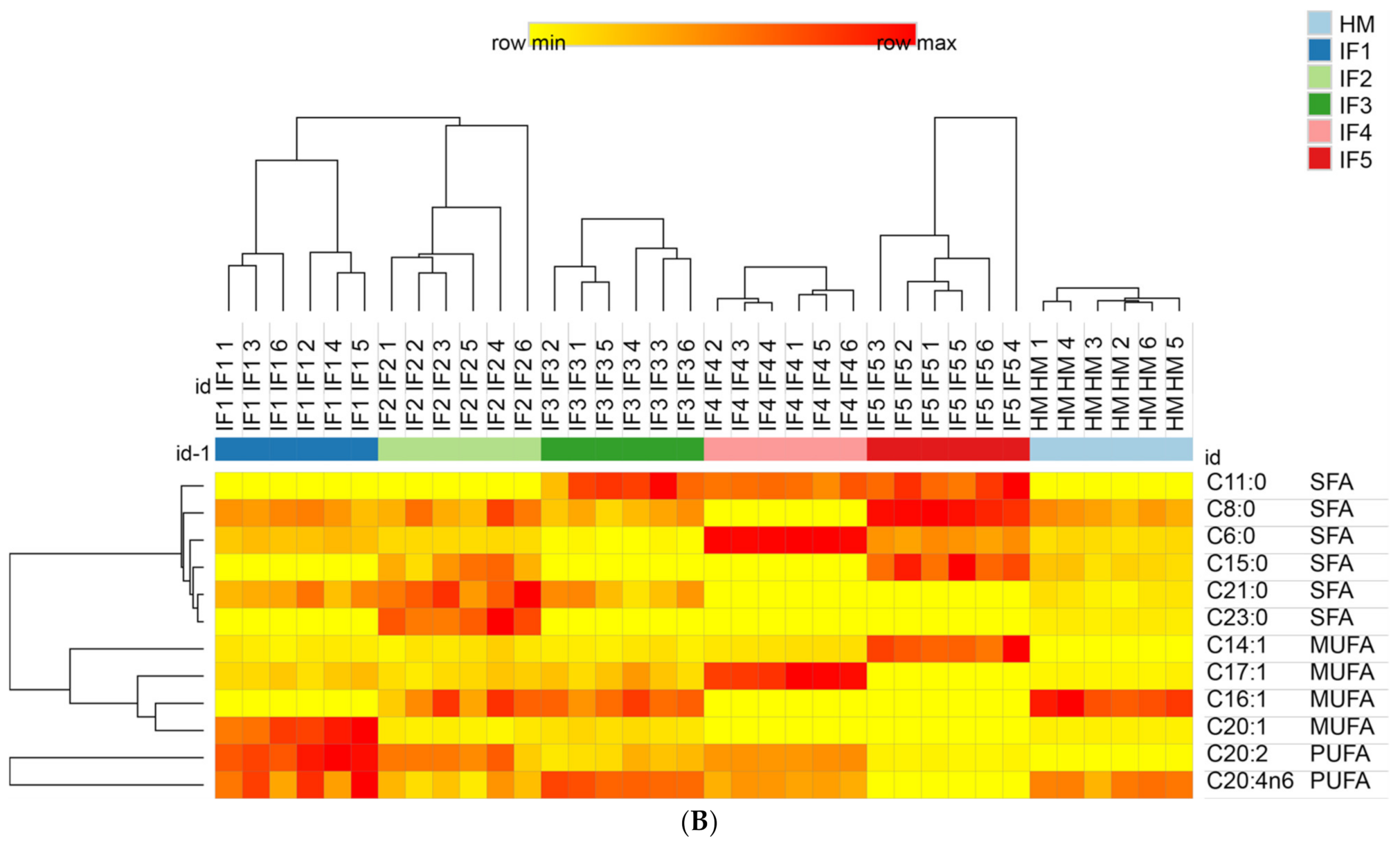
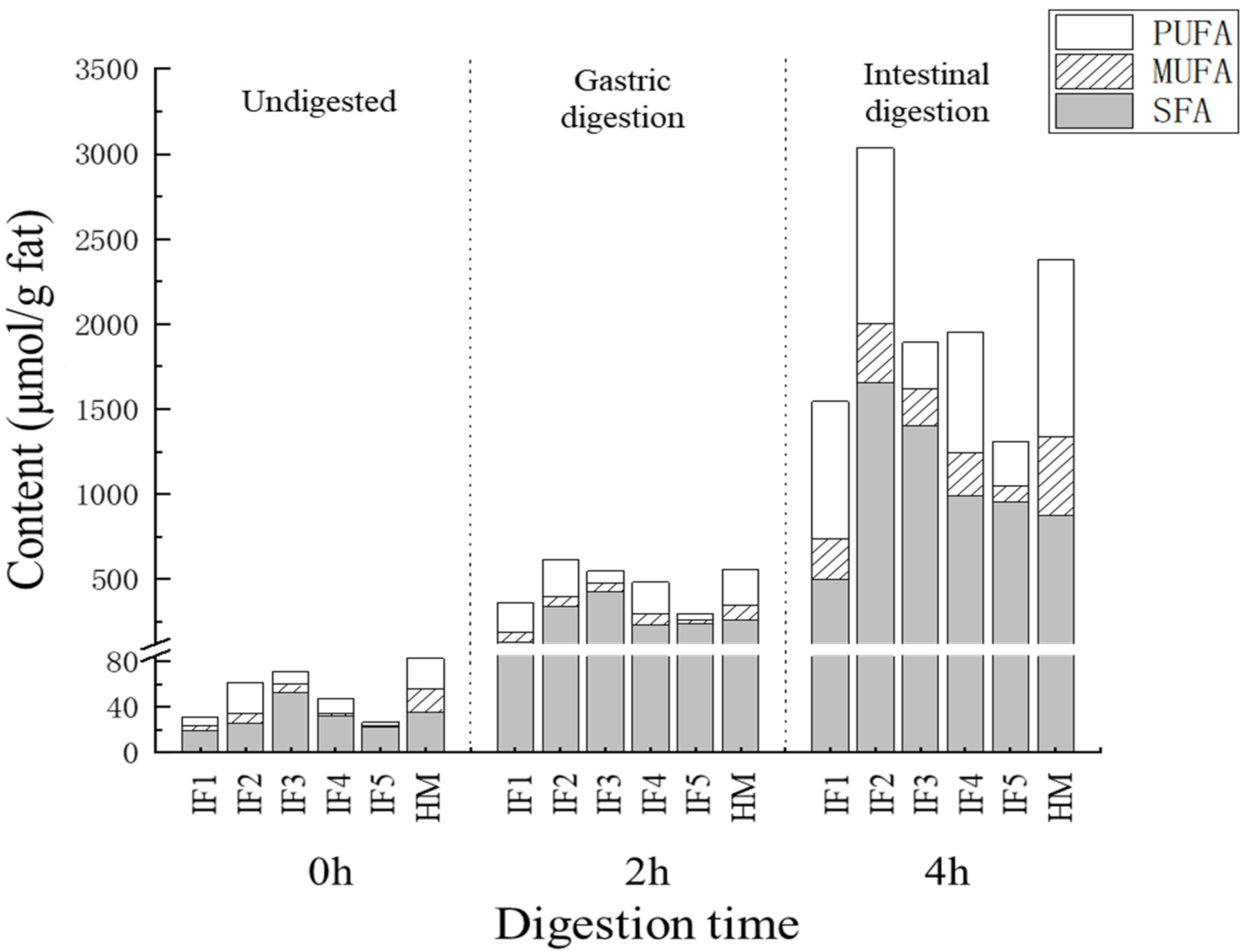
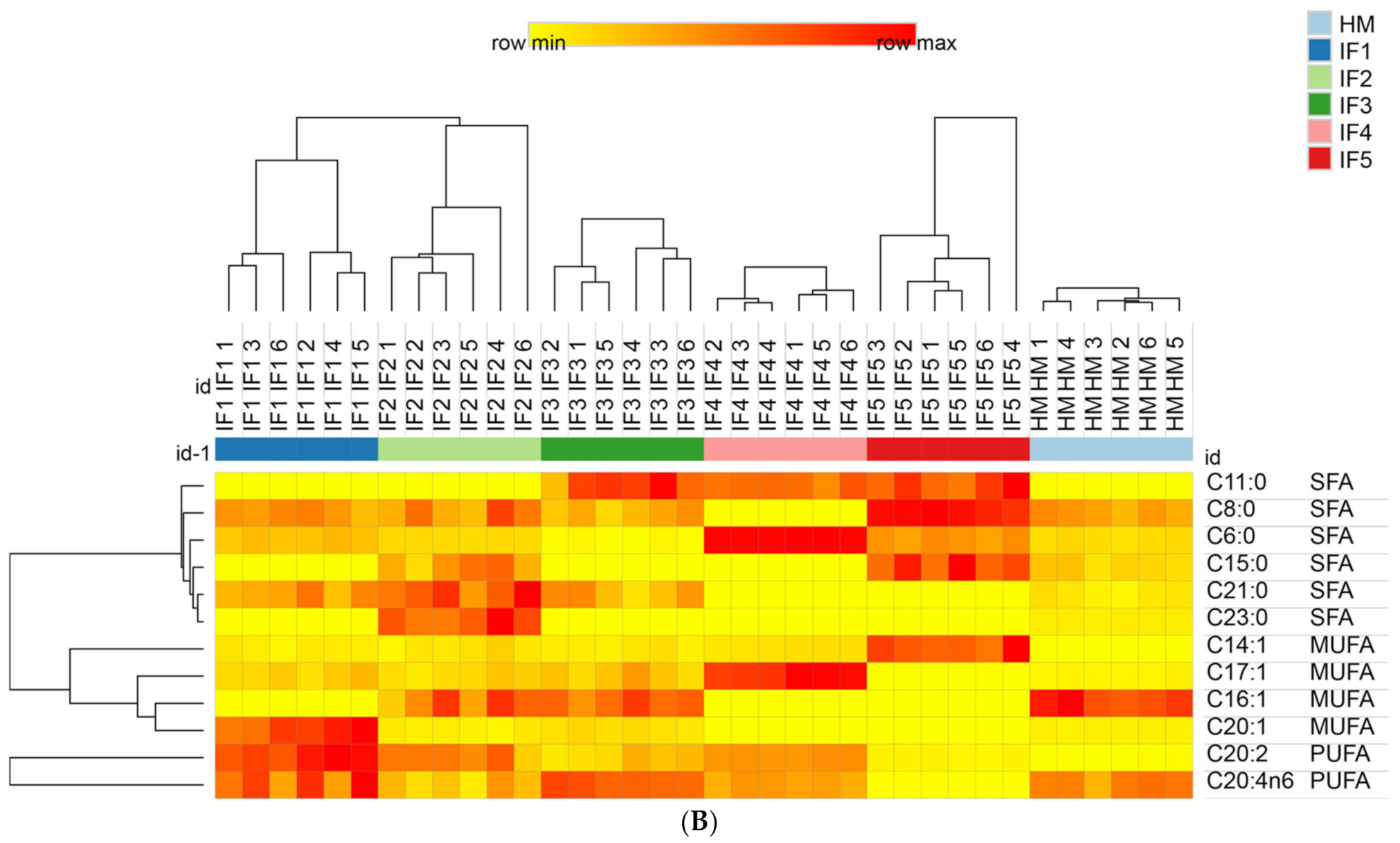
3.3.3. The Content and Proportion of SFAs, MUFAs, and PUFAs
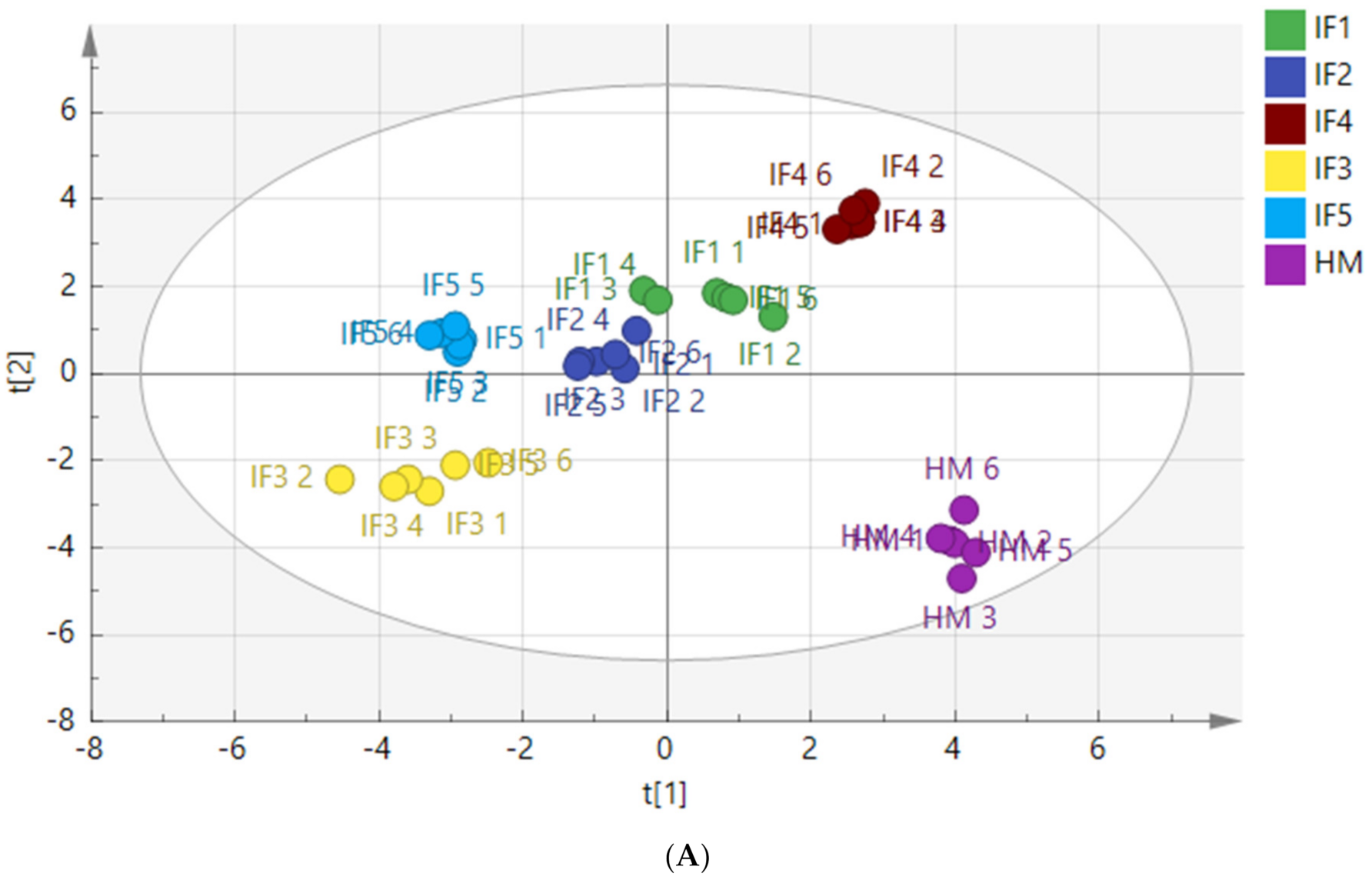
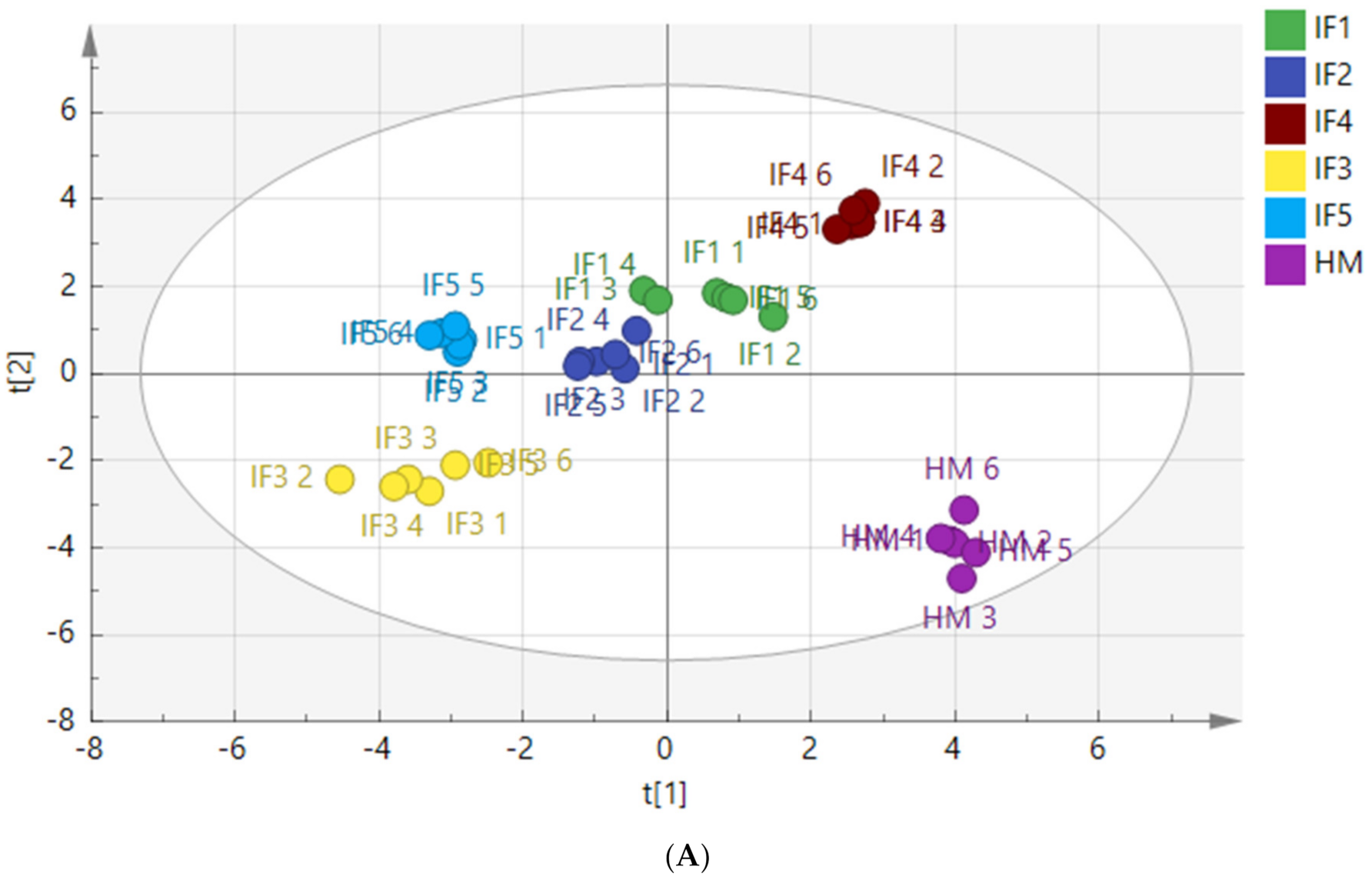
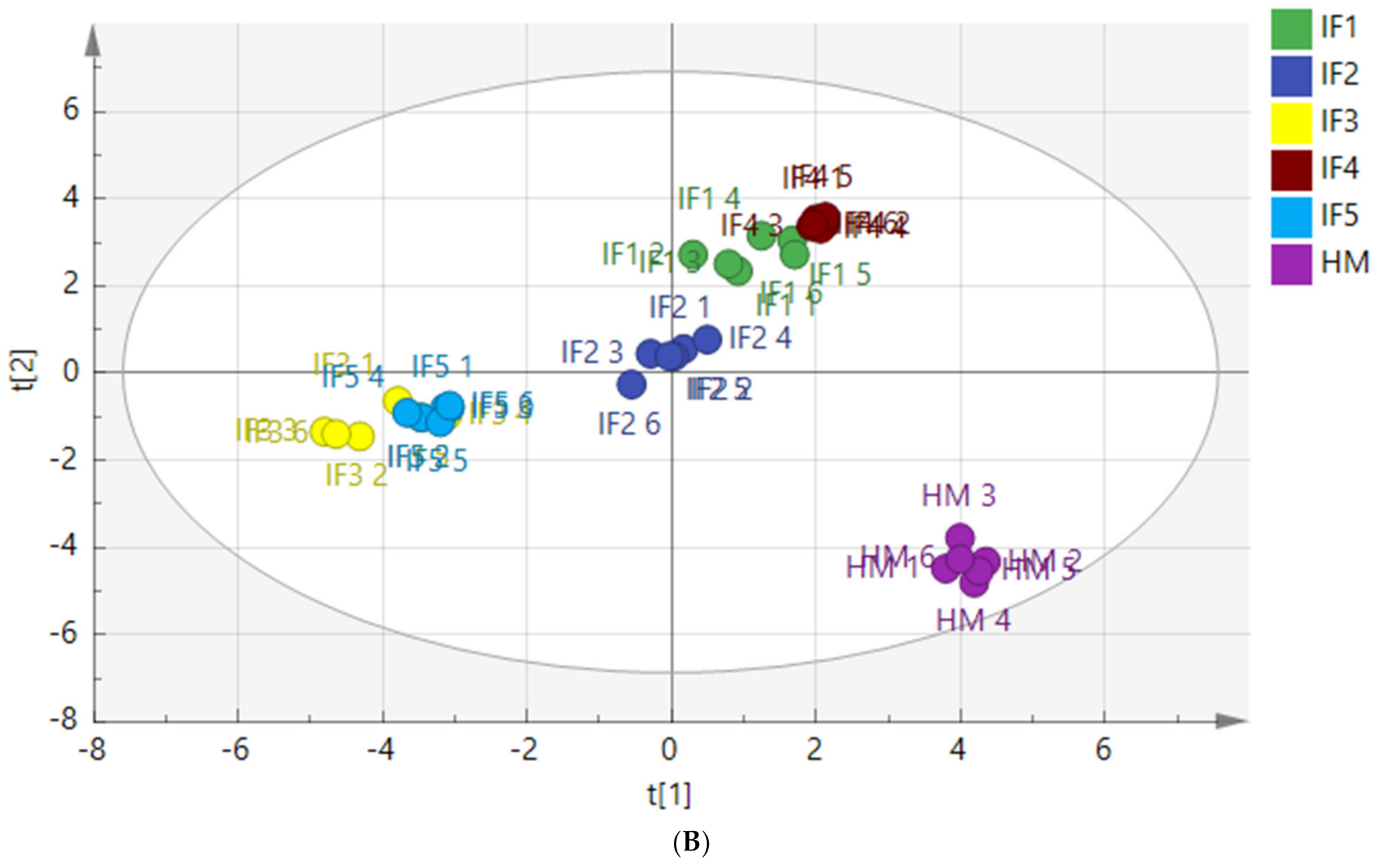
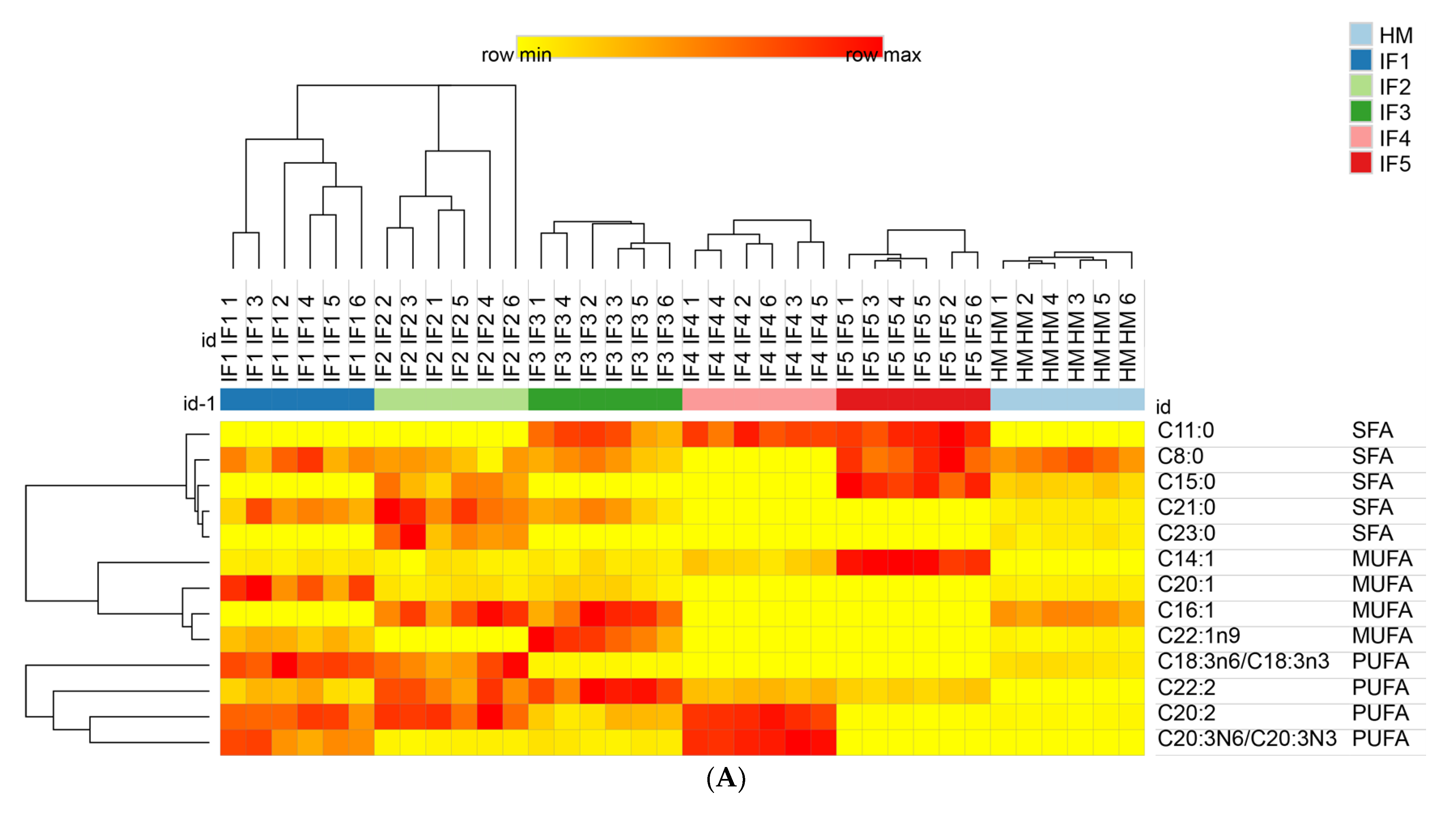
3.3.4. Composition of Free Fatty Acids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Mazzocchi:, A.; D’Oria, V.; De Cosmi, V.; Bettocchi, S.; Milani, G.P.; Silano, M.; Agostoni, C. The Role of Lipids in Human Milk and Infant Formulae. Nutrients 2018, 10, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argov-Argaman, N. Symposium review: Milk fat globule size: Practical implications and metabolic regulation. J. Dairy Sci. 2019, 102, 2783–2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.S.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Development and Function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Claumarchirant, L.; Cilla, A.; Matencio, E.; Sanchez-Siles, L.M.; Castro-Gomez, P.; Fontecha, J.; Alegría, A.; Lagarda, M.J. Addition of milk fat globule membrane as an ingredient of infant formulas for resembling the polar lipids of human milk. Int. Dairy J. 2016, 61, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid Quality in Infant Nutrition: Current Knowledge and Future Opportunities. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M. Human milk: Maternal dietary lipids and infant development. Proc. Nutr. Soc. 2007, 66, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Hageman, J.H.; Danielsen, M.; Nieuwenhuizen, A.; Feitsma, A.L.; Dalsgaard, T.K. Comparison of bovine milk fat and vegetable fat for infant formula: Implications for infant health. Int. Dairy J. 2019, 92, 37–49. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Bhandari, B.; Cichero, J.; Prakash, S. A comprehensive review on in vitro digestion of infant formula. Food Res. Int. 2015, 76, 373–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amara, S.; Patin, A.; Giuffrida, F.; Wooster, T.J.; Thakkar, S.K.; Bénarouche, A.; Poncin, I.; Robert, S.; Point, V.; Molinari, S.; et al. In vitro digestion of citric acid esters of mono- and diglycerides (CITREM) and CITREM-containing infant formula/emulsions. Food Funct. 2014, 5, 1409–1421. [Google Scholar] [CrossRef]
- Lopez, C.; Cauty, C.; Guyomarc’H, F. Organization of lipids in milks, infant milk formulas and various dairy products: Role of technological processes and potential impacts. Dairy Sci. Technol. 2015, 95, 863–893. [Google Scholar] [CrossRef]
- Lopez, C.; Cauty, C.; Guyomarc’H, F. Unraveling the Complexity of Milk Fat Globules to Tailor Bioinspired Emulsions Providing Health Benefits: The Key Role Played by the Biological Membrane. Eur. J. Lipid Sci. Technol. 2018, 121, 1800201. [Google Scholar] [CrossRef] [Green Version]
- Yuan, T.; Zhu, X.-M.; Mu, G.; Qian, F.; Xiong, H. Effects of Chemical Composition and Microstructure in Human Milk and Infant Formulas on Lipid Digestion. J. Agric. Food Chem. 2020, 68, 5462–5470. [Google Scholar] [CrossRef]
- Wei, W.; Jin, Q.; Wang, X. Human milk fat substitutes: Past achievements and current trends. Prog. Lipid Res. 2019, 74, 69–86. [Google Scholar] [CrossRef]
- Berton, A.; Rouvellac, S.; Robert, B.; Rousseau, F.; Lopez, C.; Crenon, I. Effect of the size and interface composition of milk fat globules on their in vitro digestion by the human pancreatic lipase: Native versus homogenized milk fat globules. Food Hydrocoll. 2012, 29, 123–134. [Google Scholar] [CrossRef]
- Yao, Y.; Zhao, G.; Xiang, J.; Zou, X.; Jin, Q.; Wang, X. Lipid composition and structural characteristics of bovine, caprine and human milk fat globules. Int. Dairy J. 2016, 56, 64–73. [Google Scholar] [CrossRef]
- Liang, L.; Zhang, X.; Wang, X.; Jin, Q.; McClements, D.J. Influence of Dairy Emulsifier Type and Lipid Droplet Size on Gastrointestinal Fate of Model Emulsions: In Vitro Digestion Study. J. Agric. Food Chem. 2018, 66, 9761–9769. [Google Scholar] [CrossRef] [PubMed]
- Hageman, J.H.J.; Keijer, J.; Dalsgaard, T.K.; Zeper, L.W.; Carrière, F.; Feitsma, A.L.; Nieuwenhuizen, A.G. Free fatty acid release from vegetable and bovine milk fat-based infant formulas and human milk during two-phase in vitro digestion. Food Funct. 2019, 10, 2102–2113. [Google Scholar] [CrossRef] [PubMed]
- N’Goma, J.-C.B.; Amara, S.; Dridi, K.; Jannin, V.; Carrière, F. Understanding the lipid-digestion processes in the GI tract before designing lipid-based drug-delivery systems. Ther. Deliv. 2012, 3, 105–124. [Google Scholar] [CrossRef]
- Lengsfeld, H.; Beaumier-Gallon, G.; Chahinian, H.; Caro, A.; Verger, R.; Laugier, R.; Carrière, F. Physiology of gastrointestinal lipolysis and therapeutical use of lipases and digestive lipase inhibitors. In Lipases and Phospholipases in Drug Development; Wiley-VCH: Weinheim, Germany, 2005; pp. 195–229. [Google Scholar]
- Bourlieu, C.; Mahdoueni, W.; Paboeuf, G.; Gicquel, E.; Ménard, O.; Pezennec, S.; Bouhallab, S.; Deglaire, A.; Dupont, D.; Carrière, F.; et al. Physico-chemical behaviors of human and bovine milk membrane extracts and their influence on gastric lipase adsorption. Biochimie 2019, 169, 95–105. [Google Scholar] [CrossRef]
- Bourlieu, C.; Ménard, O.; Bouzerzour, K.; Mandalari, G.; Macierzanka, A.; Mackie, A.R.; Dupont, D. Specificity of Infant Digestive Conditions: Some Clues for Developing Relevant In Vitro Models. Crit. Rev. Food Sci. Nutr. 2014, 54, 1427–1457. [Google Scholar] [CrossRef]
- Bourlieu, C.; Ménard, O.; De La Chevasnerie, A.; Sams, L.; Rousseau, F.; Madec, M.-N.; Robert, B.; Deglaire, A.; Pezennec, S.; Bouhallab, S.; et al. The structure of infant formulas impacts their lipolysis, proteolysis and disintegration during in vitro gastric digestion. Food Chem. 2015, 182, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Poquet, L.; Wooster, T.J. Infant digestion physiology and the relevance ofin vitrobiochemical models to test infant formula lipid digestion. Mol. Nutr. Food Res. 2016, 60, 1876–1895. [Google Scholar] [CrossRef] [PubMed]
- Ménard, O.; Bourlieu, C.; De Oliveira, S.; Dellarosa, N.; Laghi, L.; Carrière, F.; Capozzi, F.; Dupont, D.; Deglaire, A. A first step towards a consensus static in vitro model for simulating full-term infant digestion. Food Chem. 2018, 240, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Regueiro, J.; Gibert, J.; Díaz, I. Determination of neutral lipids from subcutaneous fat of cured ham by capillary gas chromatography and liquid chromatography. J. Chromatogr. A 1994, 667, 225–233. [Google Scholar] [CrossRef]
- Lopez, C.; Ménard, O. Human milk fat globules: Polar lipid composition and in situ structural investigations revealing the heterogeneous distribution of proteins and the lateral segregation of sphingomyelin in the biological membrane. Colloids Surf. B Biointerfaces 2011, 83, 29–41. [Google Scholar] [CrossRef]
- Fondaco, D.; Alhasawi, F.; Lan, Y.; Ben-Elazar, S.; Connolly, K.; Rogers, M.A. Biophysical Aspects of Lipid Digestion in Human Breast Milk and Similac™ Infant Formulas. Food Biophys. 2015, 10, 282–291. [Google Scholar] [CrossRef]
- Breij, L.M.; Abrahamse-Berkeveld, M.; Vandenplas, Y.; Jespers, S.N.J.; De Mol, A.C.; Khoo, P.C.; Kalenga, M.; Peeters, S.; Van Beek, R.H.T.; Norbruis, O.F.; et al. An infant formula with large, milk phospholipid-coated lipid droplets containing a mixture of dairy and vegetable lipids supports adequate growth and is well tolerated in healthy, term infants. Am. J. Clin. Nutr. 2019, 109, 586–596. [Google Scholar] [CrossRef]
- Molina, T.L.; Stoll, B.; Mohammad, M.; Mohila, C.A.; Call, L.; Cui, L.; Guthrie, G.; Kunichoff, D.; Lin, S.; Welch-Jernigan, R.; et al. New generation lipid emulsions increase brain DHA and improve body composition, but not short-term neurodevelopment in parenterally-fed preterm piglets. Brain Behav. Immun. 2019, 85, 46–56. [Google Scholar] [CrossRef]
- Luo, J.; Wang, Z.; Li, Y.; Chen, C.; Ren, F.; Guo, H. The simulated in vitro infant gastrointestinal digestion of droplets covered with milk fat globule membrane polar lipids concentrate. J. Dairy Sci. 2019, 102, 2879–2889. [Google Scholar] [CrossRef] [Green Version]
- Garcia, C.; Antona, C.; Robert, B.; Lopez, C.; Armand, M. The size and interfacial composition of milk fat globules are key factors controlling triglycerides bioavailability in simulated human gastro-duodenal digestion. Food Hydrocoll. 2013, 35, 494–504. [Google Scholar] [CrossRef]
- Hernell, O.; Bläckberg, L.; Bernbäck, S. Digestion of Human Milk Fat in Early Infancy. Acta Paediatr. 1989, 78, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, X.; Liu, L.; da Zhang, H.; Zhang, Y.; Chang, Y.H.; Zhu, Q.P. Comparative lipidomics analysis of human, bovine and caprine milk by UHPLC-Q-TOF-MS. Food Chem. 2019, 310, 125865. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Y.; Tang, T.-K.; Lai, O.-M. Health Benefits, Enzymatic Production, and Application of Medium- and Long-Chain Triacylglycerol (MLCT) in Food Industries: A Review. J. Food Sci. 2012, 77, R137–R144. [Google Scholar] [CrossRef]
- Qi, C.; Sun, J.; Xia, Y.; Yu, R.; Wei, W.; Xiang, J.; Jin, Q.; Xiao, H.; Wang, X.-G. Fatty Acid Profile and the sn-2 Position Distribution in Triacylglycerols of Breast Milk during Different Lactation Stages. J. Agric. Food Chem. 2018, 66, 3118–3126. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Digestion Stage | IF1 | IF2 | IF3 | IF4 | IF5 | Human Milk |
|---|---|---|---|---|---|---|
| Undigested | 0.79 ± 0.05 c | 0.46 ± 0.03 c | 1.29 ± 0.12 c | 1.61 ± 0.09 c | 1.09 ± 0.11 c | 4.52 ± 0.37 c |
| After gastric digestion | 56.31 ± 2.66 a | 28.55 ± 2.37 a | 38.33 ± 1.29 a | 24.09 ± 1.33 a | 60.18 ± 3.85 a | 12.48 ± 1.29 b |
| After intestinal digestion | 15.34 ± 0.72 b | 5.16 ± 0.96 b | 11.06 ± 0.63 b | 9.55 ± 0.82 b | 23.42 ± 1.38 b | 27.62 ± 2.27 a |
| Simple | Fat | Undigested | Gastric Digestion | Intestinal Digestion | ||
|---|---|---|---|---|---|---|
| 0 h | 1 h | 2 h | 3 h | 4 h | ||
| HM | TGs | 97.16 ± 4.83 a | 62.10 ± 3.45 b | 55.20 ± 2.99 b | 18.44 ± 2.25 c | 2.62 ± 1.68 d |
| DGs | 2.84 ± 0.42 c | 12.20 ± 1.65 a | 9.57 ± 1.67 b | 8.82 ± 1.86 b | 7.12 ± 1.44 b | |
| MGs | 0 | 6.42 ± 0.97 b | 6.82 ± 1.28 b | 8.78 ± 0.86 a | 4.22 ± 0.87 c | |
| FFAs | 0 | 19.28 ± 2.23 d | 28.41 ± 3.24 c | 63.96 ± 3.98 b | 86.04 ± 4.16 a | |
| IF1 | TGs | 99.38 ± 4.43 a | 63.45 ± 3.52 b | 58.18 ± 2.29 b | 22.56 ± 2.12 c | 5.67 ± 1.55 d |
| DGs | 0.62 ± 0.22 c | 11.64 ± 1.45 a | 10.23 ± 1.78 a | 10.62 ± 2.76 a | 8.42 ± 1.33 b | |
| MGs | 0 | 7.03 ± 1.85 b | 6.37 ± 1.23 b | 7.98 ± 1.67 a | 7.03 ± 0.87 b | |
| FFAs | 0 | 17.88 ± 2.11 d | 25.22 ± 2.24 c | 58.84 ± 2.88 b | 78.88 ± 3.01 a | |
| IF2 | TGs | 99.23 ± 5.13 a | 61.08 ± 3.02 b | 54.78 ± 3.69 b | 16.26 ± 2.22 c | 2.24 ± 1.35 d |
| DGs | 0.77 ± 0.12 d | 11.12 ± 2.15 a | 8.92 ± 1.88 b | 7.84 ± 1.82 b | 6.36 ± 1.23 c | |
| MGs | 0 | 7.78 ± 1.23 a | 7.37 ± 1.57 a | 6.98 ± 1.67 a | 2.73 ± 0.64 b | |
| FFAs | 0 | 20.02 ± 1.11 c | 28.93 ± 2.24 c | 68.92 ± 3.98 b | 88.67 ± 5.31 a | |
| IF3 | TGs | 99.11 ± 5.13 a | 61.42 ± 3.02 b | 58.46 ± 2.66 b | 19.22 ± 2.24 c | 3.45 ± 1.35 d |
| DGs | 0.89 ± 0.42 c | 11.78 ± 3.15 a | 10.43 ± 2.88 b | 10.06 ± 2.43 b | 9.53 ± 2.34 b | |
| MGs | 0 | 6.92 ± 1.54 a | 6.23 ± 1.43 a | 7.00 ± 2.77 a | 4.61 ± 1.64 b | |
| FFAs | 0 | 19.88 ± 1.89 d | 24.88 ± 2.64 c | 63.72 ± 3.98 b | 82.41 ± 4.66 a | |
| IF4 | TGs | 99.02 ± 5.83 a | 61.80 ± 3.45 b | 57.37 ± 2.99 b | 18.72 ± 2.25 c | 3.88 ± 0.68 d |
| DGs | 0.98 ± 0.34 d | 11.84 ± 2.65 a | 9.82 ± 1.67 b | 10.06 ± 2.86 b | 8.74 ± 1.44 c | |
| MGs | 0 | 6.81 ± 1.97 a | 6.78 ± 2.28 a | 7.24 ± 2.86 a | 4.21 ± 0.87 b | |
| FFAs | 0 | 19.55 ± 2.23 d | 26.03 ± 3.24 c | 63.98 ± 3.79 b | 83.17 ± 5.66 a | |
| IF5 | TGs | 98.74 ± 4.32 a | 65.22 ± 3.16 b | 59.24 ± 2.69 b | 23.02 ± 2.12 c | 7.05 ± 1.66 d |
| DGs | 1.26 ± 0.12 c | 12.10 ± 1.15 a | 10.28 ± 1.78 b | 12.62 ± 2.76 a | 12.50 ± 1.43 a | |
| MGs | 0 | 6.26 ± 0.85 b | 6.02 ± 1.23 b | 7.34 ± 0.67 a | 6.22 ± 0.58 b | |
| FFAs | 0 | 16.42 ± 1.11 d | 24.46 ± 1.24 c | 57.02 ± 1.97 b | 74.23 ± 4.01 a | |
| Digestion | Variable Name | VIP Value | p Value |
|---|---|---|---|
| Gastric digestion | C20:1 | 1.32404 | 0.0000054 |
| C23:0 | 1.27437 | 0.0000018 | |
| C18:3n6/C18:3n3 | 1.12550 | 0.0000028 | |
| C16:1 | 1.12003 | 0.0000078 | |
| C15:0 | 1.09888 | 0.0000025 | |
| C22:1n9 | 1.08643 | 0.0000023 | |
| C8:0 | 1.07105 | 0.0000864 | |
| C11:0 | 1.05986 | 0.0000071 | |
| C20:2 | 1.03670 | 0.0000073 | |
| C22:2 | 1.02923 | 0.0000106 | |
| C14:1 | 1.02481 | 0.0000312 | |
| C21:0 | 1.01863 | 0.0000049 | |
| C20:3n6/C20:3n3 | 1.01797 | 0.0000028 | |
| Intestinal digestion | C23:0 | 1.49 | 0.0000018 |
| C20:1 | 1.36 | 0.0000032 | |
| C21:0 | 1.23 | 0.0000051 | |
| C6:0 | 1.14 | 0.0000113 | |
| C17:1 | 1.12 | 0.0000095 | |
| C11:0 | 1.06 | 0.0000080 | |
| C20:4n6 | 1.05 | 0.0000365 | |
| C16:1 | 1.04 | 0.0000071 | |
| C8:0 | 1.03 | 0.0000813 | |
| C20:2 | 1.03 | 0.0000039 | |
| C15:0 | 1.02 | 0.0000031 | |
| C14:1 | 1.00 | 0.0000376 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Lin, S.; Ma, S.; Sun, Y.; Li, X.; Liang, S. A Comparative Analysis of Lipid Digestion in Human Milk and Infant Formulas Based on Simulated In Vitro Infant Gastrointestinal Digestion. Foods 2022, 11, 200. https://doi.org/10.3390/foods11020200
Liu L, Lin S, Ma S, Sun Y, Li X, Liang S. A Comparative Analysis of Lipid Digestion in Human Milk and Infant Formulas Based on Simulated In Vitro Infant Gastrointestinal Digestion. Foods. 2022; 11(2):200. https://doi.org/10.3390/foods11020200
Chicago/Turabian StyleLiu, Lu, Shuang Lin, Shuaiyi Ma, Yue Sun, Xiaodong Li, and Shuyan Liang. 2022. "A Comparative Analysis of Lipid Digestion in Human Milk and Infant Formulas Based on Simulated In Vitro Infant Gastrointestinal Digestion" Foods 11, no. 2: 200. https://doi.org/10.3390/foods11020200
APA StyleLiu, L., Lin, S., Ma, S., Sun, Y., Li, X., & Liang, S. (2022). A Comparative Analysis of Lipid Digestion in Human Milk and Infant Formulas Based on Simulated In Vitro Infant Gastrointestinal Digestion. Foods, 11(2), 200. https://doi.org/10.3390/foods11020200