
Lactiplantibacillus plantarum subsp. plantarum and Fructooligosaccharides Combination Inhibits the Growth, Adhesion, Invasion, and Virulence of Listeria monocytogenes


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Preparation
2.2. Sample Preparation and Inoculation
2.3. In Vitro Virulence Assays
2.4. Analysis of Virulence Factors of L. monocytogenes by Quantitative Real-Time PCR (qRT-PCR)
2.5. Statistical Analysis
3. Results
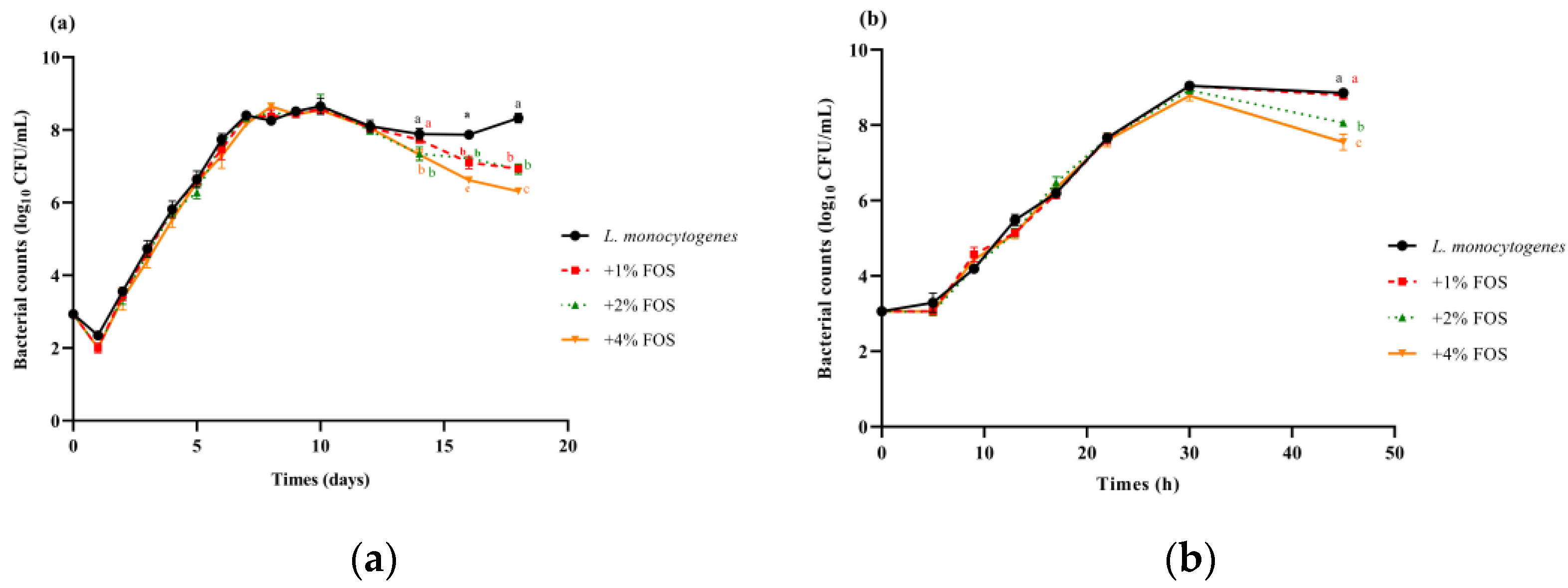
3.1. Effectiveness of FOS Treatment against L. monocytogenes in BHI
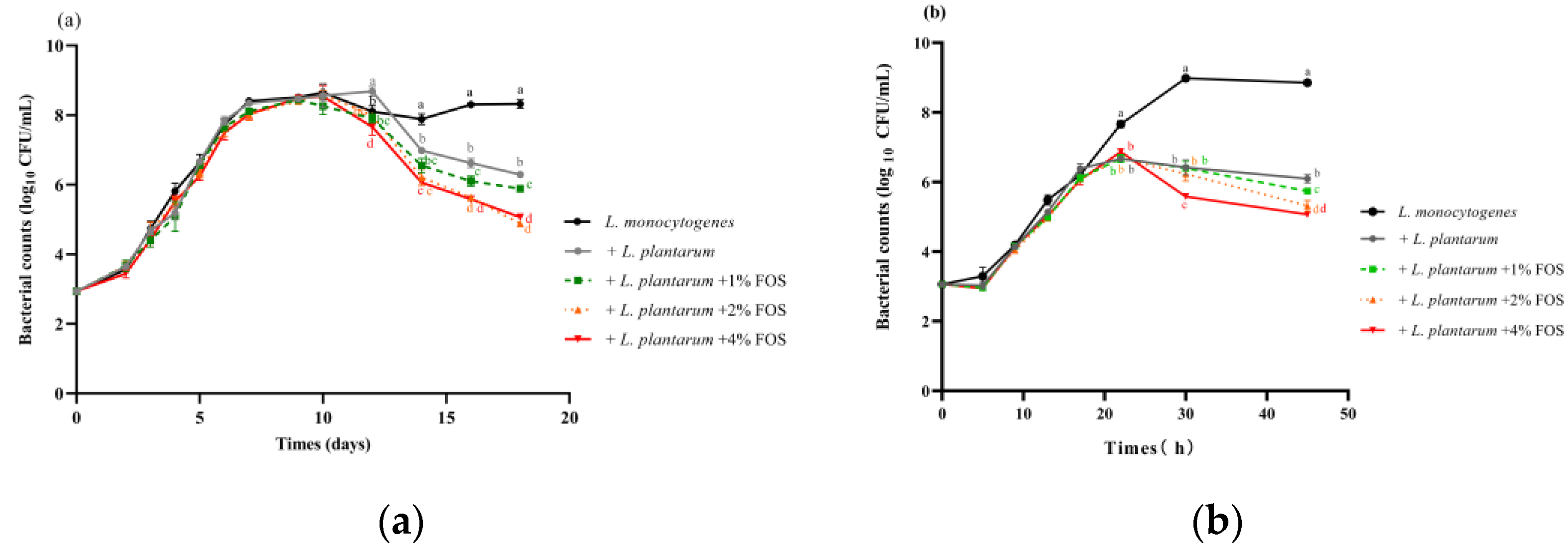
3.2. Effectiveness of L. plantarum and FOS Combination Treatment against L. monocytogenes in BHI
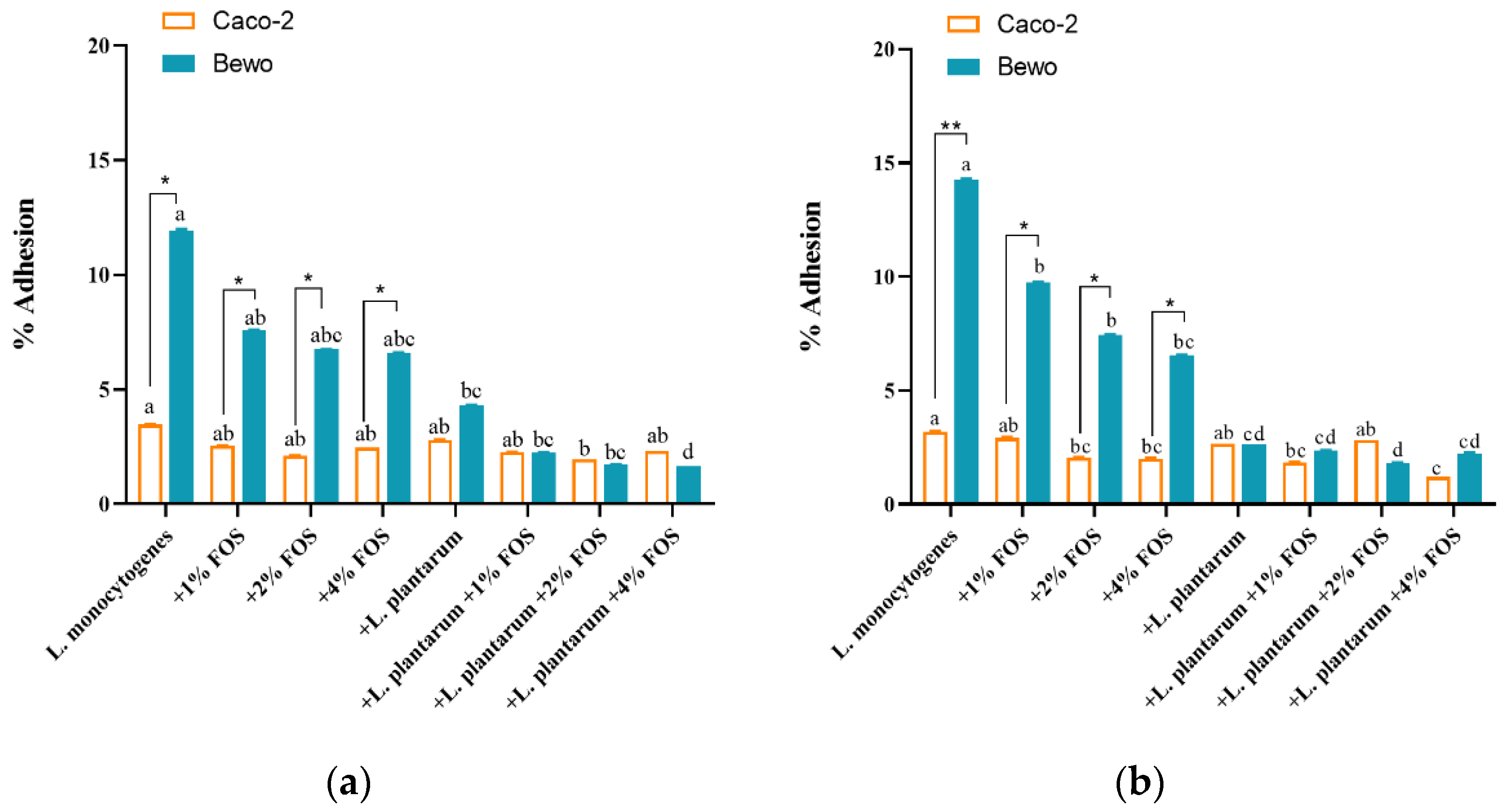
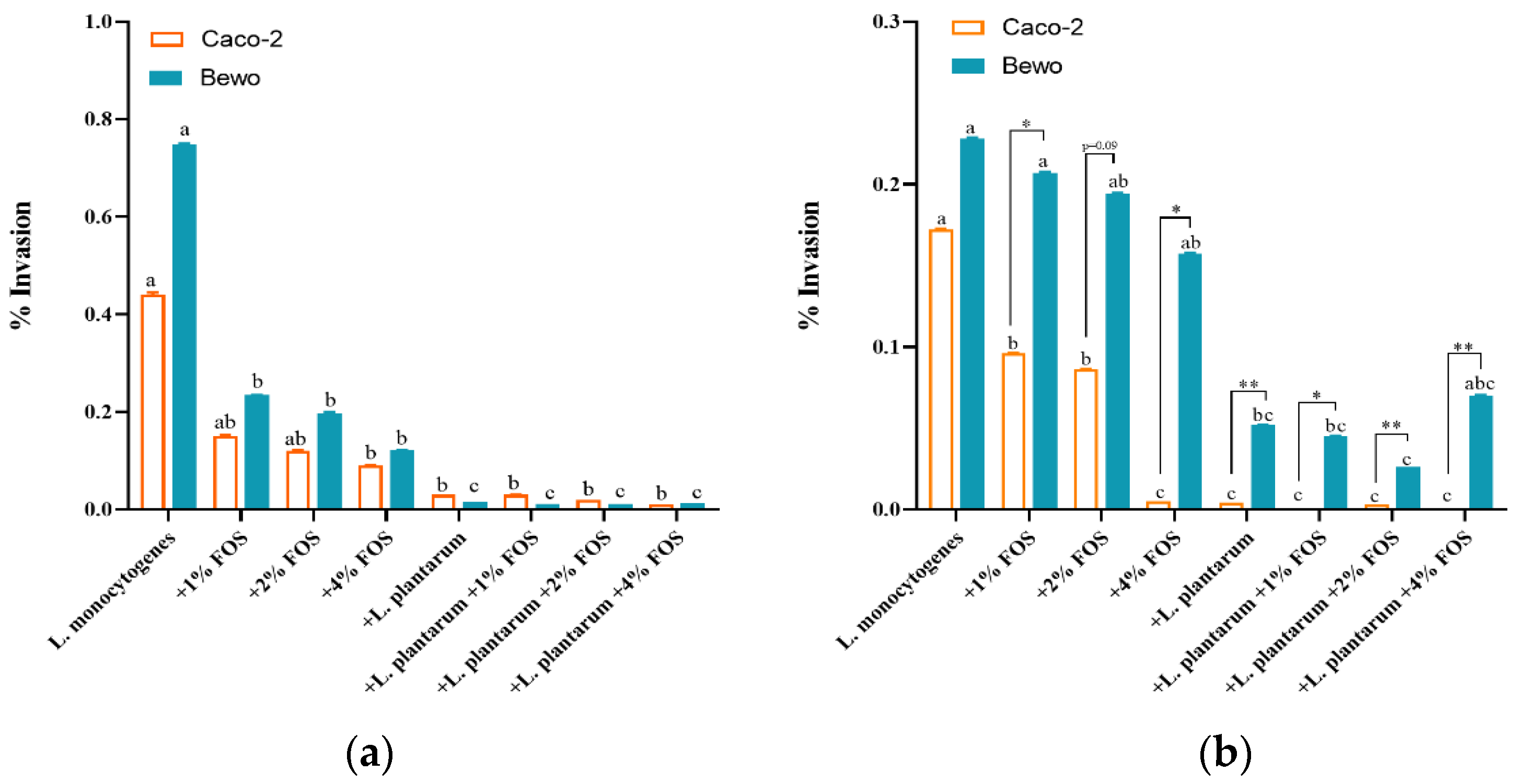
3.3. Ability of Adhesion and Invasion of L. monocytogenes to Caco-2 and BeWo Cells In Vitro
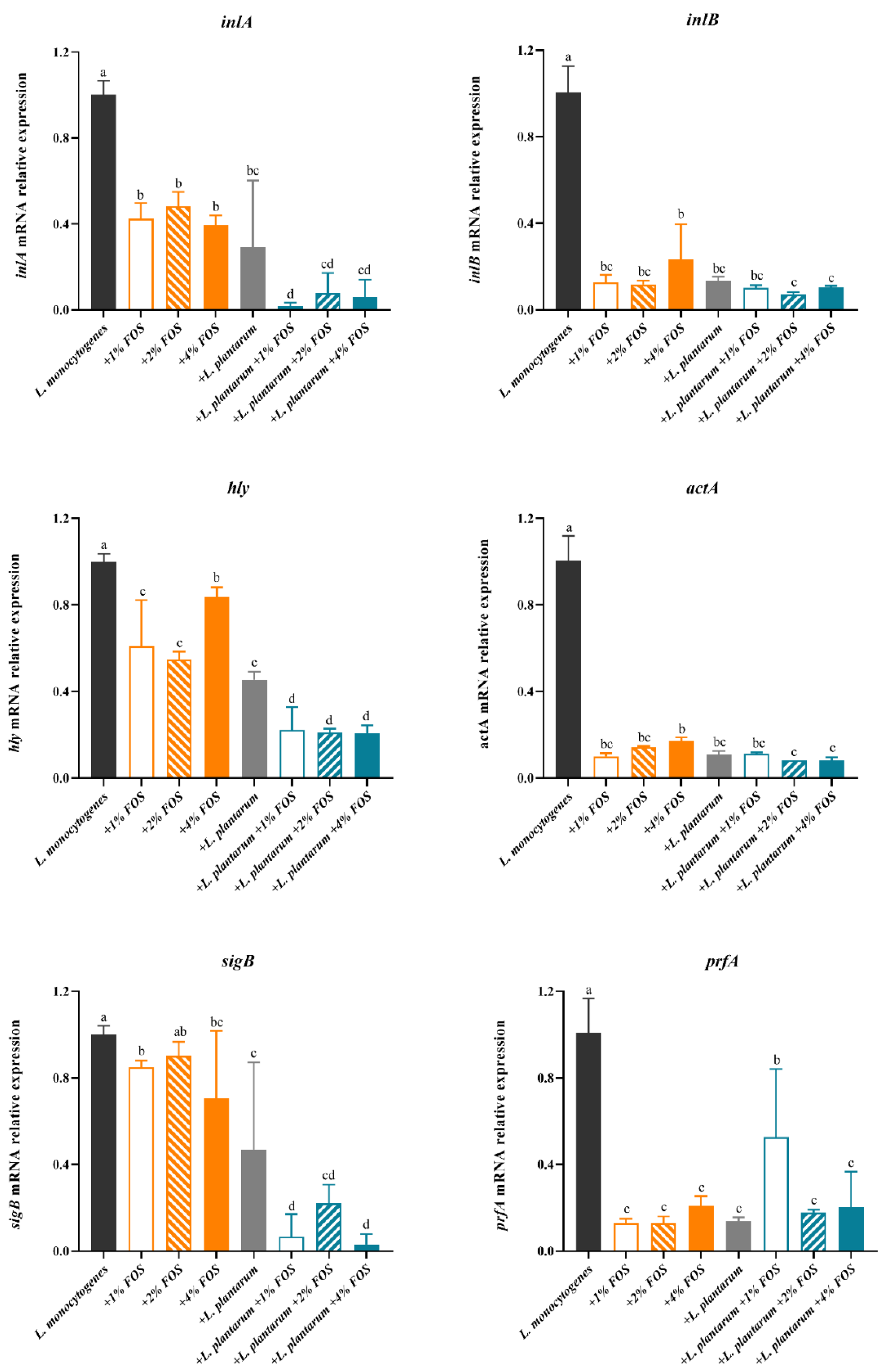
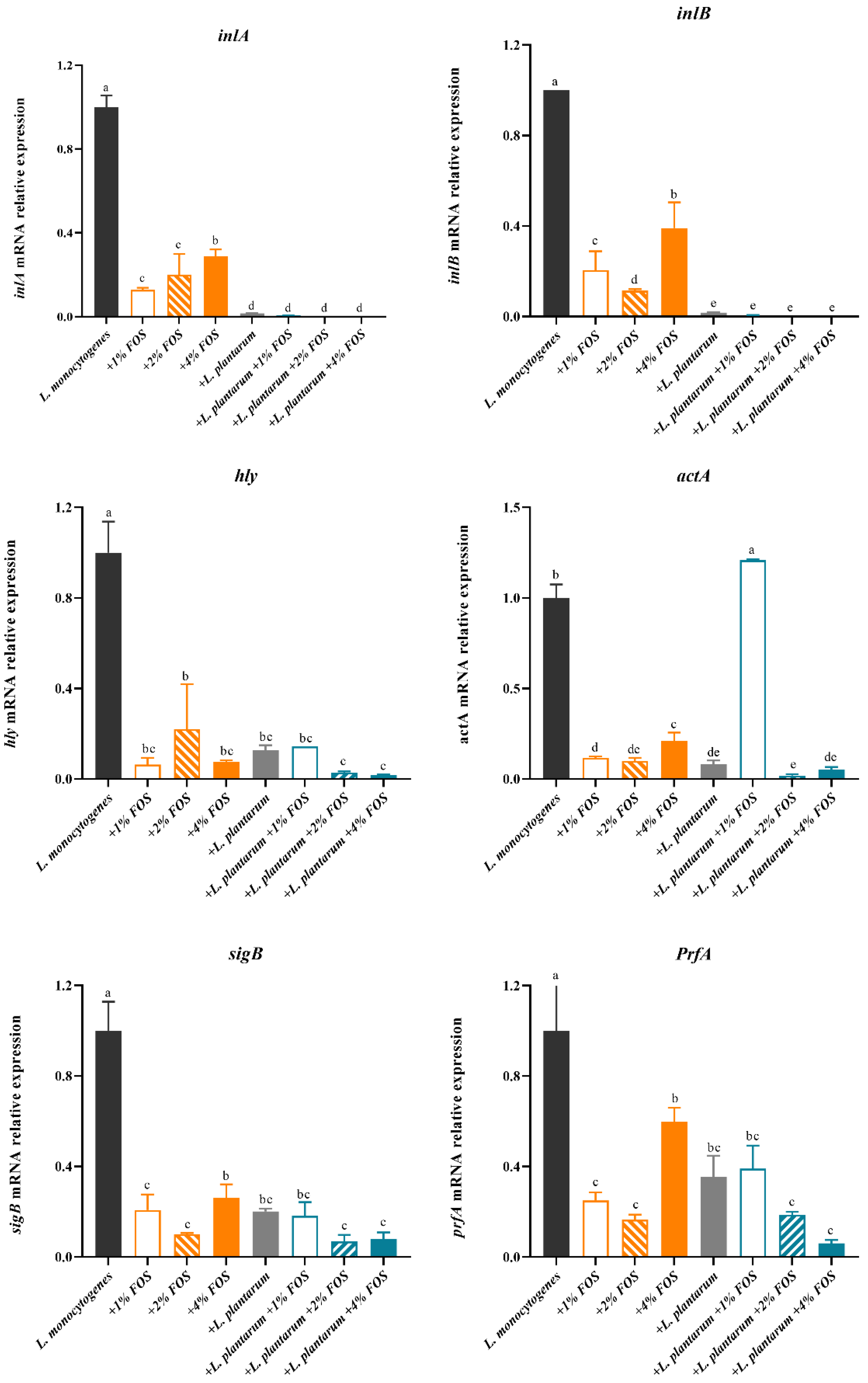
3.4. Virulence Genes Expression of L. monocytogenes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gandhi, M.; Chikindas, M.L. Listeria: A foodborne pathogen that knows how to survive. Int. J. Food Microbiol. 2007, 113, 1–15. [Google Scholar] [CrossRef]
- Oloketuyi, S.F.; Khan, F. Inhibition strategies of Listeria monocytogenes biofilms-current knowledge and future outlooks. J. Basic. Microbiol. 2017, 57, 728–743. [Google Scholar] [CrossRef]
- Thomas, J.; Govender, N.; McCarthy, K.M.; Erasmus, L.K.; Doyle, T.J.; Allam, M.; Ismail, A.; Ramalwa, N.; Sekwadi, P.; Ntshoe, G.; et al. Outbreak of Listeriosis in South Africa Associated with Processed Meat. N. Engl. J. Med. 2020, 382, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.N.; Anyoha, A.; Madoff, L.C.; Lassmann, B. Changing epidemiology of Listeria monocytogenes outbreaks, sporadic cases, and recalls globally: A review of ProMED reports from 1996 to 2018. Int. J. Infect Dis. 2019, 84, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Laine, M.; Wright, A.V.; Vuopio-Varkila, J.; Korhonen, T.; Matila-Sandh, T.M. Development Potential of Selection Criteria in Functional for Probiotic Strains to Assess Their Foods: A Nordic and European Approach. Biosci. Microflora 1996, 15, 61–67. [Google Scholar] [CrossRef][Green Version]
- Bukvicki, D.; Siroli, L.; D’Alessandro, M.; Cosentino, S.; Fliss, I.; Said, L.B.; Hassan, H.; Lanciotti, R.; Patrignani, F. Unravelling the Potential of Lactococcus lactis Strains to Be Used in Cheesemaking Production as Biocontrol Agents. Foods 2020, 9, 1815. [Google Scholar] [CrossRef]
- Park, K.M.; Yoon, S.G.; Choi, T.H.; Kim, H.J.; Park, K.J.; Koo, M. The Bactericidal Effect of a Combination of Food-Grade Compounds and their Application as Alternative Antibacterial Agents for Food Contact Surfaces. Foods 2020, 9, 59. [Google Scholar] [CrossRef]
- Kamiloğlu, A.; Kaban, G.; Kaya, M. Effects of autochthonous Lactobacillus plantarum strains on Listeria monocytogenes in sucuk during ripening. J. Food Saf. 2019, 39. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, X.; Xu, C.; Chen, Y.; Sun, W.; Liu, Q.; Dong, Q. Modeling inhibition effects of Lactobacillus plantarum subsp. plantarum CICC 6257 on growth of Listeria monocytogenes in ground pork stored at CO2-rich atmospheres. LWT-Food Sci. Technol. 2018, 97, 811–817. [Google Scholar] [CrossRef]
- Schillinger, U.; Kaya, M.; Lücke, F.K. Behaviour of Listerla monocytogenes in meat and its control by a bacteriocin-producing strain of Lactobacillus sake. J. Appl. Bacteriol. 1991, 70, 473–478. [Google Scholar] [CrossRef] [PubMed]
- RodrÍGuez, E.V.A.; Arqués, J.L.; Gaya, P.; Nuñez, M.; Medina, M. Control of Listeria monocytogenes by bacteriocins and monitoring of bacteriocin-producing lactic acid bacteria by colony hybridization in semi-hard raw milk cheese. J. Dairy Res. 2001, 68, 131–137. [Google Scholar] [CrossRef]
- Aljasir, S.F.; Gensler, C.; Sun, L.; D’Amico, D.J. The efficacy of individual and combined commercial protective cultures against Listeria monocytogenes, Salmonella, O157 and non-O157 shiga toxin-producing Escherichia coli in growth medium and raw milk. Food Control 2020, 109, 106924. [Google Scholar] [CrossRef]
- Beristain-Bauza, S.C.; Mani-López, E.; Palou, E.; López-Malo, A. Antimicrobial activity and physical properties of protein films added with cell-free supernatant of Lactobacillus rhamnosus. Food Control 2016, 62, 44–51. [Google Scholar] [CrossRef]
- Huang, Y.; Ye, K.; Yu, K.; Wang, K.; Zhou, G. The potential influence of two Enterococcus faecium on the growth of Listeria monocytogenes. Food Control 2016, 67, 18–24. [Google Scholar] [CrossRef]
- Dong, Q.; Zhang, W.; Guo, L.; Niu, H.; Liu, Q.; Wang, X. Influence of Lactobacillus plantarum individually and in combination with low O2-MAP on the pathogenic potential of Listeria monocytogenes in cabbage. Food Control 2020, 107, 106765. [Google Scholar] [CrossRef]
- Upadhyay, A.; Upadhyaya, I.; Mooyottu, S.; Venkitanarayanan, K. Eugenol in combination with lactic acid bacteria attenuates Listeria monocytogenes virulence in vitro and in invertebrate model Galleria mellonella. J. Med. Microbiol. 2016, 65, 443–455. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Speranza, B.; Campaniello, D.; Monacis, N.; Bevilacqua, A.; Sinigaglia, M.; Corbo, M.R. Functional cream cheese supplemented with Bifidobacterium animalis subsp. lactis DSM 10140 and Lactobacillus reuteri DSM 20016 and prebiotics. Food Microbiol. 2018, 72, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Adebola, O.O.; Corcoran, O.; Morgan, W.A. Synbiotics: The impact of potential prebiotics inulin, lactulose and lactobionic acid on the survival and growth of lactobacilli probiotics. J. Funct. Foods 2014, 10, 75–84. [Google Scholar] [CrossRef]
- Pranckute, R.; Kaunietis, A.; Kuisiene, N.; Citavicius, D.J. Combining prebiotics with probiotic bacteria can enhance bacterial growth and secretion of bacteriocins. Int. J. Biol. Macromol. 2016, 89, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Bengoa, A.A.; Dardis, C.; Garrote, G.L.; Abraham, A.G. Health-Promoting Properties of Lacticaseibacillus paracasei: A Focus on Kefir Isolates and Exopolysaccharide-Producing Strains. Foods 2021, 10, 2239. [Google Scholar] [CrossRef]
- Mutanda, T.; Mokoena, M.P.; Olaniran, A.O.; Wilhelmi, B.S.; Whiteley, C.G. Microbial enzymatic production and applications of short-chain fructooligosaccharides and inulooligosaccharides: Recent advances and current perspectives. J. Ind. Microbiol. Biotechnol. 2014, 41, 893–906. [Google Scholar] [CrossRef]
- Sabater-Molina, M.; Larque, E.; Torrella, F.; Zamora, S. Dietary fructooligosaccharides and potential benefits on health. J. Physiol. Biochem. 2009, 65, 315–328. [Google Scholar] [CrossRef]
- Jovanovic-Malinovska, R.; Kuzmanova, S.; Winkelhausen, E. Oligosaccharide Profile in Fruits and Vegetables as Sources of Prebiotics and Functional Foods. Int. J. Food Prop. 2014, 17, 949–965. [Google Scholar] [CrossRef]
- Chow, J. Probiotics and prebiotics: A brief overview. J. Ren. Nutr. 2002, 12, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in healthy young population. Sci. Rep. 2017, 7, 11789. [Google Scholar] [CrossRef] [PubMed]
- Cherbut, C.; Michel, C.; Lecannu, G. The prebiotic characteristics of fructooligosaccharides are necessary for reduction of TNBS-induced colitis in rats. J. Nur. 2003, 133, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-W.; Park, Y.-S.; Jung, J.-S.; Shin, W.-S. Chitosan oligosaccharides, dp 2–8, have prebiotic effect on the Bifidobacterium bifidium and Lactobacillus sp. Anaerobe 2002, 8, 319–324. [Google Scholar] [CrossRef]
- Ding, S.; Wang, Y.; Yan, W.; Li, A.; Jiang, H.; Fang, J. Effects of Lactobacillus plantarum 15-1 and fructooligosaccharides on the response of broilers to pathogenic Escherichia coli O78 challenge. PLoS ONE 2019, 14, e0212079. [Google Scholar] [CrossRef]
- Rubio-Gomez, J.M.; Santiago, C.M.; Udaondo, Z.; Garitaonaindia, M.T.; Krell, T.; Ramos, J.L.; Daddaoua, A. Full Transcriptomic Response of Pseudomonas aeruginosa to an Inulin-Derived Fructooligosaccharide. Front. Microbiol. 2020, 11, 202. [Google Scholar] [CrossRef]
- De Figueiredo, F.C.; de Barros Ranke, F.F.; de Oliva-Neto, P. Evaluation of xylooligosaccharides and fructooligosaccharides on digestive enzymes hydrolysis and as a nutrient for different probiotics and Salmonella typhimurium. LWT-Food Sci. Technol. 2020, 118, 108761. [Google Scholar] [CrossRef]
- Piotrowski, M.; Wultanska, D.; Obuch-Woszczatynski, P.; Pituch, H. Fructooligosaccharides and mannose affect Clostridium difficile adhesion and biofilm formation in a concentration-dependent manner. Eur. J. Clin. Microbiol. Infect Dis. 2019, 38, 1975–1984. [Google Scholar] [CrossRef]
- Sharma, S.; Kanwar, S.S. Effect of prebiotics on growth behavior of Lactobacillus plantarum and their impact on adherence of strict anaerobic pathogens to intestinal cell lines. J. Food Saf. 2017, 38, e12384. [Google Scholar] [CrossRef]
- Anand, S.; Mandal, S.; Singh, K.S.; Patil, P.; Tomar, S.K. Synbiotic combination of Lactobacillus rhamnosus NCDC 298 and short chain fructooligosaccharides prevents enterotoxigenic Escherichia coli infection. LWT-Food Sci. Technol. 2018, 98, 329–334. [Google Scholar] [CrossRef]
- Kariyawasam, K.; Yang, S.J.; Lee, N.K.; Paik, H.D. Probiotic Properties of Lactobacillus brevis KU200019 and Synergistic Activity with Fructooligosaccharides in Antagonistic Activity against Foodborne Pathogens. Food Sci. Anim. Resour. 2020, 40, 297–310. [Google Scholar] [CrossRef]
- Radoshevich, L.; Cossart, P. Listeria monocytogenes: Towards a complete picture of its physiology and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 32–46. [Google Scholar] [CrossRef]
- Pereira, M.G.; de Almeida, O.G.G.; da Silva, H.R.A.; Ishizawa, M.H.; De Martinis, E.C.P. Studies on host-foodborne bacteria in intestinal three-dimensional cell culture model indicate possible mechanisms of interaction. World J. Microbiol. Biotechnol. 2021, 37, 31. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Hu, X.; Chen, Y.; Xie, J.; Ying, M.; Wang, Y.; Yu, Q. Differentiated Caco-2 cell models in food-intestine interaction study: Current applications and future trends. Trends Food Sci. Technol. 2021, 107, 455–465. [Google Scholar] [CrossRef]
- Gagnon, M.; Zihler Berner, A.; Chervet, N.; Chassard, C.; Lacroix, C. Comparison of the Caco-2, HT-29 and the mucus-secreting HT29-MTX intestinal cell models to investigate Salmonella adhesion and invasion. J. Microbiol. Methods 2013, 94, 274–279. [Google Scholar] [CrossRef]
- Li, Z.; Kurosawa, O.; Iwata, H. A comparative study of key physiological stem cell parameters between three human trophoblast cell lines. Biochem. Biophys. Res. Commun. 2020, 525, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kurosawa, O.; Iwata, H. Establishment of human trophoblast stem cells from human induced pluripotent stem cell-derived cystic cells under micromesh culture. Stem. Cell Res. Ther. 2019, 10, 245. [Google Scholar] [CrossRef] [PubMed]
- Ye, K.; Zhang, X.; Huang, Y.; Liu, J.; Liu, M.; Zhou, G. Bacteriocinogenic Enterococcus faecium inhibits the virulence property of Listeria monocytogenes. LWT-Food Sci. Technol. 2018, 89, 87–92. [Google Scholar] [CrossRef]
- Mathipa, M.G.; Bhunia, A.K.; Thantsha, M.S. Internalin AB-expressing recombinant Lactobacillus casei protects Caco-2 cells from Listeria monocytogenes-induced damages under simulated intestinal conditions. PLoS ONE 2019, 14, e0220321. [Google Scholar] [CrossRef]
- Lecuit, M.; Nelson, D.M.; Smith, S.D.; Khun, H.; Huerre, M.; Vacher-Lavenu, M.C.; Gordon, J.I.; Cossart, P. Targeting and crossing of the human maternofetal barrier by Listeria monocytogenes: Role of internalin interaction with trophoblast E-cadherin. Proc. Natl. Acad. Sci. USA 2004, 101, 6152–6157. [Google Scholar] [CrossRef]
- Phelps, C.C.; Vadia, S.; Arnett, E.; Tan, Y.; Zhang, X.; Pathak-Sharma, S.; Gavrilin, M.A.; Seveau, S. Relative Roles of Listeriolysin O, InlA, and InlB in Listeria monocytogenes Uptake by Host Cells. Infect. Immun. 2018, 86, e00555-18. [Google Scholar] [CrossRef]
- Bakardjiev, A.I.; Stacy, B.A.; Fisher, S.J.; Portnoy, D.A. Listeriosis in the pregnant guinea pig: A model of vertical transmission. Infect. Immun. 2004, 72, 489–497. [Google Scholar] [CrossRef]
- Tsai, Y.H.; Disson, O.; Bierne, H.; Lecuit, M. Murinization of internalin extends its receptor repertoire, altering Listeria monocytogenes cell tropism and host responses. PLoS Pathog 2013, 9, e1003381. [Google Scholar] [CrossRef]
- Wollert, T.; Pasche, B.; Rochon, M.; Deppenmeier, S.; van den Heuvel, J.; Gruber, A.D.; Heinz, D.W.; Lengeling, A.; Schubert, W.D. Extending the host range of Listeria monocytogenes by rational protein design. Cell 2007, 129, 891–902. [Google Scholar] [CrossRef]
- Saá Ibusquiza, P.; Nierop Groot, M.; Debán-Valles, A.; Abee, T.; den Besten, H.M.W. Impact of growth conditions and role of sigB on Listeria monocytogenes fitness in single and mixed biofilms cultured with Lactobacillus plantarum. Food Res. Int. 2015, 71, 140–145. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Nogacka, A.; Salazar, N.; Hernandez-Barranco, A.M.; Cuesta, I.; Gueimonde, M.; de Los Reyes Gavilan, C.G. Bifidobacterium breve IPLA20005 affects in vitro the expression of hly and luxS genes, related to the virulence of Listeria monocytogenes Lm23. Can. J. Microbiol. 2018, 64, 215–221. [Google Scholar] [CrossRef]
- Olesen, I.; Vogensen, F.K.; Jespersen, L. Gene Transcription and Virulence Potential of Listeria monocytogenes Strains after Exposure to Acidic and NaCl Stress. Foodborne Pathog. Dis. 2009, 6, 669–680. [Google Scholar] [CrossRef]
- Kariyawasam, K.M.G.M.M.; Lee, N.-K.; Paik, H.-D. Synbiotic yoghurt supplemented with novel probiotic Lactobacillus brevis KU200019 and fructooligosaccharides. Food Biosci. 2021, 39, 100835. [Google Scholar] [CrossRef]
- Zilelidou, E.A.; Rychli, K.; Manthou, E.; Ciolacu, L.; Wagner, M.; Skandamis, P.N. Highly Invasive Listeria monocytogenes Strains Have Growth and Invasion Advantages in Strain Competition. PLoS ONE 2015, 10, e0141617. [Google Scholar] [CrossRef] [PubMed]
- Zilelidou, E.A.; Milina, V.; Paramithiotis, S.; Zoumpopoulou, G.; Poimenidou, S.V.; Mavrogonatou, E.; Kletsas, D.; Papadimitriou, K.; Tsakalidou, E.; Skandamis, P.N. Differential Modulation of Listeria monocytogenes Fitness, In Vitro Virulence, and Transcription of Virulence-Associated Genes in Response to the Presence of Different Microorganisms. Food Microbiol. 2020, 86, e01165-20. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Miranda, R.O.; Campos-Galvao, M.E.M.; Nero, L.A. Expression of genes associated with stress conditions by Listeria monocytogenes in interaction with nisin producer Lactococcus lactis. Food Res. Int. 2018, 105, 897–904. [Google Scholar] [CrossRef]
- Saraoui, T.; Fall, P.A.; Leroi, F.; Antignac, J.P.; Chereau, S.; Pilet, M.F. Inhibition mechanism of Listeria monocytogenes by a bioprotective bacteria Lactococcus piscium CNCM I-4031. Food Microbiol. 2016, 53, 70–78. [Google Scholar] [CrossRef]
- Giello, M.; La Storia, A.; De Filippis, F.; Ercolini, D.; Villani, F. Impact of Lactobacillus curvatus 54M16 on microbiota composition and growth of Listeria monocytogenes in fermented sausages. Food Microbiol. 2018, 72, 1–15. [Google Scholar] [CrossRef]
- Cornu, M.; Billoir, E.; Bergis, H.; Beaufort, A.; Zuliani, V. Modeling microbial competition in food: Application to the behavior of Listeria monocytogenes and lactic acid flora in pork meat products. Food Microbiol. 2011, 28, 639–647. [Google Scholar] [CrossRef]
- Han, E.J.; Lee, N.K.; Choi, S.Y.; Paik, H.D. Short communication: Bacteriocin KC24 produced by Lactococcus lactis KC24 from kimchi and its antilisterial effect in UHT milk. J. Dairy Sci. 2013, 96, 101–104. [Google Scholar] [CrossRef] [PubMed]
- İncili, G.K.; Karatepe, P.; İlhak, O.İ. Effect of chitosan and Pediococcus acidilactici on E. coli O157:H7, Salmonella Typhimurium and Listeria monocytogenes in meatballs. LWT-Food Sci. Technol. 2020, 117, 108706. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M.; Speranza, B.; Campaniello, D.; Altieri, C. Effects of inulin, fructooligosaccharides/glucose and pH on the shape of the death kinetic of Lactobacillus reuteri DSM 20016. Int. J. Food Sci. Technol. 2016, 51, 2251–2259. [Google Scholar] [CrossRef]
- Lu, E.; Yeung, M.; Yeung, C.K. Comparative Analysis of Lactulose and Fructooligosaccharide on Growth Kinetics, Fermentation, and Antioxidant Activity of Common Probiotics. Food Nutr. Sci. 2018, 09, 161–178. [Google Scholar] [CrossRef]
- Ibarra-Sanchez, L.A.; Kong, W.; Lu, T.; Miller, M.J. Efficacy of nisin derivatives with improved biochemical characteristics, alone and in combination with endolysin PlyP100 to control Listeria monocytogenes in laboratory-scale Queso Fresco. Food Microbiol. 2021, 94, 103668. [Google Scholar] [CrossRef]
- Chang, S.S.; Lu, W.Y.; Park, S.H.; Kang, D.H. Control of foodborne pathogens on ready-to-eat roast beef slurry by epsilon-polylysine. Int. J. Food Microbiol. 2010, 141, 236–241. [Google Scholar] [CrossRef]
- Heir, E.; Solberg, L.E.; Carlehog, M.; Moen, B.; Jensen, M.R.; Holck, A.L. Improved control of Listeria monocytogenes during storage of raw salmon by treatment with the fermentate Verdad N6 and nisin. Int. J. Food Microbiol. 2021, 336, 108895. [Google Scholar] [CrossRef]
- Castellano, P.; Pérez Ibarreche, M.; Longo Borges, L.; Niño Arias, F.C.; Ross, G.R.; De Martinis, E.C.P. Lactobacillus spp. impair the ability of Listeria monocytogenes FBUNT to adhere to and invade Caco-2 cells. Biotechnol. Lett. 2018, 40, 1237–1244. [Google Scholar] [CrossRef]
- Iglesias, M.B.; Viñas, I.; Colás-Medà, P.; Collazo, C.; Serrano, J.C.E.; Abadias, M. Adhesion and invasion of Listeria monocytogenes and interaction with Lactobacillus rhamnosus GG after habituation on fresh-cut pear. J. Funct. Foods 2017, 34, 453–460. [Google Scholar] [CrossRef]
- Moroni, O.; Kheadr, E.; Boutin, Y.; Lacroix, C.; Fliss, I. Inactivation of adhesion and invasion of food-borne Listeria monocytogenes by bacteriocin-producing Bifidobacterium strains of human origin. Appl. Environ. Microbiol. 2006, 72, 6894–6901. [Google Scholar] [CrossRef]
- Chen, P.; Reiter, T.; Huang, B.; Kong, N.; Weimer, B.C. Prebiotic Oligosaccharides Potentiate Host Protective Responses against L. Monocytogenes Infection. Pathogens 2017, 6, 68. [Google Scholar] [CrossRef]
- Vazquez-Boland, J.A.; Krypotou, E.; Scortti, M. Listeria Placental Infection. MBio 2017, 8, e00949-17. [Google Scholar] [CrossRef]
- Rothbauer, M.; Patel, N.; Gondola, H.; Siwetz, M.; Huppertz, B.; Ertl, P. A comparative study of five physiological key parameters between four different human trophoblast-derived cell lines. Sci. Rep. 2017, 7, 5892. [Google Scholar] [CrossRef]
- Faralla, C.; Rizzuto, G.A.; Lowe, D.E.; Kim, B.; Cooke, C.; Shiow, L.R.; Bakardjiev, A.I. InlP, a New Virulence Factor with Strong Placental Tropism. Infect. Immun. 2016, 84, 3584–3596. [Google Scholar] [CrossRef]
- Hadjilouka, A.; Molfeta, C.; Panagiotopoulou, O.; Paramithiotis, S.; Mataragas, M.; Drosinos, E.H. Expression of Listeria monocytogenes key virulence genes during growth in liquid medium, on rocket and melon at 4, 10 and 30 degrees C. Food Microbiol. 2016, 55, 7–15. [Google Scholar] [CrossRef]
- Tanner, S.A.; Chassard, C.; Rigozzi, E.; Lacroix, C.; Stevens, M.J. Bifidobacterium thermophilum RBL67 impacts on growth and virulence gene expression of Salmonella enterica subsp. enterica serovar Typhimurium. BMC Microbiol. 2016, 16, 46. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Cao, T.; Cheng, C.; Xia, Y.; Shan, Y.; Xin, Y.; Guo, N.; Li, X.; Song, H.; Fang, W. Activation of PrfA results in overexpression of virulence factors but does not rescue the pathogenicity of Listeria monocytogenes M7. J. Med. Microbiol. 2015, 64, 818–827. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gaballa, A.; Guariglia-Oropeza, V.; Wiedmann, M.; Boor, K.J. Cross Talk between SigB and PrfA in Listeria monocytogenes Facilitates Transitions between Extra- and Intracellular Environments. Microbiol. Mol. Biol. Rev. 2019, 83, e00034-19. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Length (bp) |
|---|---|---|
| rpoB | F: TCGTCGTCTTCGTTCTGTTGR: GTTCGCCAAGTGGATTTGTT | 221 |
| inlA | F: ATAGGCACATTGGCGAGTTT | 160 |
| R: GTGCGGTTAAACCTGCTAGG | ||
| inlB | F: AAGCAMGATTTCATGGGAGAGT | 78 |
| R: TTACCGTTCCATCAACATCATAACTT | ||
| hly | F: CTTTTAACCGGGAAACACCA | 302 |
| R: TCTTGCGTTACCTGGCAAA | ||
| actA | F: CGGGTAAATGGGTACGTGAT | 85 |
| R: TGGTCAATTAACCCTGCACTT | ||
| prfA | F: CGGGAAGCTTGGCTCTATTTG | 150 |
| R: GCTAACAGCTGAGCTATGTGC | ||
| sigB | F: TCATCGGTGTCACGGAAGAA | 310 |
| R: TGACGTTGGATTCTAGACAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Q.; Lu, X.; Gao, B.; Liu, Y.; Aslam, M.Z.; Wang, X.; Li, Z. Lactiplantibacillus plantarum subsp. plantarum and Fructooligosaccharides Combination Inhibits the Growth, Adhesion, Invasion, and Virulence of Listeria monocytogenes. Foods 2022, 11, 170. https://doi.org/10.3390/foods11020170
Dong Q, Lu X, Gao B, Liu Y, Aslam MZ, Wang X, Li Z. Lactiplantibacillus plantarum subsp. plantarum and Fructooligosaccharides Combination Inhibits the Growth, Adhesion, Invasion, and Virulence of Listeria monocytogenes. Foods. 2022; 11(2):170. https://doi.org/10.3390/foods11020170
Chicago/Turabian StyleDong, Qingli, Xinxin Lu, Binru Gao, Yangtai Liu, Muhammad Zohaib Aslam, Xiang Wang, and Zhuosi Li. 2022. "Lactiplantibacillus plantarum subsp. plantarum and Fructooligosaccharides Combination Inhibits the Growth, Adhesion, Invasion, and Virulence of Listeria monocytogenes" Foods 11, no. 2: 170. https://doi.org/10.3390/foods11020170
APA StyleDong, Q., Lu, X., Gao, B., Liu, Y., Aslam, M. Z., Wang, X., & Li, Z. (2022). Lactiplantibacillus plantarum subsp. plantarum and Fructooligosaccharides Combination Inhibits the Growth, Adhesion, Invasion, and Virulence of Listeria monocytogenes. Foods, 11(2), 170. https://doi.org/10.3390/foods11020170