

Structural Characterization and Evaluation of Interfacial Properties of Pea Protein Isolate–EGCG Molecular Complexes

 ,
, 
Abstract
:Highlights
- Pea protein isolate (PPI) and EGCG spontaneously formed complexes.
- Protein–polyphenol complexation was mainly driven by hydrogen bonding.
- The binding of EGCG influenced the structure and functionality of PPI.
- PPI-EGCG complexes had better emulsifier properties than PPI.
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of PPI Dispersions and PPI-EGCG Complexes
2.3. Spectroscopic Characterization of PPI-EGCG Complexes
2.3.1. Fluorescence Spectroscopy
2.3.2. UV Spectroscopy
2.3.3. Circular Dichroism (CD)
2.3.4. Fourier Transform Infrared Spectroscopy (FTIR)
2.4. Thermodynamic Characterization of PPI-EGCG Complexes
2.4.1. Isothermal Titration Calorimetry (ITC)
2.4.2. Differential Scanning Calorimetry (DSC)
2.5. Molecular Docking of PPI and EGCG
2.6. Physicochemical Properties of PPI-EGCG Complexes
2.6.1. Surface Hydrophobicity (H0)
2.6.2. Turbidity
2.6.3. Particle Properties
2.7. Interfacial Properties of PPI-EGCG Complexes
2.7.1. Foaming Properties
2.7.2. Emulsifying Properties
2.8. Statistical Analysis
3. Results and Discussion
3.1. Spectroscopic Analysis of PPI-EGCG Complexes
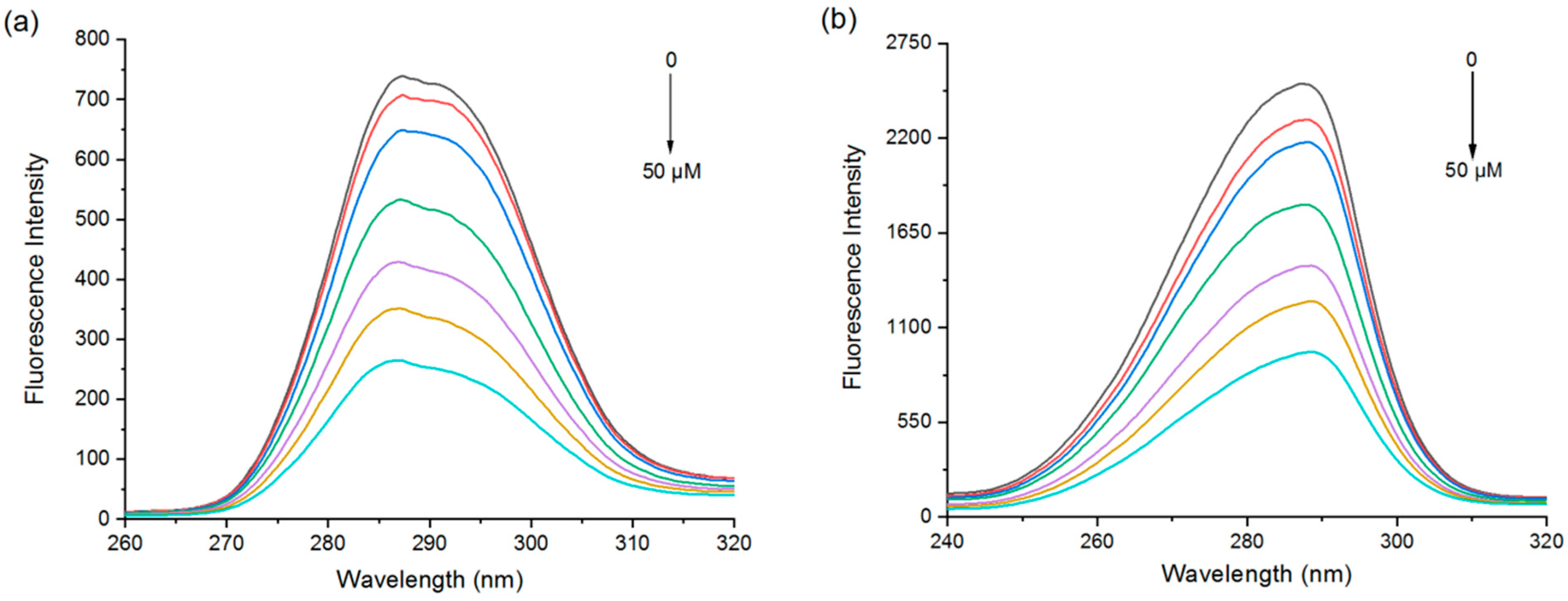
3.1.1. Variable-Temperature Fluorescence Spectroscopy
3.1.2. Synchronous Fluorescence Spectroscopy
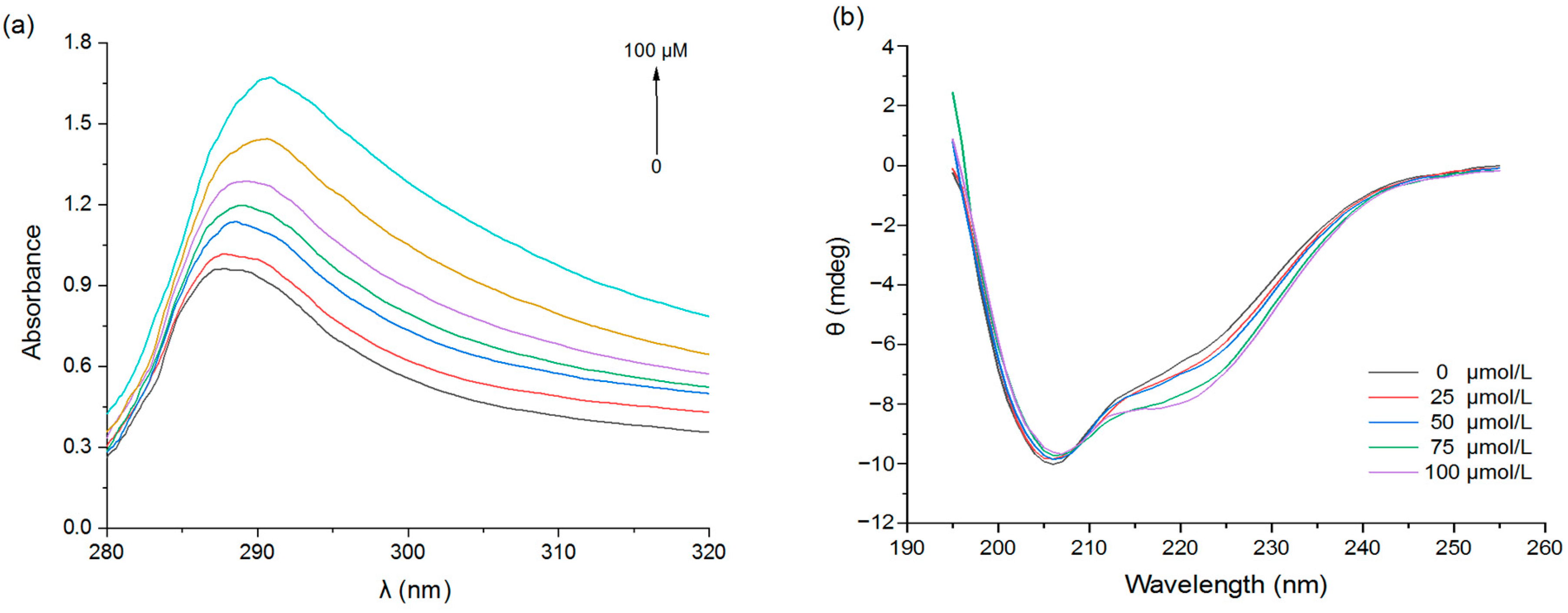
3.1.3. UV Spectroscopy
3.1.4. Circular Dichroism (CD)
3.1.5. Fourier Transform Infrared Spectroscopy (FTIR)
3.2. Thermodynamics Analysis of PPI-EGCG Complexes
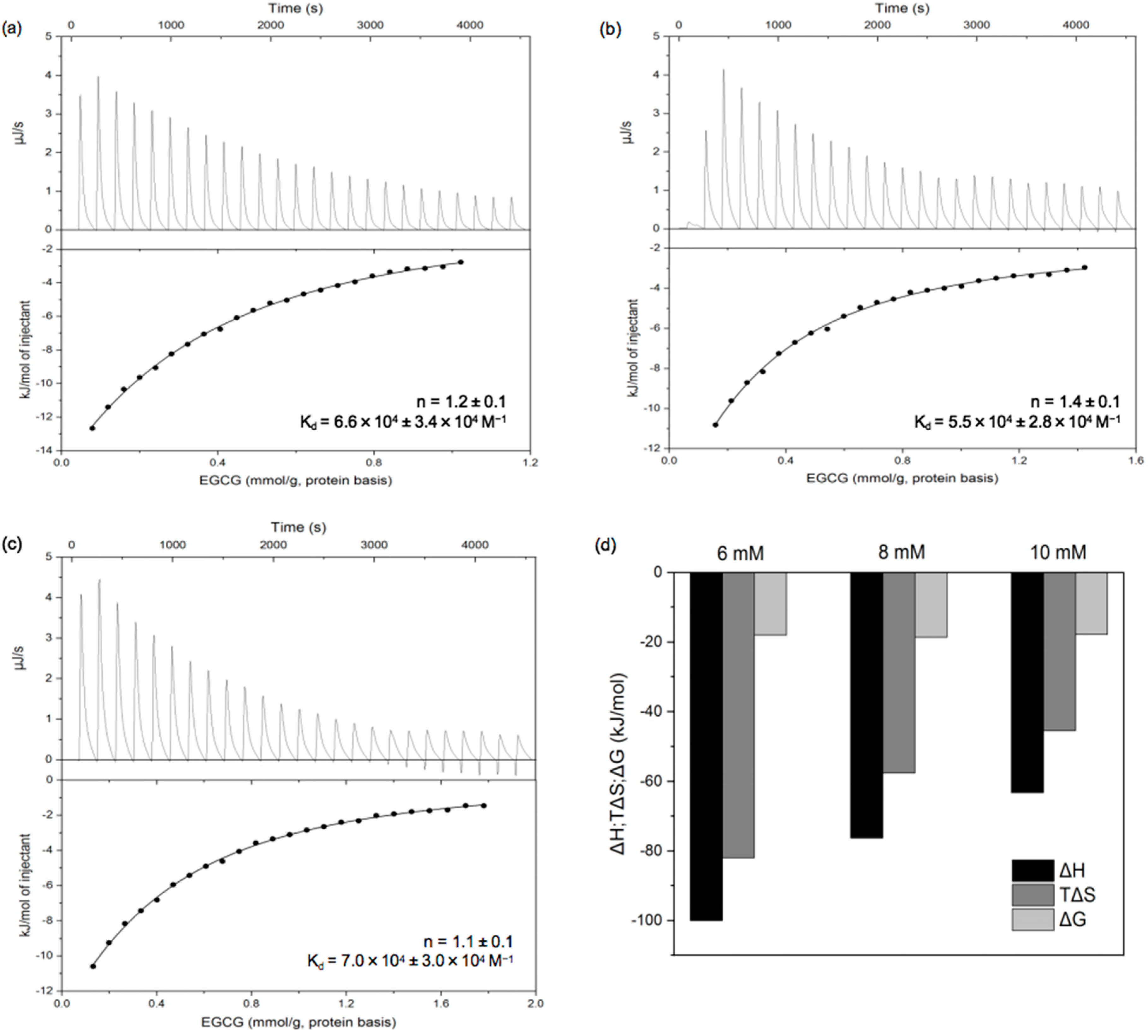
3.2.1. Isothermal Titration Calorimetry (ITC)
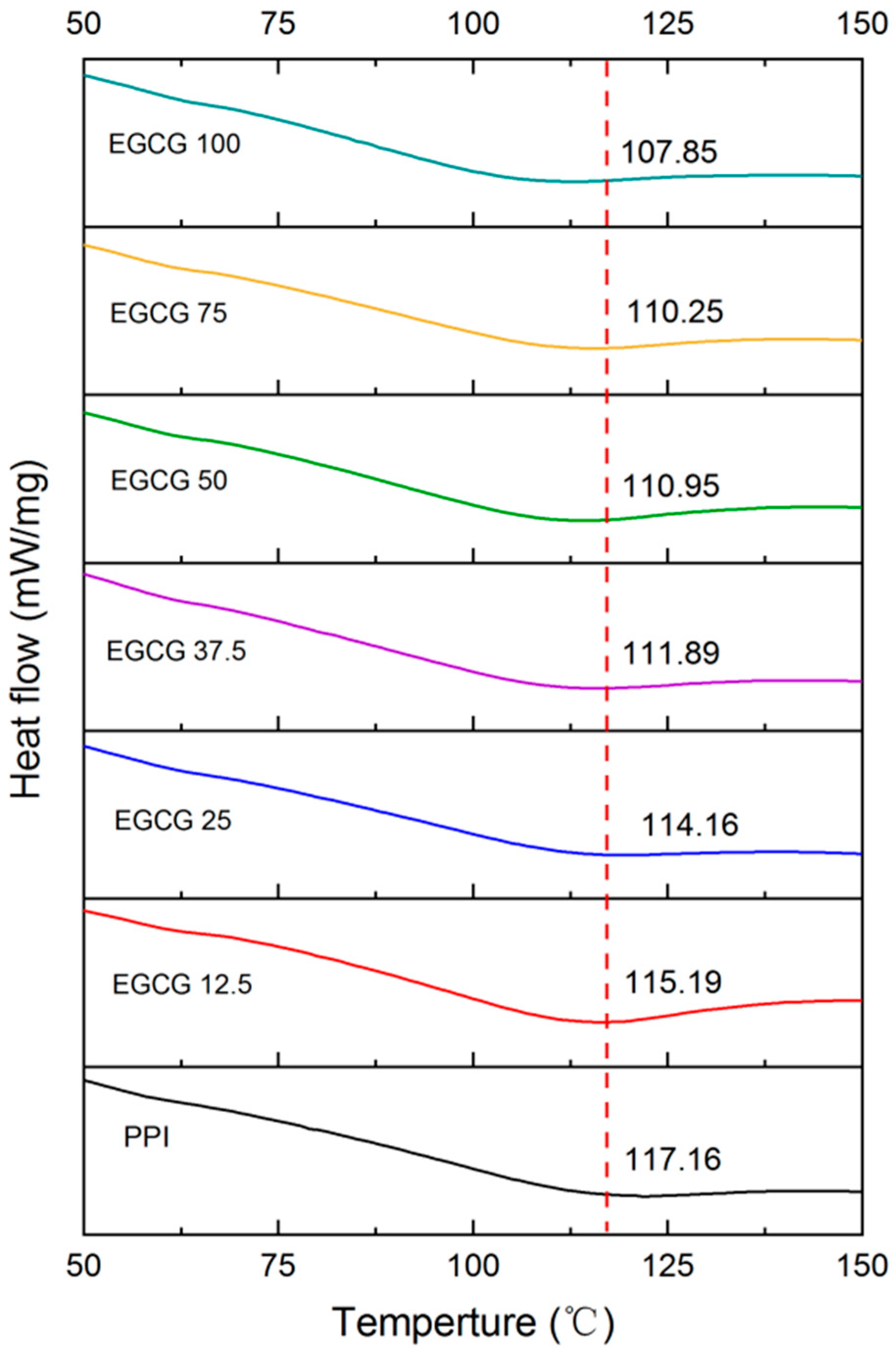
3.2.2. Differential Scanning Calorimetry (DSC)
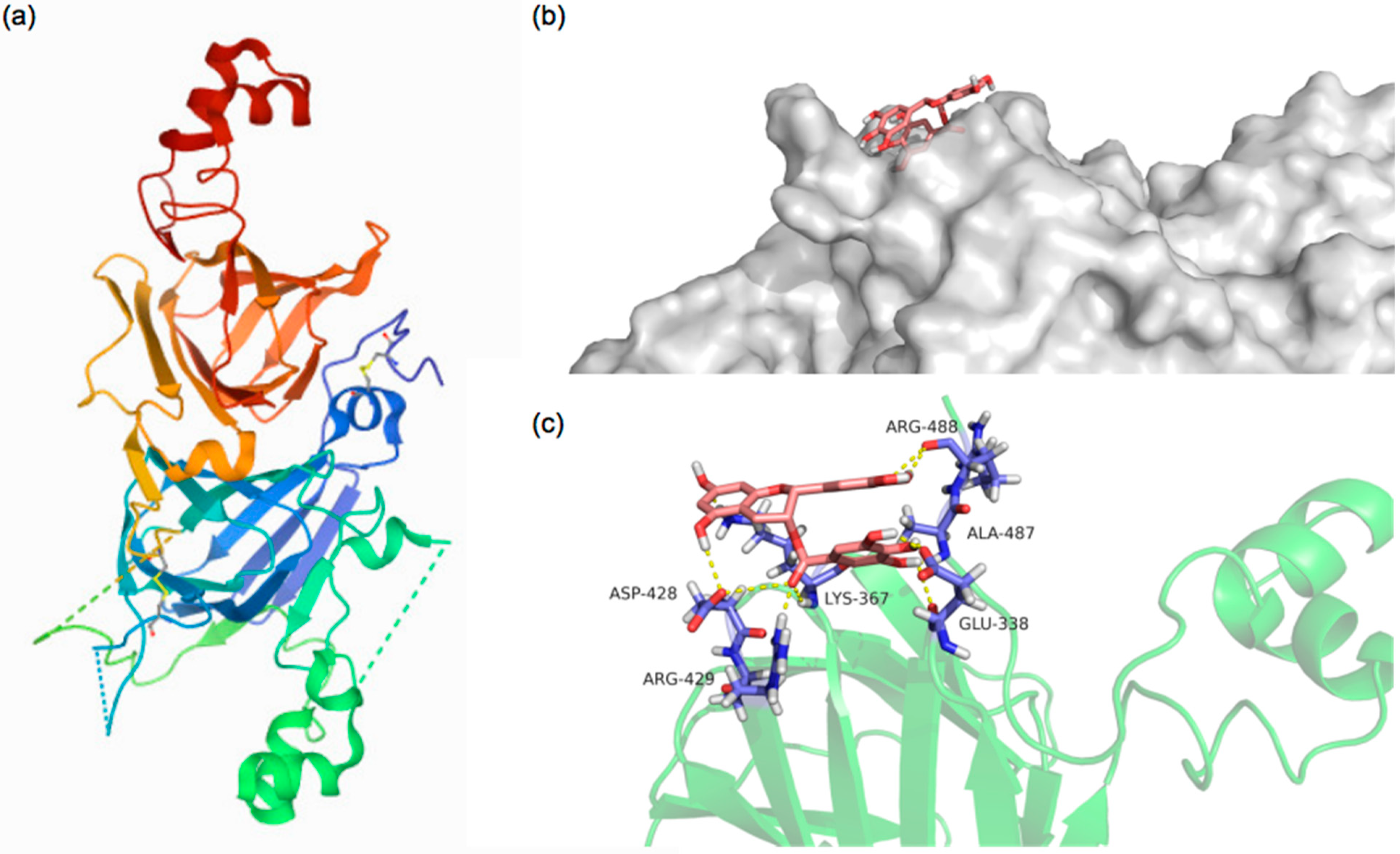
3.3. Molecular Docking Analysis
3.4. Physicochemical Property Analysis of PPI-EGCG Complexes
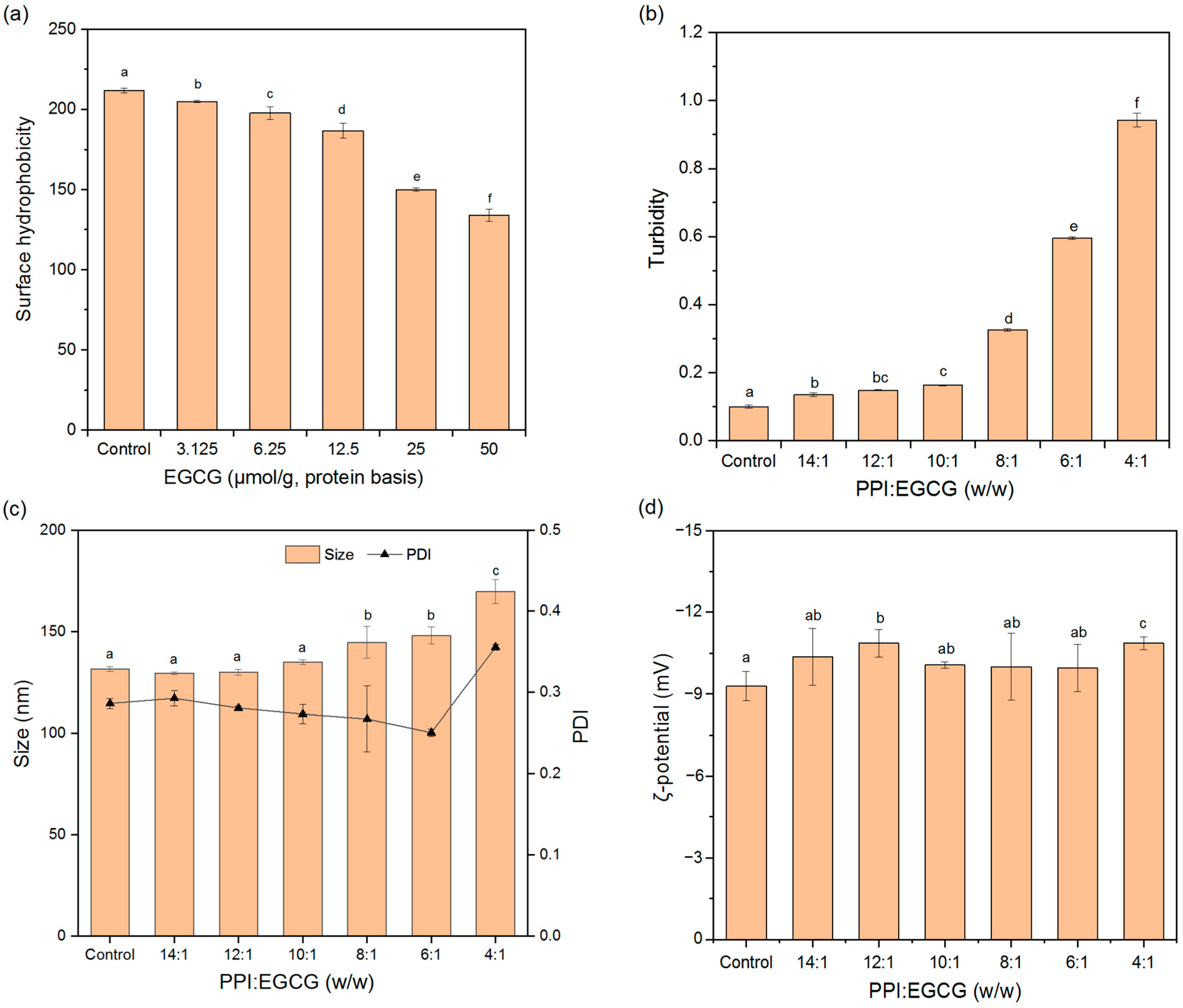
3.4.1. Surface Hydrophobicity (H0)
3.4.2. Turbidity
3.4.3. Particle Properties
3.5. Interfacial Property Analysis of PPI-EGCG Complexes
3.5.1. Foaming Properties
3.5.2. Emulsifying Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef]
- Willett, W. Healthy Diets From Sustainable Food Systems: Summary Report of the EAT-Lancet Commission. EAT-Lancet Comm. 2019, 393, 447–492. [Google Scholar] [CrossRef]
- McClements, D.J.; Grossmann, L. The science of plant-based foods: Constructing next-generation meat, fish, milk, and egg analogs. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4049–4100. [Google Scholar] [CrossRef]
- McClements, D.J.; Grossmann, L. A brief review of the science behind the design of healthy and sustainable plant-based foods. NPJ Sci. Food 2021, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hua, Y.; Li, X.; Kong, X.; Chen, Y. Key volatile off-flavor compounds in peas (Pisum sativum L.) and their relations with the endogenous precursors and enzymes using soybean (Glycine max) as a reference. Food Chem. 2020, 333, 127469. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, N.A.; Kennedy, D.; Hogan, S.A.; Kelly, P.M.; Thapa, K.; Murphy, K.M.; Fenelon, M.A. Emulsification properties of pea protein isolate using homogenization, microfluidization and ultrasonication. Food Res. Int. 2016, 89, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, S.; Chen, Q.; Liu, Q.; Kong, B. Antioxidant activities and emulsifying properties of porcine plasma protein hydrolysates modified by oxidized tannic acid and oxidized chlorogenic acid. Process Biochem. 2019, 79, 105–113. [Google Scholar] [CrossRef]
- Ge, J.; Sun, C.X.; Corke, H.; Gul, K.; Gan, R.Y.; Fang, Y. The health benefits, functional properties, modifications, and applications of pea (Pisum sativum L.) protein: Current status, challenges, and perspectives. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1835–1876. [Google Scholar] [CrossRef] [PubMed]
- Pirestani, S.; Nasirpour, A.; Keramat, J.; Desobry, S.; Jasniewski, J. Effect of glycosylation with gum Arabic by Maillard reaction in a liquid system on the emulsifying properties of canola protein isolate. Carbohydr. Polym. 2017, 157, 1620–1627. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, Y.; Ma, C.; McClements, D.J.; Liu, F.; Liu, X. Pea protein isolate-inulin conjugates prepared by pH-shift treatment and ultrasonic-enhanced glycosylation: Structural and functional properties. Food Chem. 2022, 384, 132511. [Google Scholar] [CrossRef]
- Chao, D.; Aluko, R.E. Modification of the structural, emulsifying, and foaming properties of an isolated pea protein by thermal pretreatment. CyTA-J. Food 2018, 16, 357–366. [Google Scholar] [CrossRef]
- Wei, Y.; Cai, Z.; Wu, M.; Guo, Y.; Tao, R.; Li, R.; Wang, P.; Ma, A.; Zhang, H. Comparative studies on the stabilization of pea protein dispersions by using various polysaccharides. Food Hydrocoll. 2020, 98, 105233. [Google Scholar] [CrossRef]
- Quan, T.H.; Benjakul, S.; Sae-leaw, T.; Balange, A.K.; Maqsood, S. Protein-polyphenol conjugates: Antioxidant property, functionalities and their applications. Trends Food Sci. Technol. 2019, 91, 507–517. [Google Scholar] [CrossRef]
- Park, R.; Jang, M.; Park, Y.I.; Park, Y.; Jung, W.; Park, J.; Park, J. Epigallocatechin gallate (EGCG), a green tea polyphenol, reduces coronavirus replication in a mouse model. Viruses 2021, 13, 2533. [Google Scholar] [CrossRef]
- McClements, D.J. Designing biopolymer microgels to encapsulate, protect and deliver bioactive components: Physicochemical aspects. Adv. Colloid Interface Sci. 2017, 240, 31–59. [Google Scholar] [CrossRef]
- Cheng, J.; Zhu, M.; Liu, X. Insight into the conformational and functional properties of myofibrillar protein modified by mulberry polyphenols. Food Chem. 2020, 308, 125592. [Google Scholar] [CrossRef]
- Arimboor, R.; Arumughan, C. Sea buckthorn (hippophae rhamnoides) proanthocyanidins inhibit in vitro enzymatic hydrolysis of protein. J. Food Sci. 2011, 76, 130–137. [Google Scholar] [CrossRef]
- Jia, Z.; Zheng, M.; Tao, F.; Chen, W.; Huang, G.; Jiang, J. Effect of covalent modification by (−)-epigallocatechin-3-gallate on physicochemical and functional properties of whey protein isolate. LWT-Food Sci. Technol. 2016, 66, 305–310. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Y.; Ma, C.; McClements, D.J.; Liu, F.; Liu, X. The effect of non-covalent interaction of chlorogenic acid with whey protein and casein on physicochemical and radical-scavenging activity of in vitro protein digests. Food Chem. 2018, 268, 334–341. [Google Scholar] [CrossRef]
- Li, Q.; Shi, J.L.; Du, X.L.; McClements, D.J.; Chen, X.Q.; Duan, M.R.; Liu, L.; Li, J.; Shao, Y.D.; Cheng, Y. Polysaccharide conjugates from Chin brick tea (Camellia sinensis) improve the physicochemical stability and bioaccessibility of β-carotene in oil-in-water nanoemulsions. Food Chem. 2021, 357, 129714. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, Y.; Gao, L.; Zhang, Y.; Yi, J. Oxidative stability and in vitro digestion of menhaden oil emulsions with whey protein: Effects of EGCG conjugation and interfacial cross-linking. Food Chem. 2018, 265, 200. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.D.; Lin, Y.F.; Xu, X.; Meng, L.; Dong, M.S. Effect of non-covalent and covalent complexation of (−)-epigallocatechin gallate with soybean protein isolate on protein structure and in vitro digestion characteristics. Food Chem. 2020, 309, 125718. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Sun, J.; Pei, M.; Zhang, G.; Li, C.; Li, C.; Ma, X.; He, S.; Liu, L. Impact of non-covalent bound polyphenols on conformational, functional properties and in vitro digestibility of pea protein. Food Chem. 2022, 383, 132623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liang, L.; Tian, Z.; Chen, L.; Subirade, M. Preparation and in vitro evaluation of calcium-induced soy protein isolate nanoparticles and their formation mechanism study. Food Chem. 2012, 133, 390–399. [Google Scholar] [CrossRef]
- Warake, R.A.; Jarag, R.J.; Dhavale, R.P.; Jarag, R.R.; Lohar, N.S. Evaluation of in vitro antioxidant, anticancer activities and molecular docking studies of Capparis zeylanica Linn. leaves. Future J. Pharm. Sci. 2021, 7, 1–12. [Google Scholar] [CrossRef]
- Zhan, F.; Yang, J.; Li, J.; Wang, Y.; Li, B. Characteristics of the interaction mechanism between tannic acid and sodium caseinate using multispectroscopic and thermodynamics methods. Food Hydrocoll. 2018, 75, 81–87. [Google Scholar] [CrossRef]
- Cui, F.; McClements, D.J.; Liu, X.; Liu, F.; Ngai, T. Development of pH-responsive emulsions stabilized by whey protein fibrils. Food Hydrocoll. 2022, 122, 107067. [Google Scholar] [CrossRef]
- Sui, X.; Sun, H.; Qi, B.; Zhang, M.; Li, Y.; Jiang, L. Functional and conformational changes to soy proteins accompanying anthocyanins: Focus on covalent and non-covalent interactions. Food Chem. 2018, 245, 871–878. [Google Scholar] [CrossRef]
- Meng, Y.; Li, C. Conformational changes and functional properties of whey protein isolate-polyphenol complexes formed by non-covalent interaction. Food Chem. 2021, 364, 129622. [Google Scholar] [CrossRef]
- Al-Hanish, A.; Stanic-Vucinic, D.; Mihailovic, J.; Prodic, I.; Minic, S.; Stojadinovic, M.; Radibratovic, M.; Milcic, M.; Velickovic, T.C. Noncovalent interactions of bovine α-lactalbumin with green tea polyphenol, epigalocatechin-3-gallate. Food Hydrocoll. 2016, 61, 241–250. [Google Scholar] [CrossRef]
- Yue, Y.; Sun, Y.; Dong, Q.; Liu, R.; Yan, X.; Zhang, Y.; Liu, J. Interaction of human serum albumin with novel imidazole derivatives studied by spectroscopy and molecular docking. Luminescence 2016, 31, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Cai, X.; Luo, L.; Wang, J.; Zeng, L. Influence of tea polyphenol and bovine serum albumin on tea cream formation by multiple spectroscopy methods and molecular docking. Food Chem. 2020, 333, 127432. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, F.; Xu, C.; Yuan, F.; Gao, Y. Molecular interaction between (−)-epigallocatechin-3-gallate and bovine lactoferrin using multi-spectroscopic method and isothermal titration calorimetry. Food Res. Int. 2014, 64, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Biter, A.B.; Pollet, J.; Chen, W.H.; Strych, U.; Bottazzi, M.E. A method to probe protein structure from UV absorbance spectra. Anal. Biochem. 2019, 587, 113450. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Tian, L.; Shi, M.; Zhao, S.; Guo, S.; Luo, W.; Wang, C.; Tian, Z. Investigation of the interaction of a polyamine-modified flavonoid with bovine serum albumin (BSA) by spectroscopic methods and molecular simulation. J. Photochem. Photobiol. B Biol. 2020, 209, 111917. [Google Scholar] [CrossRef]
- Dai, T.; Chen, J.; McClements, D.J.; Li, T.; Liu, C. Investigation the interaction between procyanidin dimer and α-glucosidase: Spectroscopic analyses and molecular docking simulation. Int. J. Biol. Macromol. 2019, 130, 315–322. [Google Scholar] [CrossRef]
- Wu, X.; Lu, Y.; Xu, H.; Lin, D.; He, Z.; Wu, H.; Liu, L.; Wang, Z. Reducing the allergenic capacity of β-lactoglobulin by covalent conjugation with dietary polyphenols. Food Chem. 2018, 256, 427–434. [Google Scholar] [CrossRef]
- Kaspchak, E.; Mafra, L.I.; Mafra, M.R. Effect of heating and ionic strength on the interaction of bovine serum albumin and the antinutrients tannic and phytic acids, and its influence on in vitro protein digestibility. Food Chem. 2018, 252, 1–8. [Google Scholar] [CrossRef]
- Tayyab, S.; Izzudin, M.M.; Kabir, M.Z.; Feroz, S.R.; Tee, W.V.; Mohamad, S.B.; Alias, Z. Binding of an anticancer drug, axitinib to human serum albumin: Fluorescence quenching and molecular docking study. J. Photochem. Photobiol. B Biol. 2016, 162, 386–394. [Google Scholar] [CrossRef]
- Liu, F.; Wang, D.; Ma, C.; Gao, Y. Conjugation of polyphenols prevents lactoferrin from thermal aggregation at neutral pH. Food Hydrocoll. 2016, 58, 49–59. [Google Scholar] [CrossRef]
- Cao, Y.; Xiong, Y.L. Interaction of whey proteins with phenolic derivatives under neutral and acidic pH conditions. J. Food Sci. 2017, 82, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ma, C.; McClements, D.J.; Gao, Y. A comparative study of covalent and non-covalent interactions between zein and polyphenols in ethanol-water solution. Food Hydrocoll. 2017, 63, 625–634. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, C.; Qi, Z.; Zhao, R.; Wang, C.; Zhang, T. Pea protein based nanocarriers for lipophilic polyphenols: Spectroscopic analysis, characterization, chemical stability, antioxidant and molecular docking. Food Res. Int. 2022, 160, 111713. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Dong, Y.; Zhang, A.; Wang, X.; Zhao, X.H. Multiple spectra analysis and calculation of the interaction between Anthocyanins and whey protein isolate. Food Biosci. 2021, 44, 101353. [Google Scholar] [CrossRef]
- Cao, Y.; Xiong, Y.L.; Cao, Y.; True, A.D. Interfacial properties of whey protein foams as influenced by preheating and phenolic binding at neutral pH. Food Hydrocoll. 2018, 82, 379–387. [Google Scholar] [CrossRef]
- Ding, D. Effects of emulsifier on emulsification, physical and chemical properties of soybean protein. IOP Conf. Ser. Earth Environ. Sci. 2021, 792, 012017. [Google Scholar] [CrossRef]
- Afizah, M.N.; Rizvi, S.S. Functional properties of whey protein concentrate texturized at acidic pH: Effect of extrusion temperature. LWT-Food Sci. Technol. 2014, 57, 290–298. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T(K) | Ksv280nm(L·mol−1) | R2 | Kq280nm(L·mol−1s−1) | Ksv295nm(L·mol−1) | R2 | Kq295nm(L·mol−1s−1) |
|---|---|---|---|---|---|---|
| 298 | 2.796 × 104 | 0.991 | 2.796 × 1012 | 2.874 × 104 | 0.988 | 2.874 × 1012 |
| 304 | 3.312 × 104 | 0.997 | 3.312 × 1012 | 3.049 × 104 | 0.998 | 3.049 × 1012 |
| 310 | 3.314 × 104 | 0.995 | 3.314 × 1012 | 3.564 × 104 | 0.997 | 3.564 × 1012 |
| T(K) | Ka280nm(L·mol−1) | n280nm | R2 | Ka295nm(L·mol−1) | n290nm | R2 |
|---|---|---|---|---|---|---|
| 298 | 1.09 × 105 | 1.14 | 0.999 | 2.52 × 105 | 1.00 | 0.995 |
| 304 | 4.80 × 105 | 1.04 | 0.971 | 3.86 × 105 | 1.03 | 0.997 |
| 310 | 5.00 × 105 | 1.05 | 0.987 | 6.37 × 105 | 1.07 | 0.973 |
| EGCG (10−5 mol/L) | α-Helix (%) | β-Sheet (%) | β-Turn (%) | Random Coil (%) |
|---|---|---|---|---|
| 0.00 | 27.0 ± 0.5 a | 16.2 ± 0.4 a | 21.9 ± 0.1 a | 35.0 ± 0.1 a |
| 2.50 | 26.9 ± 0.7 a | 16.8 ± 0.4 ab | 21.7 ± 0.2 ab | 34.6 ± 0.2 b |
| 5.00 | 26.6 ± 0.6 a | 17.1 ± 0.3 b | 21.7 ± 0.2 ab | 34.6 ± 0.2 b |
| 7.50 | 25.1 ± 0.3 b | 19.3 ± 0.1 c | 21.5 ± 0.1 c | 34.1 ± 0.1 c |
| 10.0 | 24.7 ± 0.7 b | 19.7 ± 0.4 c | 21.6 ± 0.2 ab | 34.0 ± 0.1 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.; Cui, F.; McClements, D.J.; Xu, X.; Ma, C.; Wang, Y.; Liu, X.; Liu, F. Structural Characterization and Evaluation of Interfacial Properties of Pea Protein Isolate–EGCG Molecular Complexes. Foods 2022, 11, 2895. https://doi.org/10.3390/foods11182895
Han S, Cui F, McClements DJ, Xu X, Ma C, Wang Y, Liu X, Liu F. Structural Characterization and Evaluation of Interfacial Properties of Pea Protein Isolate–EGCG Molecular Complexes. Foods. 2022; 11(18):2895. https://doi.org/10.3390/foods11182895
Chicago/Turabian StyleHan, Shuang, Fengzhan Cui, David Julian McClements, Xingfeng Xu, Cuicui Ma, Yutang Wang, Xuebo Liu, and Fuguo Liu. 2022. "Structural Characterization and Evaluation of Interfacial Properties of Pea Protein Isolate–EGCG Molecular Complexes" Foods 11, no. 18: 2895. https://doi.org/10.3390/foods11182895
APA StyleHan, S., Cui, F., McClements, D. J., Xu, X., Ma, C., Wang, Y., Liu, X., & Liu, F. (2022). Structural Characterization and Evaluation of Interfacial Properties of Pea Protein Isolate–EGCG Molecular Complexes. Foods, 11(18), 2895. https://doi.org/10.3390/foods11182895