
The Starch Physicochemical Properties between Superior and Inferior Grains of Japonica Rice under Panicle Nitrogen Fertilizer Determine the Difference in Eating Quality

 ,
,  ,
,
Abstract
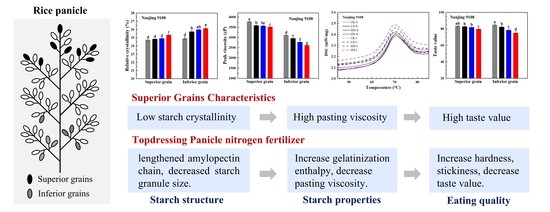
:
1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.2. Sampling
2.3. Measurements and Methods
2.3.1. Determination of Starch, Protein and Crude Fat Content
2.3.2. Starch Isolation
2.3.3. Analysis of Starch Granule Size Distribution
2.3.4. Analysis of Amylopectin Chain Length Distribution
2.3.5. Analysis of Starch X-ray Diffraction (XRD)
2.3.6. Analysis of Starch 13C CP/MAS NMR
2.3.7. Analysis of Starch Solubility and Swelling Power
2.3.8. Determination of Starch Thermal Properties
2.3.9. Determination of Starch Pasting Properties
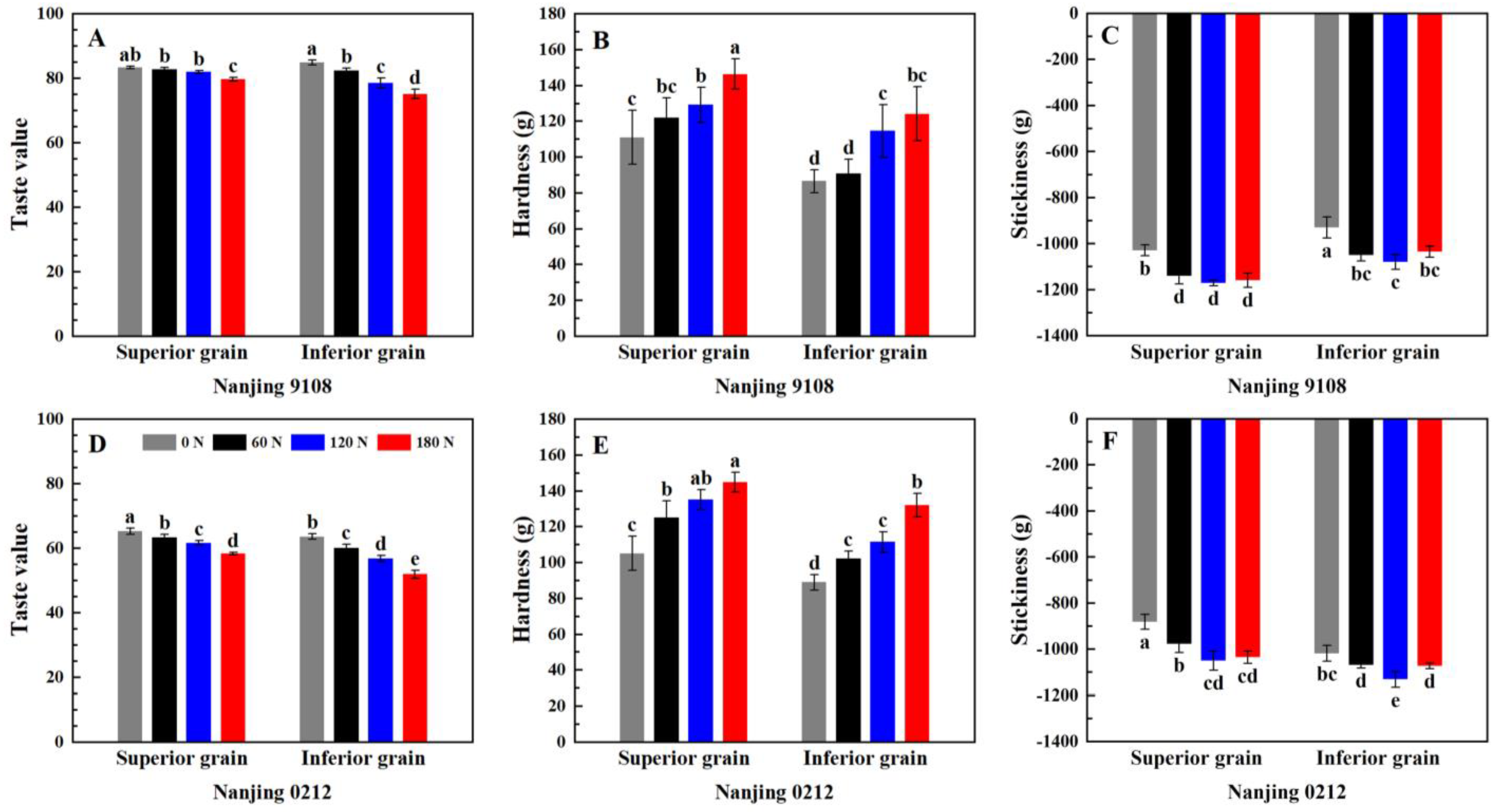
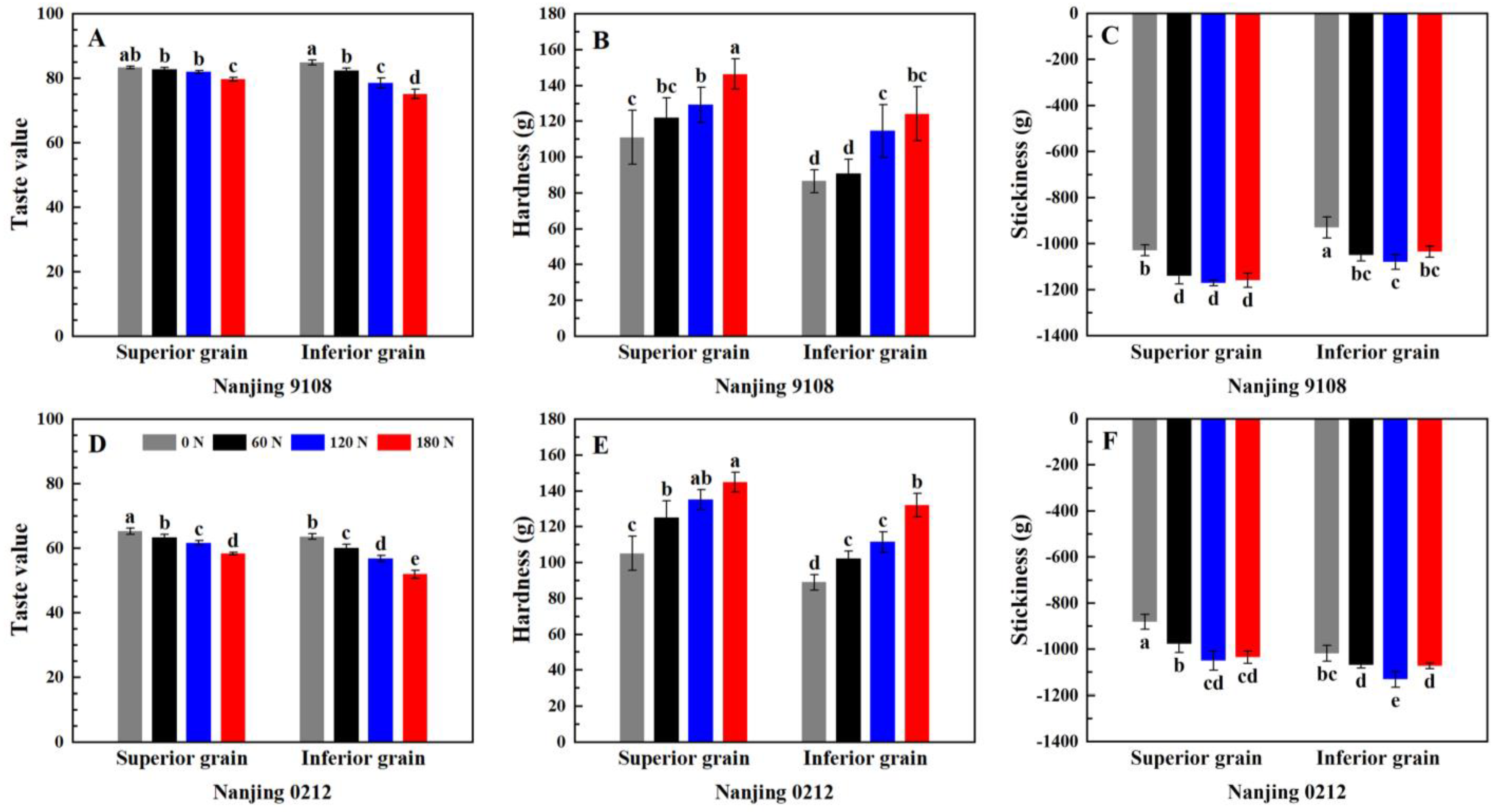
2.3.10. Analysis of Taste Value and Texture Properties
2.4. Statistical Processing and Analysis
3. Results and Discussion
3.1. Content of Starch, Protein and Crude Fat
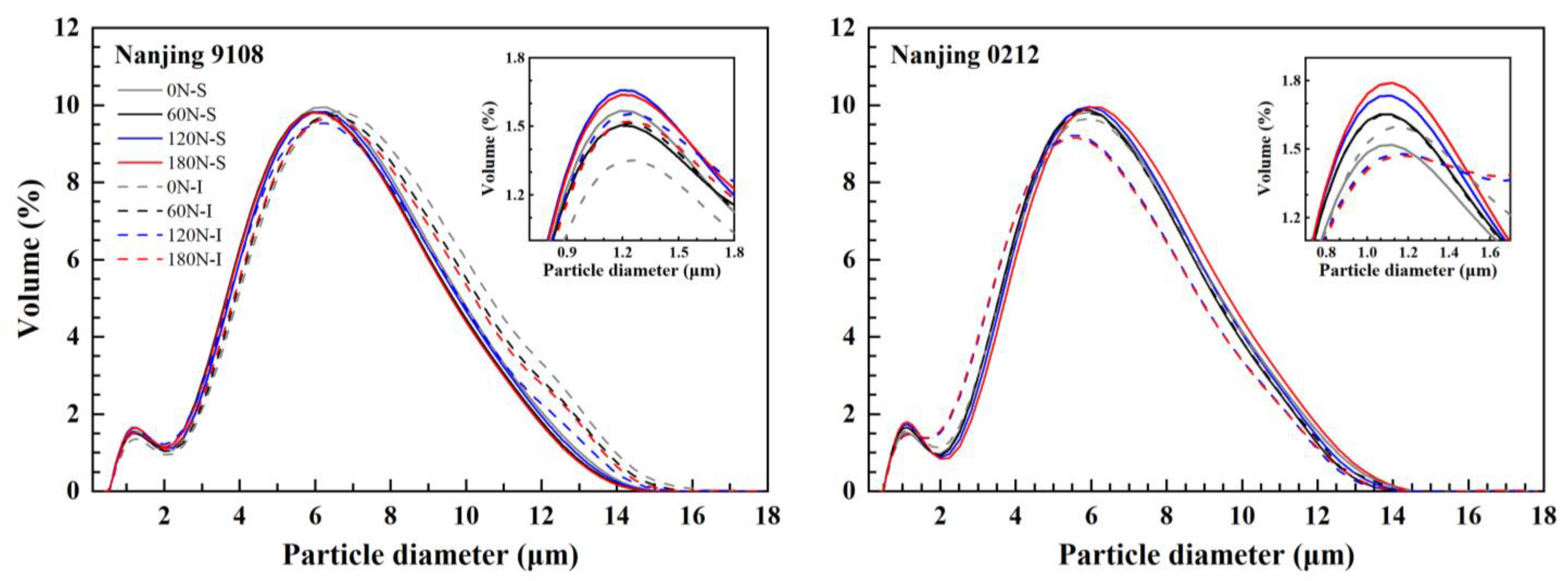
3.2. Starch Granule Size Distribution
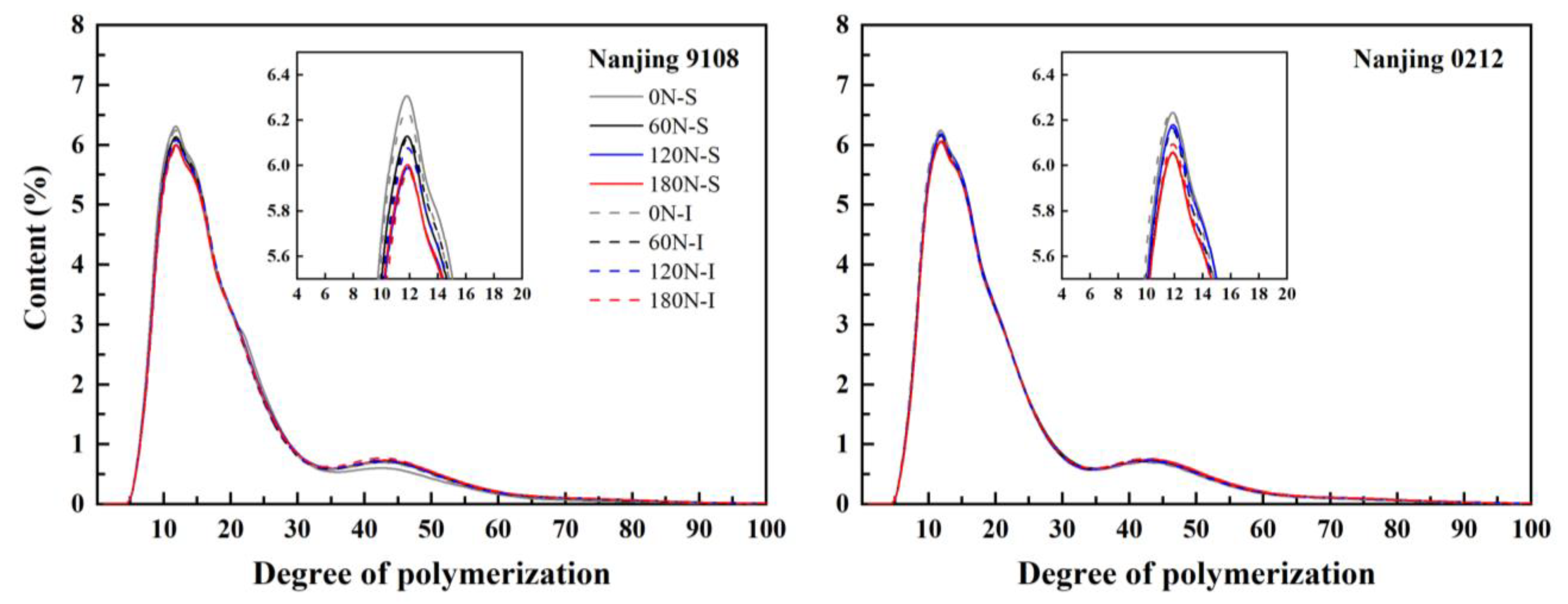
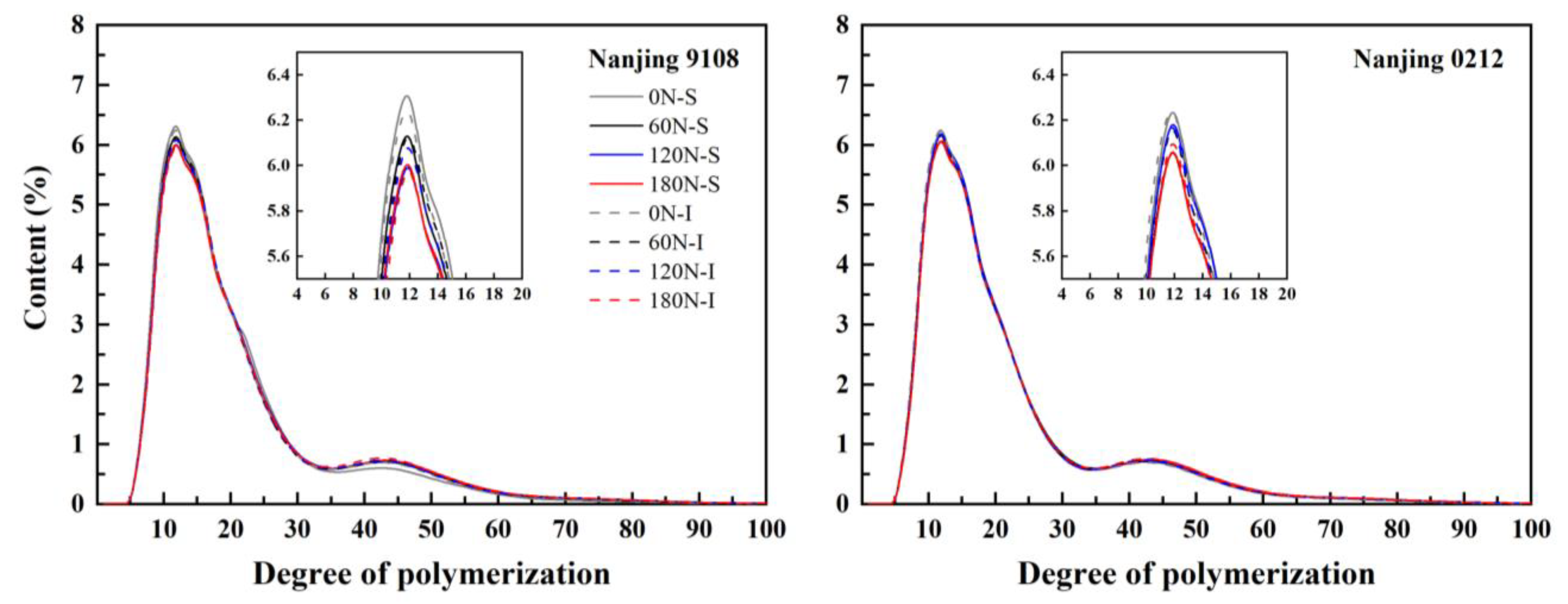
3.3. Amylopectin Chain Length Distribution
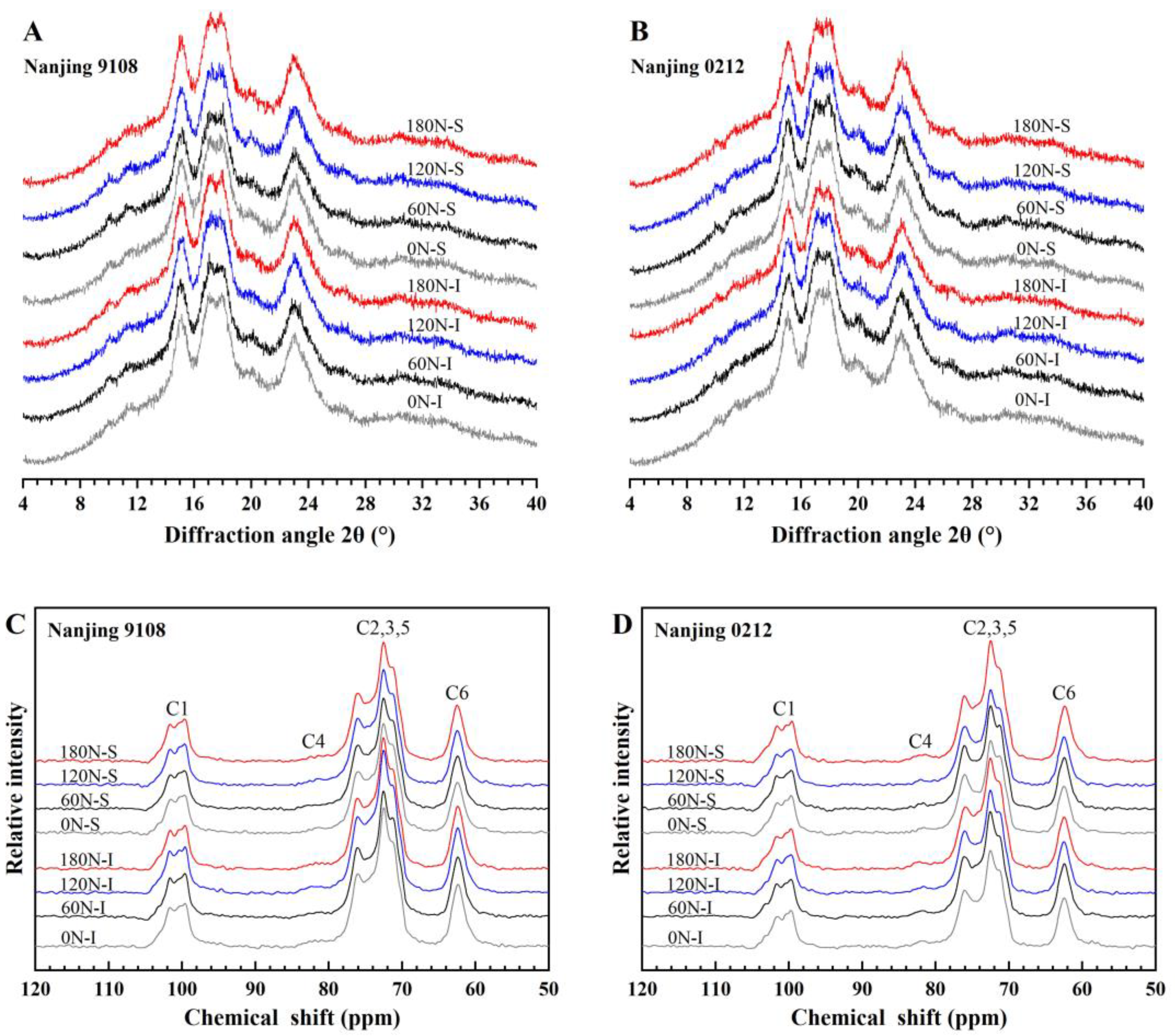
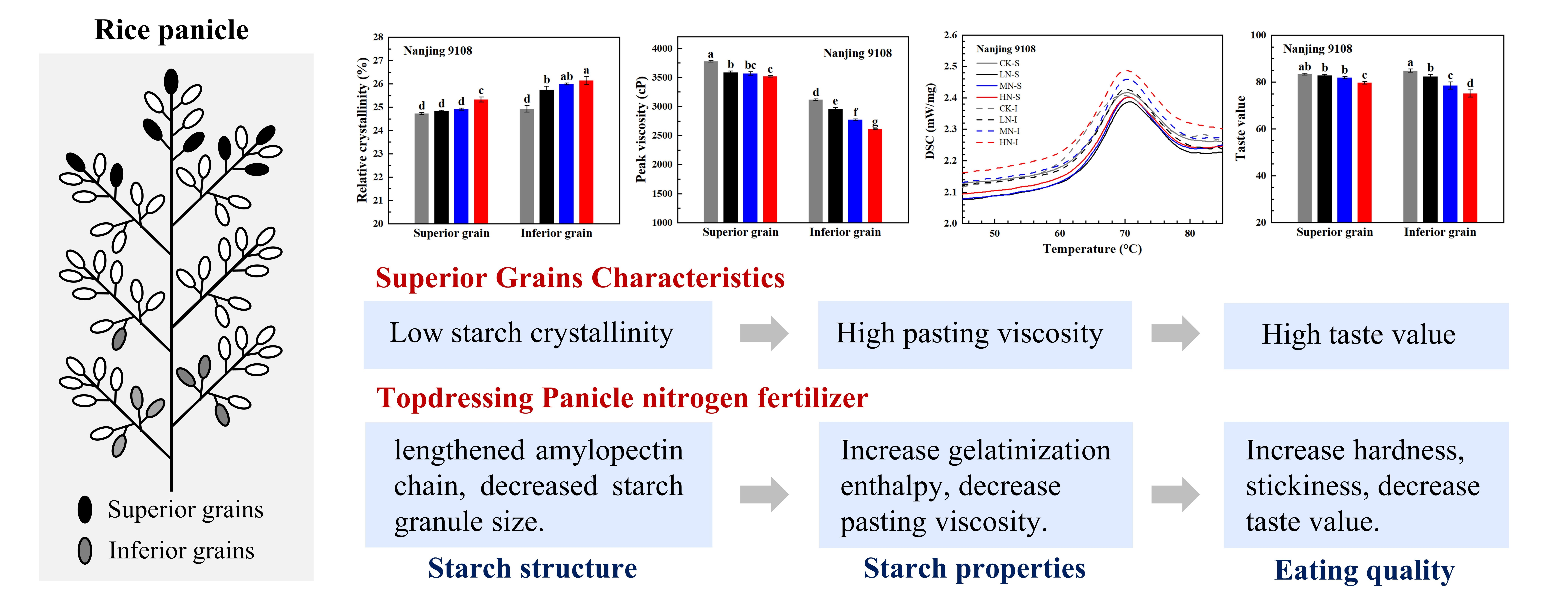
3.4. Starch Crystalline Structure
3.4.1. XRD Pattern and Relative Crystallinity
3.4.2. 13C CP/MAS NMR Analysis
3.5. Starch Solubility and Swelling Power
3.6. Starch Thermal Properties
3.7. Starch Pasting Properties
3.8. Instrumental Measurement of Eating Quality
3.8.1. Taste Value Measurement by NIR
3.8.2. Texture Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cao, L.Y.; Zhan, X.D.; Chen, S.G.; Feng, Y.; Wu, W.M.; Shen, X.H.; Cheng, S.H. Breeding methodology and practice of super rice in China. Rice Sci. 2010, 17, 87–93. [Google Scholar] [CrossRef]
- Cheng, B.; Jiang, Y.; Cao, C. Balance rice yield and eating quality by changing the traditional nitrogen management for sustainable production in China. J. Clean. Prod. 2021, 312, 127793. [Google Scholar] [CrossRef]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.Y.; Zhang, F.D.; Li, C.; Gilbert, R.G. Molecular structural differences between maize leaf and endosperm starches. Carbohydr. Polym. 2017, 161, 10–15. [Google Scholar] [CrossRef]
- Seung, D. Amylose in starch: Towards an understanding of biosynthesis, structure and function. New Phytologist. 2020, 228, 1490–1504. [Google Scholar] [CrossRef]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch—Composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Deng, F.; Yang, F.; Li, Q.P.; Zeng, Y.L.; Li, B.; Zhong, X.Y.; Lu, H.; Wang, L.; Chen, H.; Chen, Y.; et al. Differences in starch structural and physicochemical properties and texture characteristics of cooked rice between the main crop and ratoon rice. Food Hydrocoll. 2021, 116, 106643. [Google Scholar] [CrossRef]
- Li, C.; Hu, Y.; Gu, F.; Gong, B. Causal relations among starch fine molecular structure, lamellar/crystalline structure andin vitro digestion kinetics of native rice starch. Food Funct. 2021, 12, 682–695. [Google Scholar] [CrossRef]
- Zhou, T.Y.; Zhou, Q.; Li, E.P.; Yuan, L.M.; Wang, W.I.; Zhang, H.; Liu, L.J.; Wang, Z.Q.; Yang, J.C.; Gu, J.F. Effects of nitrogen fertilizer on structure and physicochemical properties of ‘super’ rice starch. Carbohydr. Polym. 2020, 239, 116237. [Google Scholar] [CrossRef]
- Li, H.Y.; Gilbert, R.G. Starch molecular structure: The basis for an improved understanding of cooked rice texture. Carbohydr. Polym. 2018, 195, 9–17. [Google Scholar] [CrossRef]
- Chen, F.; Lu, Y.; Pan, L.; Fan, X.; Li, Q.; Huang, L.; Zhao, D.; Zhang, C.; Liu, Q. The Underlying Physicochemical Properties and Starch Structures of indica Rice Grains with Translucent Endosperms under Low-Moisture Conditions. Foods 2022, 11, 1378. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, C.; Yan, Y.; Hu, Z.; Wang, K.; Zhou, J.; Zhou, Y.; Cao, L.; Wu, S. Influence of starch fine structure and storage proteins on the eating quality of rice varieties with similar amylose contents. J. Sci. Food Agric. 2021, 101, 3811–3818. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Lu, C.; Yang, Y.; Lu, Y.; Li, Q.; Huang, L.; Fan, X.; Liu, Q.; Zhang, C. The physicochemical properties of starch are affected by wxlv in indica rice. Foods 2021, 10, 3089. [Google Scholar] [CrossRef] [PubMed]
- Wani, A.A.; Singh, P.; Shah, M.A.; Schweiggert-Weisz, U.; Gul, K.; Wani, I.A. Rice starch diversity: Effects on structural, morphological, thermal, and physicochemical Properties—A review. Compr. Rev. Food Sci. F 2012, 11, 417–436. [Google Scholar] [CrossRef]
- Peng, Y.; Mao, B.; Zhang, C.; Shao, Y.; Wu, T.; Hu, L.; Hu, Y.; Tang, L.; Li, Y.; Tang, W.; et al. Influence of physicochemical properties and starch fine structure on the eating quality of hybrid rice with similar apparent amylose content. Food Chem. 2021, 353, 129461. [Google Scholar] [CrossRef]
- Zhu, Y.; Xu, D.; Ma, Z.; Chen, X.; Zhang, M.; Zhang, C.; Liu, G.; Wei, H.; Zhang, H. Differences in Eating Quality Attributes between Japonica Rice from the Northeast Region and Semiglutinous Japonica Rice from the Yangtze River Delta of China. Foods 2021, 10, 2770. [Google Scholar] [CrossRef]
- Li, H.Y.; Prakash, S.; Nicholson, T.M.; Fitzgerald, M.A.; Gilbert, R.G. Instrumental measurement of cooked rice texture by dynamic rheological testing and its relation to the fine structure of rice starch. Carbohydr. Polym. 2016, 146, 253–263. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.Y.; Song, N.Y.; Chen, Q.L.; Sun, H.Z.; Peng, T.; Huang, S.; Zhao, Q.Z. Response of grain-filling rate and grain quality of mid-season indica rice to nitrogen application. J. Integr. Agric. 2021, 20, 1465–1473. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J. Grain-filling problem in “super” rice. J. Exp. Bot. 2010, 61, 1–5. [Google Scholar] [CrossRef]
- Zhang, C.; Jiang, D.; Liu, F.; Cai, J.; Dai, T.; Cao, W. Starch granules size distribution in superior and inferior grains of wheat is related to enzyme activities and their gene expressions during grain filling. J. Cereal Sci. 2010, 51, 226–233. [Google Scholar] [CrossRef]
- Ma, Z.H.; Cheng, H.T.; Nitta, Y.; Aoki, N.; Chen, Y.; Chen, H.X.; Ohsugi, R.; Lyu, W.Y. Differences in viscosity of superior and inferior spikelets of japonica rice with various percentages of apparent amylose content. J. Agric. Food Chem. 2017, 65, 4237–4246. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.H.; Chen, H.X.; Lyu, W.Y.; Cheng, H.T.; Chen, Y.; Wang, Y.B. Comparison of the chemical and textural properties of germ-remaining soft rice grains from different spikelet positions. Cereal Chem. 2019, 96, 1137–1147. [Google Scholar] [CrossRef]
- Zhu, D.W.; Fang, C.Y.; Qian, Z.H.; Guo, B.W.; Huo, Z.Y. Differences in starch structure, physicochemical properties and texture characteristics in superior and inferior grains of rice varieties with different amylose contents. Food Hydrocoll. 2021, 110, 106170. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, Y.; Jia, B.; Wang, Y.; Wang, Y.; Xu, Q.; Li, R.; Wang, S.; Dou, F. Effects of cultivar, nitrogen rate, and planting density on Rice-Grain quality. Agronomy 2018, 8, 246. [Google Scholar] [CrossRef]
- Ayabe, S.; Kasai, M.; Ohishi, K.; Hatae, K. Textural properties and structures of starches from indica and japonica rice with similar amylose content. Food Sci. Technol. Res. 2009, 15, 299–306. [Google Scholar] [CrossRef]
- Lu, S.; Zhang, X.; Sun, H.; Cao, X.; Liu, J.; Zhang, Z.Z.Y.E. Relationship of grain protein content with cooking and eating quality as affected by nitrogen fertilizer at late growth stage for different types of rice varieties. Chin. J. Rice Sci. 2019, 33, 541–552. [Google Scholar] [CrossRef]
- Cagampang, G.B.; Perez, C.M.; Juliano, B.O. A gel consistency test for eating quality of rice. J. Sci. Food Agric. 1973, 24, 1589–1594. [Google Scholar] [CrossRef]
- Yang, Y.; Lin, G.Q.; Yu, X.R.; Wu, Y.F.; Xiong, F. Rice starch accumulation at different endosperm regions and physical properties under nitrogen treatment at panicle initiation stage. Int. J. Biol. Macromol. 2020, 160, 328–339. [Google Scholar] [CrossRef]
- Tao, K.; Yu, W.; Prakash, S.; Gilbert, R.G. High-amylose rice: Starch molecular structural features controlling cooked rice texture and preference. Carbohydr. Polym. 2019, 219, 251–260. [Google Scholar] [CrossRef]
- Liu, Z.H.; Cheng, F.M.; Cheng, W.D.; Zhang, G.P. Positional variations in phytic acid and protein content within a panicle of japonica rice. J. Cereal Sci. 2005, 41, 297–303. [Google Scholar] [CrossRef]
- Wu, A.C.; Li, E.; Gilbert, R.G. Exploring extraction/dissolution procedures for analysis of starch chain-length distributions. Carbohydr. Polym. 2014, 114, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.J.; Chen, L.; Zhao, Y.; Li, X.X. Structure and enzymatic resistivity of debranched high temperature–pressure treated high-amylose corn starch. J. Cereal Sci. 2013, 57, 348–355. [Google Scholar] [CrossRef]
- Tan, I.; Flanagan, B.M.; Halley, P.J.; Whittaker, A.K.; Gidley, M.J. A method for estimating the nature and relative proportions of amorphous, single, and Double-Helical components in starch granules by13 c CP/MAS NMR. Biomacromolecules 2007, 8, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.M.; Sun, H.Y.; Wang, C.G.; Ren, X.J.; Liu, H.F.; Zhang, Z.J. Effects of late-stage nitrogen fertilizer application on the starch structure and cooking quality of rice. J. Sci. Food Agric. 2018, 98, 2332–2340. [Google Scholar] [CrossRef]
- Hu, Q.; Liu, Q.Y.; Jiang, W.Q.; Qiu, S.; Wei, H.Y.; Zhang, H.C.; Liu, G.D.; Xing, Z.P.; Hu, Y.J.; Guo, B.W.; et al. Effects of mid-stage nitrogen application timing on the morphological structure and physicochemical properties of japonica rice starch. J. Sci. Food Agric. 2021, 101, 2463–2471. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, Y.; Chen, T.; Zhang, H.; Yang, J.; Zhang, J. Abscisic acid and the key enzymes and genes in sucrose-to-starch conversion in rice spikelets in response to soil drying during grain filling. Planta 2015, 241, 1091–1107. [Google Scholar] [CrossRef]
- Sun, T.; Tong, L.; Zhao, S.; Wang, H.; Han, Y.; Zhang, Z.; Jin, Z.Z.H.C. Effects of nitrogen fertilizer application on starch quality, activities and gene expression levels of related enzymes in rice endosperm. Chin. J. Rice Sci. 2018, 32, 475–484. [Google Scholar] [CrossRef]
- Xiong, F.; Wang, Z.; Gu, Y.; Chen, G.; Zhou, P. Effects of Nitrogen Application Time on Caryopsis Development and Grain Quality of Rice Variety Yangdao 6. Rice Sci. 2008, 15, 57–62. [Google Scholar] [CrossRef]
- Hu, Y.J.; Li, L.; Tian, J.Y.; Zhang, C.G.; Wang, J.; Yu, E.W.; Xing, Z.P.; Guo, B.W.; Wei, H.Y.; Huo, Z.Y.; et al. Effects of dynamic low temperature during the grain filling stage on starch morphological structure, physicochemical properties, and eating quality of softjaponica rice. Cereal Chem. 2020, 97, 540–550. [Google Scholar] [CrossRef]
- Hanashiro, I.; Abe, J.; Hizukuri, S. A periodic distribution of the chain length of amylopectin as revealed by high-performance anion-exchange chromatography. Carbohydr. Res. 1996, 283, 151–159. [Google Scholar] [CrossRef]
- Yang, X.Y.; Bi, J.G.; Gilbert, R.G.; Li, G.H.; Liu, Z.H.; Wang, S.H.; Ding, Y.F. Amylopectin chain length distribution in grains of japonica rice as affected by nitrogen fertilizer and genotype. J. Cereal Sci. 2016, 71, 230–238. [Google Scholar] [CrossRef]
- Mcmaugh, S.J.; Thistleton, J.L.; Anschaw, E.; Luo, J.; Konik-Rose, C.; Wang, H.; Huang, M.; Larroque, O.; Regina, A.; Jobling, S.A.; et al. Suppression of starch synthase I expression affects the granule morphology and granule size and fine structure of starch in wheat endosperm. J. Exp. Bot. 2014, 65, 2189–2201. [Google Scholar] [CrossRef] [PubMed]
- Fujita, N.; Toyosawa, Y.; Utsumi, Y.; Higuchi, T.; Hanashiro, I.; Ikegami, A.; Akuzawa, S.; Yoshida, M.; Mori, A.; Inomata, K.; et al. Characterization of pullulanase (PUL)-deficient mutants of rice (Oryza sativa L.) and the function of PUL on starch biosynthesis in the developing rice endosperm. J. Exp. Bot. 2009, 60, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Francisco, P.B.; Hosaka, Y.; Sato, A.; Sawada, T.; Kubo, A.; Fujita, N. Essential amino acids of starch synthase IIa differentiate amylopectin structure and starch quality between japonica and indica rice varieties. Plant Mol. Biol. 2005, 58, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Ando, H.; Watanabe, K.; Takeda, Y.; Mitsunaga, T. Physicochemical properties and structure of large, medium and small granule starches in fractions of normal barley endosperm. Carbohydr. Res. 2001, 330, 241–248. [Google Scholar] [CrossRef]
- Jenkins, P.J.; Donald, A.M. The influence of amylose on starch granule structure. Int. J. Biol. Macromol. 1995, 17, 315–321. [Google Scholar] [CrossRef]
- Zhu, D.W.; Zhang, H.C.; Guo, B.W.; Xu, K.; Dai, Q.G.; Wei, C.X.; Wei, H.Y.; Gao, H.; Hu, Y.J.; Cui, P.Y.; et al. Effect of nitrogen management on the structure and physicochemical properties of rice starch. J. Agric. Food Chem. 2016, 64, 8019–8025. [Google Scholar] [CrossRef]
- You, S.Y.; Oh, S.K.; Kim, H.S.; Chung, H.J. Influence of molecular structure on physicochemical properties and digestibility of normal rice starches. Int. J. Biol. Macromol. 2015, 77, 375–382. [Google Scholar] [CrossRef]
- Cooke, D.; Gidley, M.J. Loss of crystalline and molecular order during starch gelatinisation: Origin of the enthalpic transition. Carbohydr. Res. 1992, 227, 103–112. [Google Scholar] [CrossRef]
- Snape, C.E.; Morrison, W.R.; Maroto-Valer, M.M.; Karkalas, J.; Pethrick, R.A. Solid state 13C NMR investigation of lipid ligands in V-amylose inclusion complexes. Carbohydr. Polym. 1998, 36, 225–237. [Google Scholar] [CrossRef]
- Morrison, W.R.; Law, R.V.; Snape, C.E. Evidence for inclusion complexes of lipids with v-amylose in maize, rice and oat starches. J. Cereal Sci. 1993, 18, 107–109. [Google Scholar] [CrossRef]
- Chiotelli, E.; Le Meste, M. Effect of small and large wheat starch granules on thermomechanical behavior of starch. Cereal Chem. 2002, 79, 286–293. [Google Scholar] [CrossRef]
- Kaur, L.; Singh, J.; Mccarthy, O.J.; Singh, H. Physico-chemical, rheological and structural properties of fractionated potato starches. J. Food Eng. 2007, 82, 383–394. [Google Scholar] [CrossRef]
- Tao, K.; Li, C.; Yu, W.; Gilbert, R.G.; Li, E. How amylose molecular fine structure of rice starch affects functional properties. Carbohydr. Polym. 2019, 204, 24–31. [Google Scholar] [CrossRef]
- Chung, H.J.; Liu, Q.; Lee, L.; Wei, D.Z. Relationship between the structure, physicochemical properties and in vitro digestibility of rice starches with different amylose contents. Food Hydrocoll. 2011, 25, 968–975. [Google Scholar] [CrossRef]
- Kong, X.; Bertoft, E.; Bao, J.; Corke, H. Molecular structure of amylopectin from amaranth starch and its effect on physicochemical properties. Int. J. Biol. Macromol. 2008, 43, 377–382. [Google Scholar] [CrossRef]
- Li, C.; Gong, B. Insights into chain-length distributions of amylopectin and amylose molecules on the gelatinization property of rice starches. Int. J. Biol. Macromol. 2020, 155, 721–729. [Google Scholar] [CrossRef]
- Kong, X.; Zhu, P.; Sui, Z.; Bao, J. Physicochemical properties of starches from diverse rice cultivars varying in apparent amylose content and gelatinisation temperature combinations. Food Chem. 2015, 172, 433–440. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, L.; Shao, K.; Gu, M.; Liu, Q. Toward underlying reasons for rice starches having low viscosity and high amylose: Physiochemical and structural characteristics. J. Sci. Food Agric. 2013, 93, 1543–1551. [Google Scholar] [CrossRef]
- Srichuwong, S.; Sunarti, T.C.; Mishima, T.; Isono, N.; Hisamatsu, M. Starches from different botanical sources II: Contribution of starch structure to swelling and pasting properties. Carbohydr. Polym. 2005, 62, 25–34. [Google Scholar] [CrossRef]
- Hu, Y.J.; Cong, S.M.; Zhang, H.C. Comparison of the grain quality and starch physicochemical properties between japonica rice cultivars with different contents of amylose, as affected by nitrogen fertilization. Agriculture 2021, 11, 616. [Google Scholar] [CrossRef]
- Horibata, T.; Nakamoto, M.; Fuwa, H.; Inouchi, N. Structural and physicochemical characteristics of endosperm starches of rice cultivars recently bred in Japan. J. Appl. Glycosci. 2004, 51, 303–313. [Google Scholar] [CrossRef]
- Saleh, M.I.; Meullenet, J. Effect of protein disruption using proteolytic treatment on cooked rice texture properties. J. Texture Stud. 2007, 38, 423–437. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Position | PNF (kg ha−1) | Starch Content (%) | Amylose Content (%) | Amylopectin Content (%) | Amylose/Amylopectin |
|---|---|---|---|---|---|---|
| Nanjing 9108 | Superior grain | 0 | 82.91 ± 1.23 b | 11.86 ± 0.07 a | 70.76 ± 0.93 c | 0.168 ± 0.002 a |
| 60 | 81.48 ± 1.34 b | 11.53 ± 0.20 a | 69.53 ± 0.50 de | 0.166 ± 0.001 a | ||
| 120 | 79.07 ± 0.97 cd | 10.35 ± 0.11 b | 68.55 ± 0.92 e | 0.151 ± 0.002 b | ||
| 180 | 75.46 ± 0.59 e | 9.35 ± 0.38 cd | 66.00 ± 0.64 f | 0.142 ± 0.001 c | ||
| Inferior grain | 0 | 85.27 ± 1.53 a | 10.37 ± 0.33 b | 75.44 ± 0.74 a | 0.138 ± 0.001 c | |
| 60 | 82.58 ± 1.35 b | 9.75 ± 0.08 bc | 72.66 ± 1.23 b | 0.134 ± 0.002 d | ||
| 120 | 79.75 ± 0.70 c | 9.39 ± 0.15 cd | 70.48 ± 0.67 cd | 0.133 ± 0.001 d | ||
| 180 | 77.96 ± 0.77 d | 8.92 ± 0.16 d | 68.79 ± 0.32 e | 0.130 ± 0.001 d | ||
| Nanjing 0212 | Superior grain | 0 | 83.97 ± 0.57 b | 20.86 ± 0.27 a | 63.14 ± 0.47 e | 0.330 ± 0.002 a |
| 60 | 81.84 ± 0.29 c | 20.12 ± 0.36 b | 61.85 ± 0.38 f | 0.325 ± 0.002 b | ||
| 120 | 81.08 ± 0.34 cd | 19.53 ± 0.23 b | 61.47 ± 0.13 f | 0.318 ± 0.001 c | ||
| 180 | 79.91 ± 0.78 d | 18.79 ± 0.55 c | 61.00 ± 0.89 f | 0.308 ± 0.004 d | ||
| Inferior grain | 0 | 86.67 ± 0.44 a | 16.69 ± 0.17 d | 70.05 ± 0.42 a | 0.238 ± 0.001 e | |
| 60 | 83.76 ± 0.44 b | 16.08 ± 0.12 d | 67.78 ± 0.35 b | 0.237 ± 0.001 e | ||
| 120 | 82.20 ± 0.81 c | 15.29 ± 0.04 e | 66.73 ± 0.64 c | 0.229 ± 0.002 f | ||
| 180 | 80.23 ± 0.93 d | 14.49 ± 0.56 f | 65.24 ± 0.26 d | 0.222 ± 0.001 g |
| Variety | Position | PNF (kg ha−1) | Total Protein Content (%) | Albumin Content (mg g−1) | Globulin Content (mg g−1) | Prolamin Content (mg g−1) | Glutelin Content (mg g−1) | Crude Fat Content (%) |
|---|---|---|---|---|---|---|---|---|
| Nanjing 9108 | Superior grain | 0 | 5.94 ± 0.03 h | 3.45 ± 0.10 f | 4.32 ± 0.04 d | 4.10 ± 0.02 f | 33.44 ± 0.27 f | 0.65 ± 0.02 c |
| 60 | 6.64 ± 0.05 f | 3.57 ± 0.08 f | 4.44 ± 0.05 d | 4.51 ± 0.04 d | 38.56 ± 1.42 e | 0.45 ± 0.01 d | ||
| 120 | 7.04 ± 0.01 e | 3.77 ± 0.09 e | 4.61 ± 0.05 c | 4.86 ± 0.07 c | 42.09 ± 0.75 d | 0.42 ± 0.04 d | ||
| 180 | 7.19 ± 0.03 d | 4.03 ± 0.05 d | 4.74 ± 0.06 c | 5.35 ± 0.03 b | 43.75 ± 0.98 cd | 0.42 ± 0.02 d | ||
| Inferior grain | 0 | 6.54 ± 0.02 g | 4.39 ± 0.16 c | 4.41 ± 0.04 d | 4.57 ± 0.05 e | 35.03 ± 0.39 f | 1.28 ± 0.03 a | |
| 60 | 7.49 ± 0.06 c | 4.52 ± 0.05 bc | 4.71 ± 0.06 c | 4.67 ± 0.03 cd | 45.49 ± 0.67 bc | 0.84 ± 0.04 b | ||
| 120 | 7.90 ± 0.07 b | 4.66 ± 0.07 b | 4.89 ± 0.07 b | 4.98 ± 0.02 bc | 46.27 ± 0.46 b | 0.83 ± 0.03 b | ||
| 180 | 8.29 ± 0.00 a | 4.90 ± 0.10 a | 5.09 ± 0.05 a | 5.43 ± 0.06 a | 48.68 ± 0.72 a | 0.82 ± 0.02 b | ||
| Nanjing 0212 | Superior grain | 0 | 5.69 ± 0.01 h | 2.97 ± 0.06 d | 4.16 ± 0.04 e | 4.12 ± 0.04 f | 37.04 ± 0.70 e | 0.52 ± 0.03 d |
| 60 | 6.30 ± 0.00 f | 3.13 ± 0.06 d | 4.28 ± 0.06 de | 4.80 ± 0.10 e | 37.53 ± 0.51 e | 0.39 ± 0.03 e | ||
| 120 | 6.62 ± 0.01 e | 3.42 ± 0.08 c | 4.40 ± 0.08 d | 5.02 ± 0.05 d | 41.44 ± 0.31 c | 0.34 ± 0.03 f | ||
| 180 | 6.85 ± 0.01 d | 3.72 ± 0.16 b | 4.71 ± 0.10 b | 5.22 ± 0.10 a | 44.94 ± 0.90 b | 0.33 ± 0.02 f | ||
| Inferior grain | 0 | 6.05 ± 0.00 g | 3.63 ± 0.05 b | 4.55 ± 0.03 c | 4.47 ± 0.04 f | 38.54 ± 0.69 de | 1.79 ± 0.02 a | |
| 60 | 7.59 ± 0.00 c | 3.75 ± 0.08 b | 4.57 ± 0.11 c | 4.94 ± 0.06 d | 39.92 ± 0.30 cd | 1.28 ± 0.02 b | ||
| 120 | 8.44 ± 0.01 b | 3.97 ± 0.10 a | 5.04 ± 0.05 a | 5.09 ± 0.09 c | 48.38 ± 0.58 a | 1.18 ± 0.03 c | ||
| 180 | 8.58 ± 0.04 a | 4.12 ± 0.08 a | 5.13 ± 0.09 a | 5.48 ± 0.06 a | 49.88 ± 1.00 a | 1.16 ± 0.02 c |
| Variety | Position | PNF (kg ha−1) | Small Starch Granules ≤2 μm (%) | Middle Starch Granules 2–5 μm (%) | Large Starch Granules ≥5 μm (%) | Volume Mean Diameter (μm) |
|---|---|---|---|---|---|---|
| Nanjing 9108 | Superior grain | 0 | 12.72 ± 0.03 c | 34.11 ± 0.06 e | 53.17 ± 0.09 d | 5.33 ± 0.01 d |
| 60 | 12.60 ± 0.05 c | 36.31 ± 0.01 a | 51.10 ± 0.06 g | 5.22 ± 0.00 g | ||
| 120 | 13.47 ± 0.11 a | 34.26 ± 0.02 d | 52.27 ± 0.13 e | 5.27 ± 0.01 f | ||
| 180 | 13.56 ± 0.12 a | 35.89 ± 0.13 b | 50.55 ± 0.25 h | 5.17 ± 0.02 h | ||
| Inferior grain | 0 | 10.97 ± 0.08 e | 30.04 ± 0.04 h | 58.99 ± 0.13 a | 5.78 ± 0.01 a | |
| 60 | 12.29 ± 0.09 d | 31.28 ± 0.05 g | 56.43 ± 0.13 b | 5.58 ± 0.01 b | ||
| 120 | 13.14 ± 0.12 b | 34.84 ± 0.04 c | 52.02 ± 0.15 f | 5.31 ± 0.01 e | ||
| 180 | 12.59 ± 0.14 c | 32.26 ± 0.04 f | 55.15 ± 0.17 c | 5.50 ± 0.01 c | ||
| Nanjing 0212 | Superior grain | 0 | 13.54 ± 0.11 d | 34.51 ± 0.08 h | 51.95 ± 0.11 a | 5.22 ± 0.01 a |
| 60 | 14.01 ± 0.07 c | 36.41 ± 0.02 g | 49.58 ± 0.06 b | 5.07 ± 0.00 b | ||
| 120 | 14.22 ± 0.06 b | 37.80 ± 0.05 d | 47.98 ± 0.11 e | 4.99 ± 0.01 c | ||
| 180 | 14.45 ± 0.02 a | 36.57 ± 0.03 f | 48.98 ± 0.01 c | 5.06 ± 0.00 b | ||
| Inferior grain | 0 | 13.96 ± 0.02 c | 37.37 ± 0.04 e | 48.67 ± 0.05 d | 5.06 ± 0.00 b | |
| 60 | 14.09 ± 0.17 bc | 38.82 ± 0.03 c | 47.09 ± 0.16 f | 4.99 ± 0.01 c | ||
| 120 | 14.40 ± 0.06 a | 42.32 ± 0.03 b | 43.28 ± 0.06 g | 4.77 ± 0.00 d | ||
| 180 | 14.44 ± 0.02 a | 42.53 ± 0.05 a | 43.02 ± 0.07 h | 4.77 ± 0.01 d |
| Variety | Position | PNF (kg ha−1) | A Chain (%) | B1 Chain (%) | B2 Chain (%) | B3 Chain (%) | (A + B1)/(B2 + B3) | Amylopectin Average DP |
|---|---|---|---|---|---|---|---|---|
| Nanjing 9108 | Superior grain | 0 | 28.60 ± 0.10 a | 47.21 ± 0.21 a | 10.83 ± 0.08 c | 13.36 ± 0.04 e | 3.13 ± 0.01 a | 20.76 ± 0.05 e |
| 60 | 27.16 ± 0.13 c | 46.82 ± 0.22 a | 11.36 ± 0.13 ab | 14.66 ± 0.06 cd | 2.84 ± 0.02 bc | 21.61 ± 0.05 c | ||
| 120 | 26.26 ± 0.12 f | 46.60 ± 0.21 a | 11.53 ± 0.14 a | 15.61 ± 0.19 ab | 2.72 ± 0.01 d | 22.12 ± 0.07 a | ||
| 180 | 26.51 ± 0.20 ef | 46.50 ± 0.16 a | 11.28 ± 0.03 ab | 15.76 ± 0.20 a | 2.70 ± 0.01 d | 22.12 ± 0.12 a | ||
| Inferior grain | 0 | 28.19 ± 0.10 b | 46.43 ± 0.16 a | 10.93 ± 0.04 c | 14.46 ± 0.11 d | 2.94 ± 0.01 b | 21.40 ± 0.14 d | |
| 60 | 26.94 ± 0.02 cd | 46.98 ± 0.04 a | 10.96 ± 0.12 c | 15.12 ± 0.06 bc | 2.83 ± 0.01 bc | 21.80 ± 0.01 bc | ||
| 120 | 26.70 ± 0.03 de | 46.96 ± 0.24 a | 11.11 ± 0.04 bc | 15.23 ± 0.15 ab | 2.80 ± 0.03 cd | 21.84 ± 0.13 b | ||
| 180 | 26.53 ± 0.15 ef | 47.01 ± 0.11 a | 11.08 ± 0.18 bc | 15.39 ± 0.22 ab | 2.78 ± 0.03 cd | 21.93 ± 0.11 ab | ||
| Nanjing 0212 | Superior grain | 0 | 27.26 ± 0.07 b | 47.06 ± 0.25 a | 10.92 ± 0.16 ab | 14.76 ± 0.12 cd | 2.89 ± 0.02 ab | 21.62 ± 0.07 d |
| 60 | 26.71 ± 0.02 cd | 47.06 ± 0.18 a | 11.01 ± 0.12 a | 15.21 ± 0.15 bc | 2.81 ± 0.02 bc | 21.86 ± 0.10 bc | ||
| 120 | 26.28 ± 0.07 e | 47.02 ± 0.19 a | 11.08 ± 0.06 a | 15.61 ± 0.14 ab | 2.75 ± 0.01 c | 22.05 ± 0.08 ab | ||
| 180 | 26.36 ± 0.08 e | 46.82 ± 0.04 a | 11.04 ± 0.03 a | 15.78 ± 0.05 a | 2.73 ± 0.01 c | 22.13 ± 0.04 a | ||
| Inferior grain | 0 | 27.85 ± 0.15 a | 47.01 ± 0.07 a | 10.68 ± 0.08 b | 14.51 ± 0.19 d | 2.97 ± 0.04 a | 21.53 ± 0.03 d | |
| 60 | 26.77 ± 0.04 c | 47.17 ± 0.23 a | 10.96 ± 0.05 ab | 15.10 ± 0.14 bc | 2.84 ± 0.02 bc | 21.71 ± 0.07 cd | ||
| 120 | 26.88 ± 0.01 c | 46.80 ± 0.13 a | 10.88 ± 0.08 ab | 15.44 ± 0.21 ab | 2.80 ± 0.02 bc | 21.90 ± 0.10 bc | ||
| 180 | 26.44 ± 0.08 de | 47.03 ± 0.13 a | 11.02 ± 0.02 a | 15.52 ± 0.10 ab | 2.76 ± 0.01 c | 21.94 ± 0.09 ab |
| Variety | Position | PNF (kg ha−1) | Relative Crystallinity (%) | Amorphous (%) | Single Helix (%) | Double Helix (%) |
|---|---|---|---|---|---|---|
| Nanjing 9108 | Superior grain | 0 | 24.72 ± 0.05 d | 57.04 ± 0.40 a | 3.86 ± 0.12 bc | 39.10 ± 0.49 e |
| 60 | 24.82 ± 0.04 d | 56.81 ± 0.31 a | 3.82 ± 0.06 bc | 39.37 ± 0.25 e | ||
| 120 | 24.91 ± 0.04 d | 56.56 ± 0.12 a | 3.07 ± 0.08 d | 40.37 ± 0.07 d | ||
| 180 | 25.33 ± 0.11 c | 55.82 ± 0.14 b | 2.90 ± 0.06 d | 41.28 ± 0.20 c | ||
| Inferior grain | 0 | 24.92 ± 0.14 d | 54.63 ± 0.45 c | 4.41 ± 0.09 a | 40.96 ± 0.36 cd | |
| 60 | 25.74 ± 0.15 b | 54.41 ± 0.21 c | 4.12 ± 0.05 ab | 41.47 ± 0.21 c | ||
| 120 | 25.99 ± 0.04 ab | 52.82 ± 0.10 d | 3.98 ± 0.24 bc | 43.19 ± 0.17 b | ||
| 180 | 26.14 ± 0.17 a | 52.26 ± 0.35 d | 3.68 ± 0.01 c | 44.06 ± 0.35 a | ||
| Nanjing 0212 | Superior grain | 0 | 23.35 ± 0.16 f | 64.27 ± 0.07 a | 3.53 ± 0.13 b | 32.21 ± 0.14 f |
| 60 | 23.57 ± 0.11 ef | 63.20 ± 0.15 b | 3.34 ± 0.09 b | 33.46 ± 0.17 e | ||
| 120 | 23.99 ± 0.07 cd | 62.68 ± 0.12 b | 3.01 ± 0.35 c | 34.32 ± 0.39 d | ||
| 180 | 24.46 ± 0.11 b | 59.18 ± 0.56 d | 2.92 ± 0.15 c | 37.90 ± 0.41 b | ||
| Inferior grain | 0 | 23.82 ± 0.19 de | 63.02 ± 0.22 b | 4.23 ± 0.34 a | 32.74 ± 0.48 f | |
| 60 | 24.27 ± 0.12 bc | 62.78 ± 0.07 b | 3.59 ± 0.09 b | 33.64 ± 0.08 e | ||
| 120 | 24.90 ± 0.09 a | 61.26 ± 0.25 c | 3.42 ± 0.41 b | 35.32 ± 0.31 c | ||
| 180 | 25.27 ± 0.15 a | 57.86 ± 0.32 e | 3.34 ± 0.07 b | 38.81 ± 0.36 a |
| Variety | Position | PNF (kg ha−1) | △Hgel (J·g−1) | TO (°C) | TP (°C) | TC (°C) | △Hret (J·g−1) | R (%) |
|---|---|---|---|---|---|---|---|---|
| Nanjing 9108 | Superior grain | 0 | 10.20 ± 0.07 e | 62.80 ± 0.17 c | 70.33 ± 0.15 bc | 77.53 ± 0.12 bc | 2.83 ± 0.03 a | 27.75 ± 0.49 a |
| 60 | 11.67 ± 0.03 c | 63.37 ± 0.15 b | 70.57 ± 0.06 ab | 77.80 ± 0.20 ab | 2.72 ± 0.04 b | 23.31 ± 0.35 c | ||
| 120 | 12.00 ± 0.03 b | 63.73 ± 0.12 a | 70.70 ± 0.17 a | 77.93 ± 0.21 a | 2.69 ± 0.08 b | 22.44 ± 0.62 d | ||
| 180 | 11.73 ± 0.06 c | 63.80 ± 0.20 a | 70.70 ± 0.20 a | 78.00 ± 0.26 a | 2.56 ± 0.03 c | 21.85 ± 0.28 de | ||
| Inferior grain | 0 | 10.61 ± 0.05 d | 59.63 ± 0.32 e | 69.93 ± 0.15 d | 77.23 ± 0.15 c | 2.72 ± 0.05 b | 25.67 ± 0.37 b | |
| 60 | 11.94 ± 0.08 b | 62.17 ± 0.15 d | 70.17 ± 0.15 cd | 77.53 ± 0.21 bc | 2.58 ± 0.06 c | 21.63 ± 0.44 e | ||
| 120 | 12.14 ± 0.04 a | 62.53 ± 0.15 c | 70.43 ± 0.06 b | 77.67 ± 0.32 ab | 2.53 ± 0.04 c | 20.84 ± 0.28 f | ||
| 180 | 11.95 ± 0.08 b | 62.57 ± 0.12 c | 70.50 ± 0.10 ab | 77.77 ± 0.21 ab | 2.38 ± 0.01 d | 19.94 ± 0.19 g | ||
| Nanjing 0212 | Superior grain | 0 | 10.71 ± 0.05 f | 63.47 ± 0.12 a | 70.60 ± 0.10 a | 77.63 ± 0.12 a | 3.20 ± 0.08 a | 29.92 ± 0.61 a |
| 60 | 11.31 ± 0.08 e | 63.23 ± 0.06 ab | 70.43 ± 0.12 a | 77.37 ± 0.06 ab | 2.95 ± 0.05 c | 26.06 ± 0.54 c | ||
| 120 | 11.84 ± 0.08 c | 63.10 ± 0.10 bc | 70.17 ± 0.06 b | 77.23 ± 0.25 b | 2.90 ± 0.06 cd | 24.50 ± 0.69 d | ||
| 180 | 11.87 ± 0.05 c | 61.93 ± 0.21 d | 69.20 ± 0.30 d | 76.83 ± 0.25 c | 2.57 ± 0.03 f | 21.66 ± 0.36 f | ||
| Inferior grain | 0 | 11.31 ± 0.07 e | 62.90 ± 0.10 c | 69.57 ± 0.06 c | 76.63 ± 0.15 cd | 3.09 ± 0.04 b | 27.33 ± 0.44 b | |
| 60 | 11.61 ± 0.08 d | 61.27 ± 0.21 e | 69.03 ± 0.25 de | 76.30 ± 0.20 de | 2.83 ± 0.05 d | 24.37 ± 0.46 d | ||
| 120 | 12.00 ± 0.07 b | 61.23 ± 0.12 e | 69.00 ± 0.00 de | 76.17 ± 0.06 e | 2.75 ± 0.05 e | 22.89 ± 0.51 e | ||
| 180 | 12.60 ± 0.05 a | 61.00 ± 0.10 e | 68.90 ± 0.10 e | 75.97 ± 0.15 e | 2.47 ± 0.01 g | 19.58 ± 0.17 g |
| Variety | Position | PNF (kg ha−1) | Peak Viscosity (cP) | Hot Viscosity (cP) | Breakdown (cP) | Final Viscosity (cP) | Setback (cP) | Pasting Temperature (°C) |
|---|---|---|---|---|---|---|---|---|
| Nanjing 9108 | Superior grain | 0 | 3776 ± 11 a | 2509 ± 12 a | 1267 ± 14 a | 3003± 9 a | −773 ± 19 d | 76.33 ± 0.31 a |
| 60 | 3584 ± 27 b | 2298 ± 30 b | 1286 ± 18 a | 2789 ± 43 b | −795 ± 21 de | 76.80 ± 0.05 a | ||
| 120 | 3566 ± 30 bc | 2274 ± 49 b | 1292 ± 28 a | 2721 ± 33 c | −844 ± 29 f | 76.78 ± 0.06 a | ||
| 180 | 3518 ± 15 c | 2267 ± 24 b | 1251 ± 14 a | 2701 ± 17 c | −817 ± 13 ef | 76.80 ± 0.05 a | ||
| Inferior grain | 0 | 3120 ± 12 d | 2008 ± 32 c | 1112 ± 21 bc | 2582 ± 51 d | −537 ± 38 a | 75.52 ± 0.35 b | |
| 60 | 2956 ± 28 e | 1822 ± 11 d | 1133 ± 18 b | 2264 ± 15 e | −692 ± 15 c | 76.30 ± 0.38 a | ||
| 120 | 2774 ± 11 f | 1634 ± 31 e | 1140 ± 27 b | 2067 ± 30 f | −707 ± 20 c | 76.62 ± 0.28 a | ||
| 180 | 2612 ± 15 g | 1523 ± 27 f | 1089 ± 26 c | 1988 ± 51 g | −624 ± 37 b | 76.70 ± 0.09 a | ||
| Nanjing 0212 | Superior grain | 0 | 3478 ± 11 a | 2769 ± 20 a | 710 ± 13 a | 3794 ± 24 a | 316 ± 14 ab | 77.38 ± 0.03 d |
| 60 | 3212 ± 41 b | 2491 ± 51 b | 721 ± 14 a | 3475 ± 30 b | 263 ± 12 cd | 77.60 ± 0.48 d | ||
| 120 | 2992 ± 26 c | 2266 ± 41 d | 726 ± 18 a | 3228 ± 43 d | 237 ± 17 d | 77.73 ± 0.33 cd | ||
| 180 | 2925 ± 24 d | 2219 ± 25 d | 706 ± 16 a | 3207 ± 12 d | 282 ± 12 bc | 77.67 ± 0.45 d | ||
| Inferior grain | 0 | 3038 ± 11 c | 2387 ± 28 c | 651 ± 33 b | 3397 ± 19 c | 359 ± 10 a | 78.18 ± 0.03 bc | |
| 60 | 2360 ± 61 e | 1671 ± 66 e | 689 ± 30 ab | 2677 ± 63 e | 317 ± 11 ab | 78.60 ± 0.09 b | ||
| 120 | 2179 ± 47 f | 1471 ± 32 f | 708 ± 16 a | 2478 ± 37 f | 298 ± 13 bc | 79.43 ± 0.46 a | ||
| 180 | 2007 ± 32 g | 1348 ± 20 g | 660 ± 14 b | 2353 ± 24 g | 345 ± 11 a | 79.70 ± 0.05 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Chen, Y.; Zhao, C.; Liu, G.; Shi, Y.; Zhao, L.; Wang, Y.; Wang, W.; Xu, K.; Li, G.; et al. The Starch Physicochemical Properties between Superior and Inferior Grains of Japonica Rice under Panicle Nitrogen Fertilizer Determine the Difference in Eating Quality. Foods 2022, 11, 2489. https://doi.org/10.3390/foods11162489
Jiang Y, Chen Y, Zhao C, Liu G, Shi Y, Zhao L, Wang Y, Wang W, Xu K, Li G, et al. The Starch Physicochemical Properties between Superior and Inferior Grains of Japonica Rice under Panicle Nitrogen Fertilizer Determine the Difference in Eating Quality. Foods. 2022; 11(16):2489. https://doi.org/10.3390/foods11162489
Chicago/Turabian StyleJiang, Yan, Yue Chen, Can Zhao, Guangming Liu, Yi Shi, Lingtian Zhao, Yuan Wang, Weiling Wang, Ke Xu, Guohui Li, and et al. 2022. "The Starch Physicochemical Properties between Superior and Inferior Grains of Japonica Rice under Panicle Nitrogen Fertilizer Determine the Difference in Eating Quality" Foods 11, no. 16: 2489. https://doi.org/10.3390/foods11162489
APA StyleJiang, Y., Chen, Y., Zhao, C., Liu, G., Shi, Y., Zhao, L., Wang, Y., Wang, W., Xu, K., Li, G., Dai, Q., & Huo, Z. (2022). The Starch Physicochemical Properties between Superior and Inferior Grains of Japonica Rice under Panicle Nitrogen Fertilizer Determine the Difference in Eating Quality. Foods, 11(16), 2489. https://doi.org/10.3390/foods11162489