
Dietary Supplementation with Sea Buckthorn Berry Puree Alters Plasma Metabolomic Profile and Gut Microbiota Composition in Hypercholesterolemia Population



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Study Participants and Intervention
2.3. Biochemical Parameters
2.4. 1H NMR Metabolomic Analysis
2.5. Collection of Fecal Samples and Analysis of Gut Microbiota
2.6. Statistical Analysis
3. Results
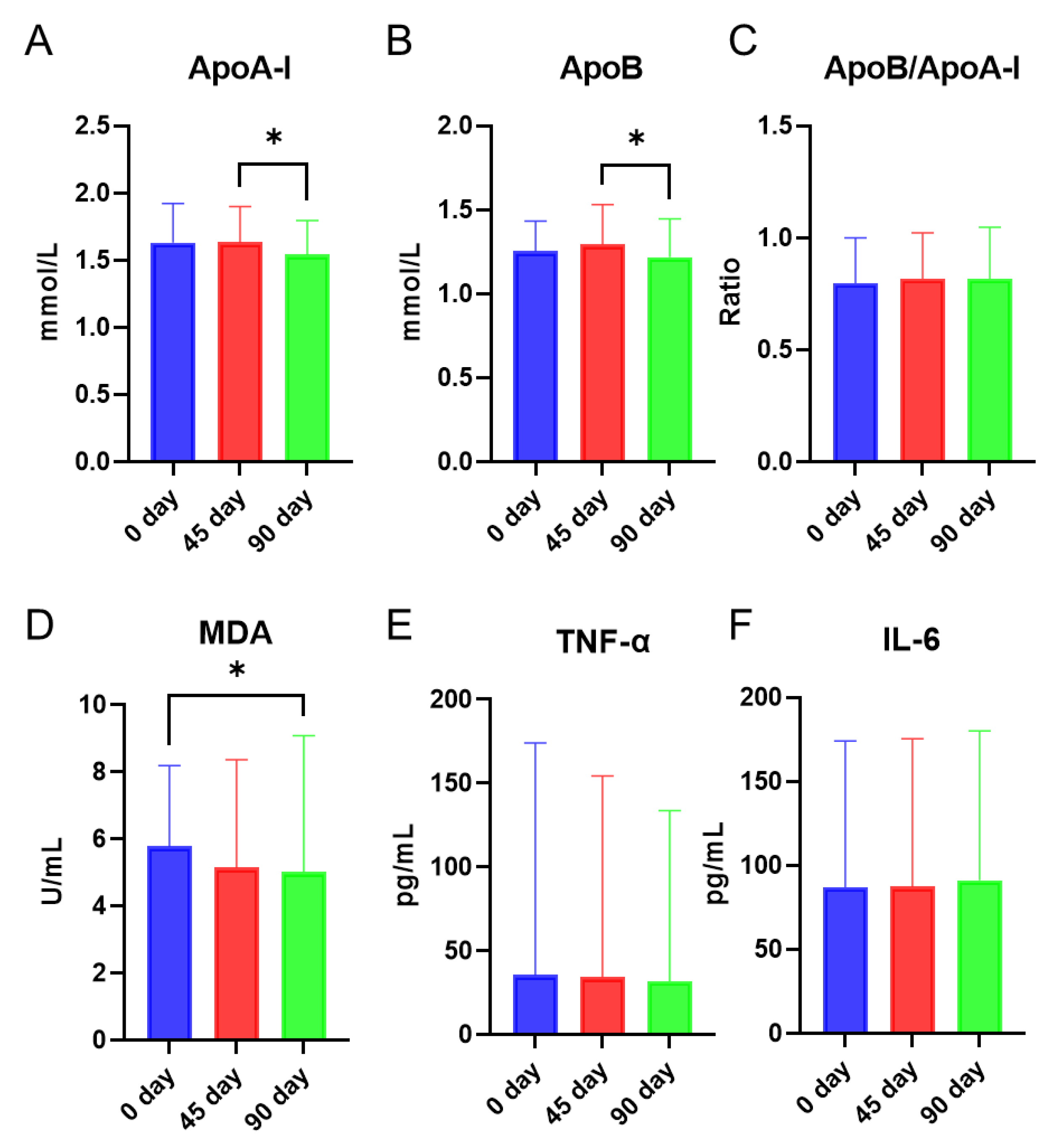
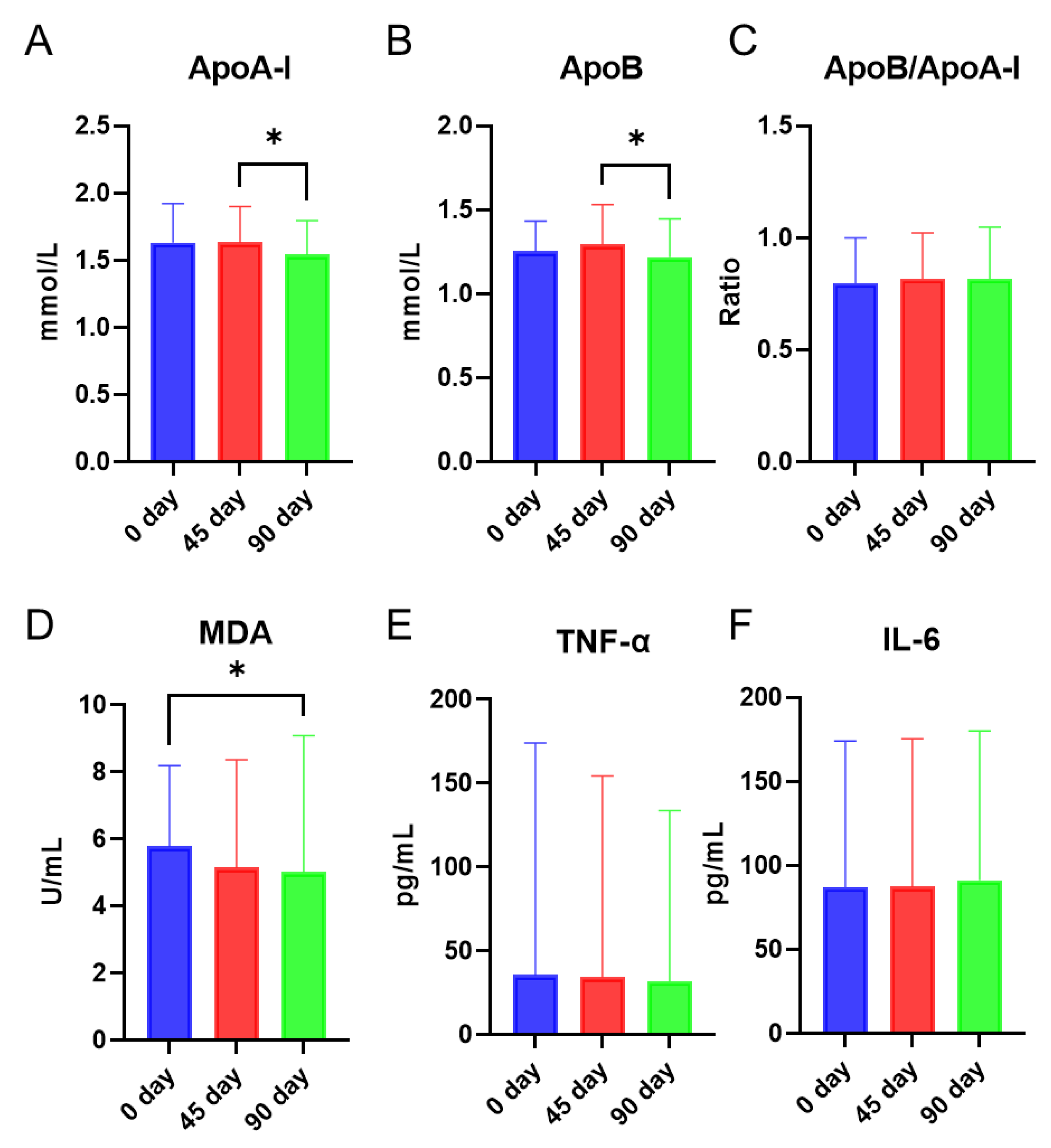
3.1. Effects of Sea Buckthorn Puree on Lipid Biomarkers, Lipid Peroxidation Product, and Inflammatory Markers
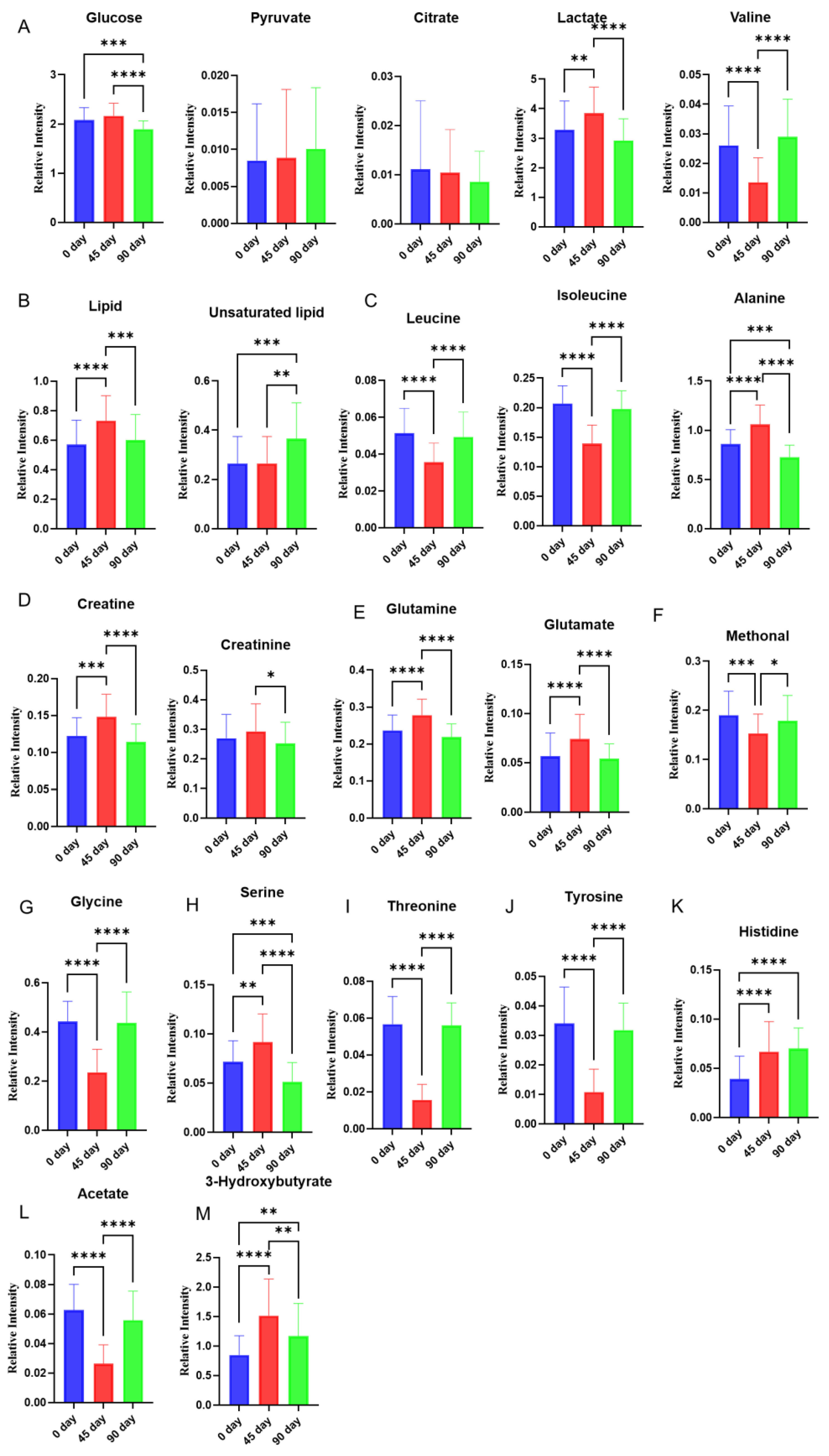
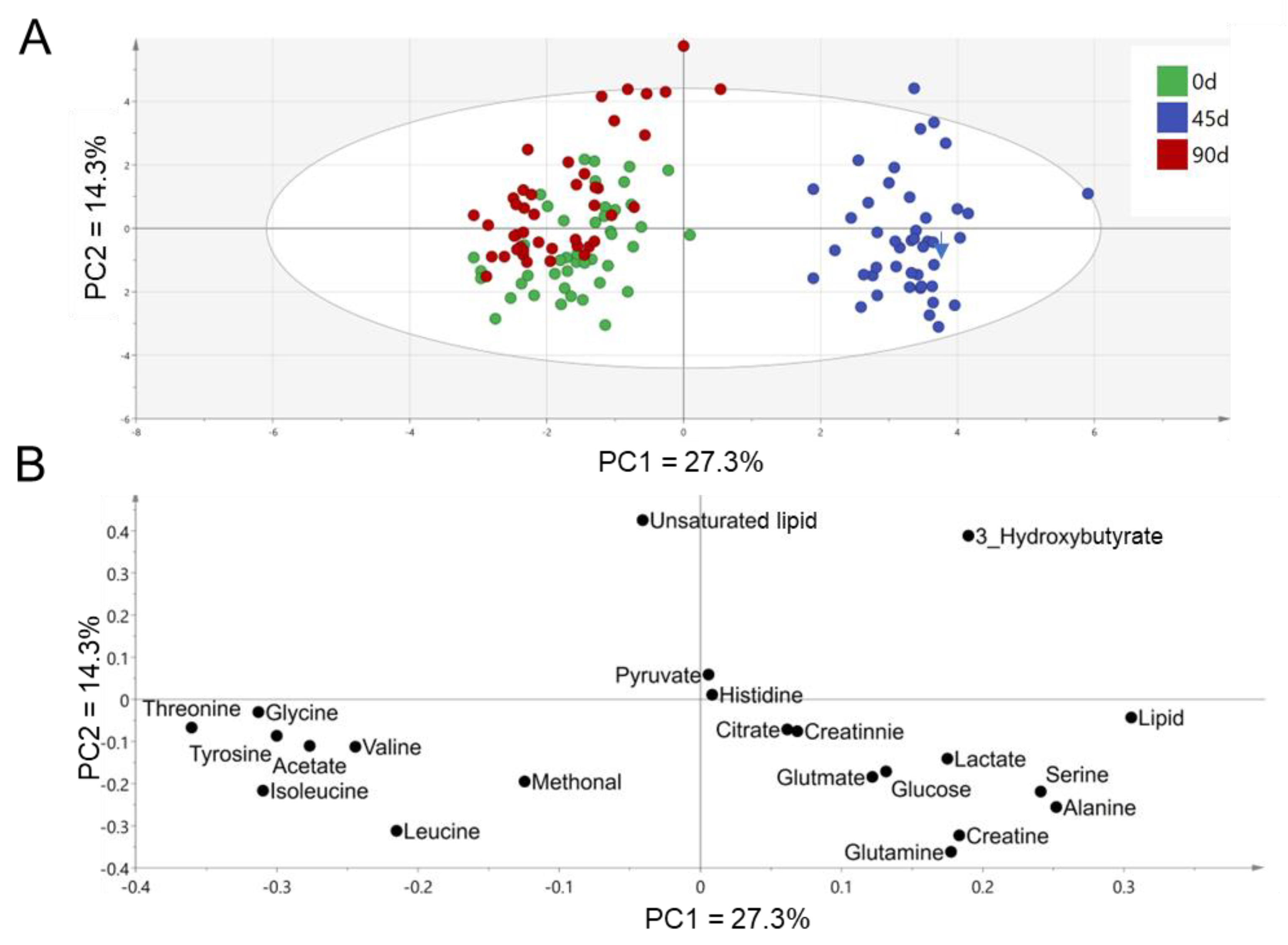
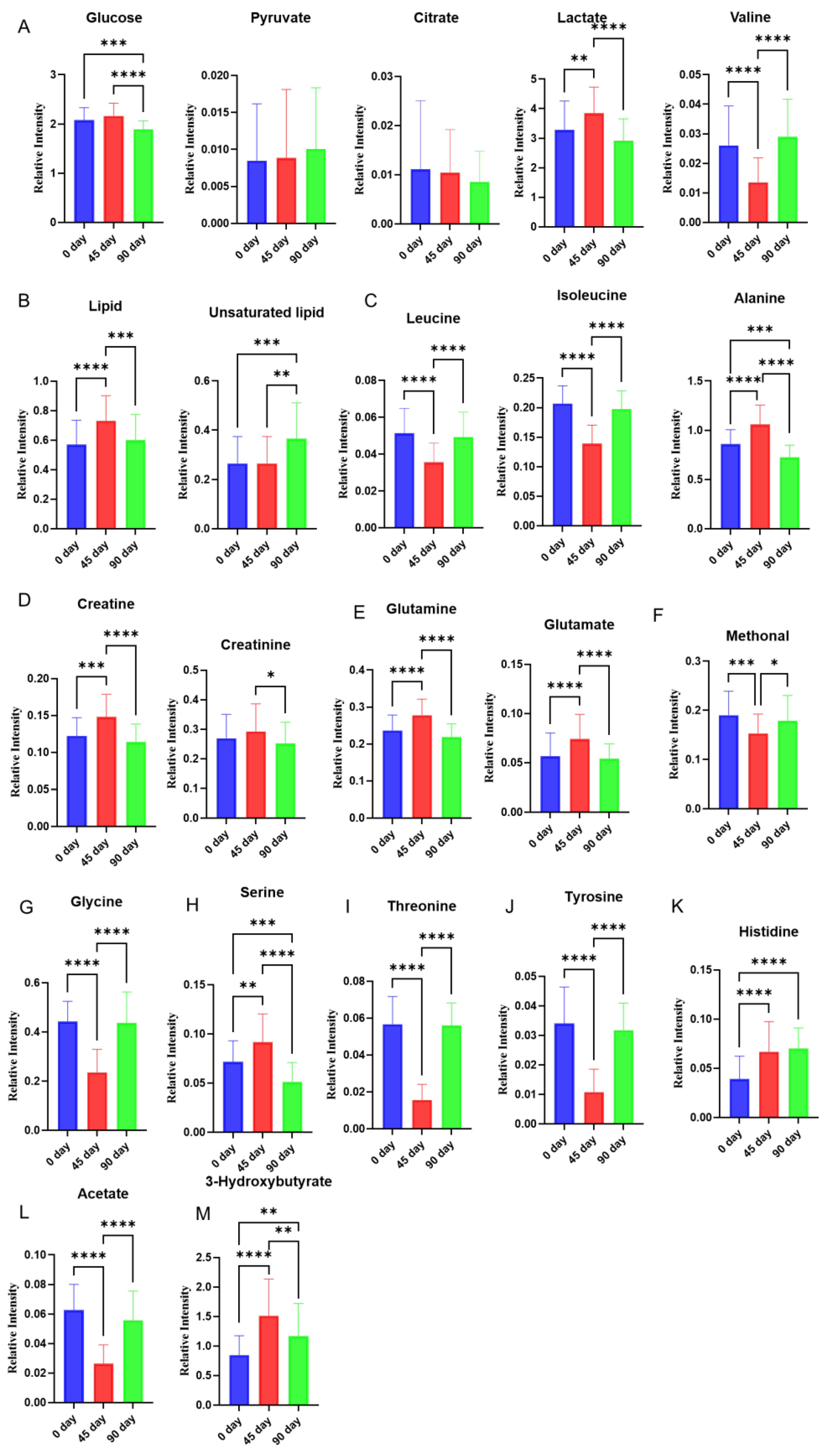
3.2. Effects of Sea Buckthorn Berries Puree on the Plasma Metabolites
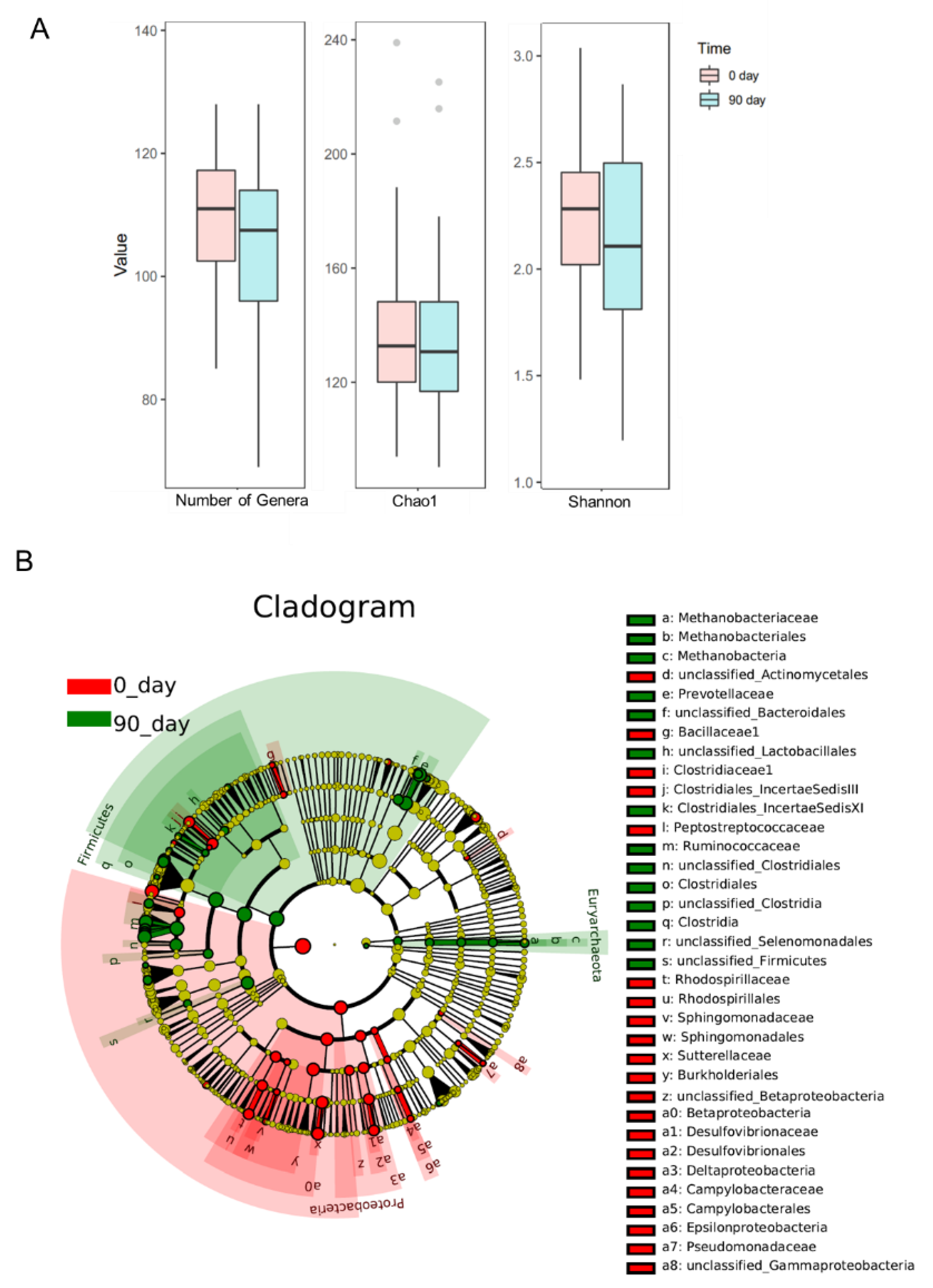
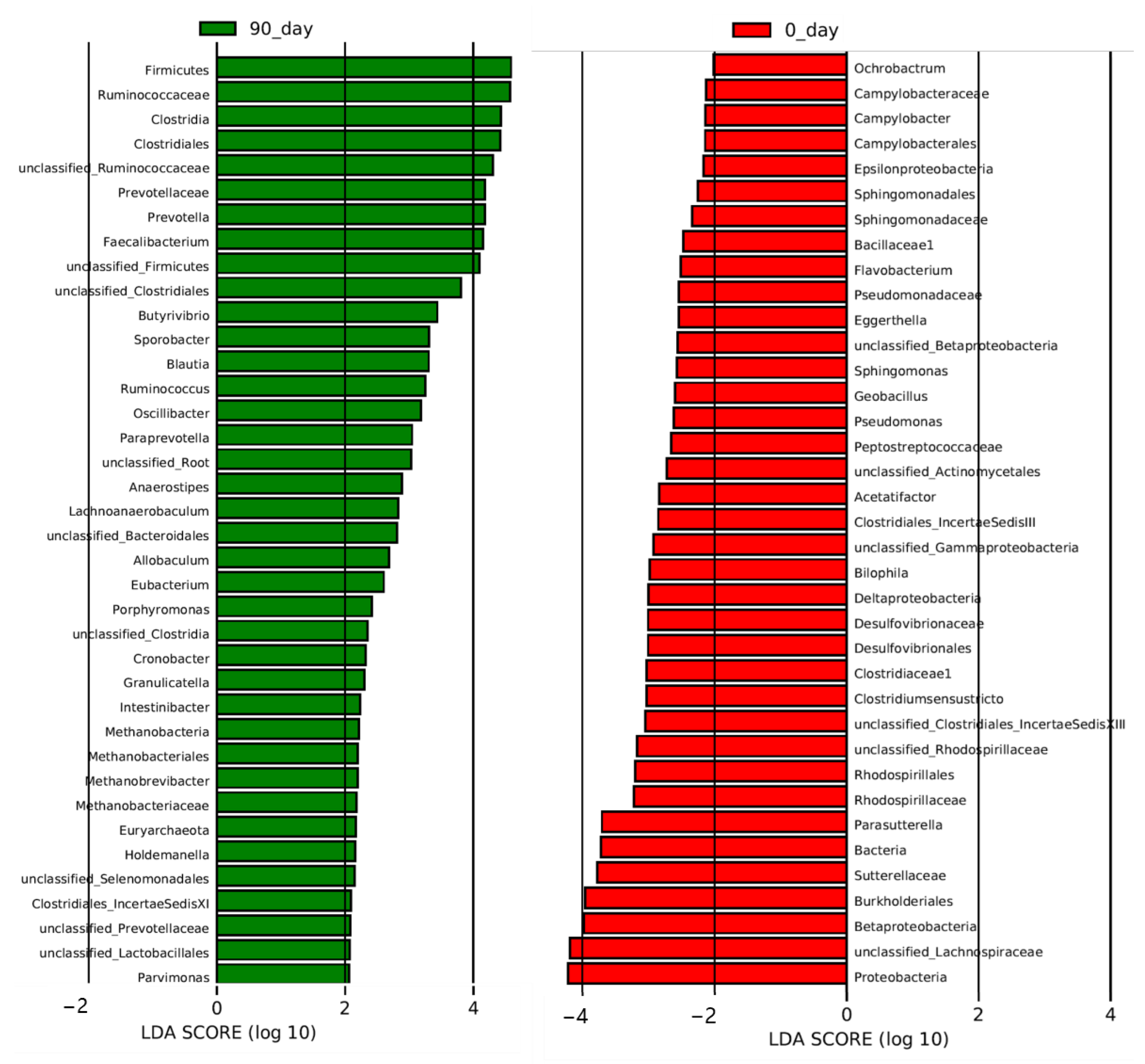
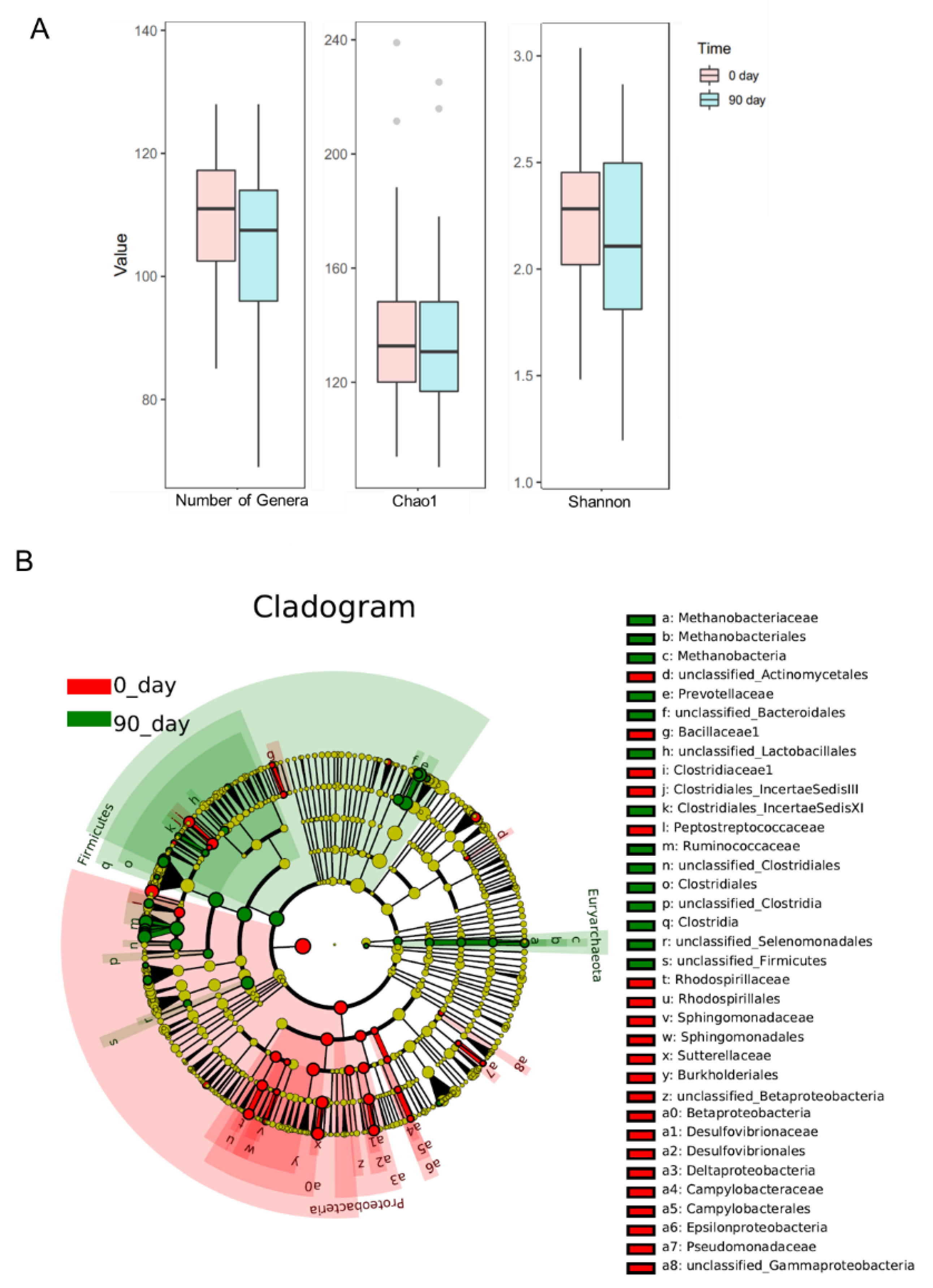
3.3. Effects of Sea Buckthorn Puree on Gut Microbiota
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steinberg, D. Atherogenesis in Perspective: Hypercholesterolemia and Inflammation as Partners in Crime. Nat. Med. 2002, 8, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Durrington, P. Dyslipidaemia. Lancet 2003, 362, 717–731. [Google Scholar] [CrossRef]
- Larmo, P.S.; Kangas, A.J.; Soininen, P.; Lehtonen, H.M.; Suomela, J.P.; Yang, B.; Viikari, J.; Ala-Korpela, M.; Kallio, H.P. Effects of Sea Buckthorn and Bilberry on Serum Metabolites Differ According to Baseline Metabolic Profiles in Overweight Women: A Randomized Crossover Trial. Am. J. Clin. Nutr. 2013, 98, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, H.M.; Suomela, J.P.; Tahvonen, R.; Vaarno, J.; Venojärvi, M.; Viikari, J.; Kallio, H. Berry Meals and Risk Factors Associated with Metabolic Syndrome. Eur. J. Clin. Nutr. 2010, 64, 614–621. [Google Scholar] [CrossRef]
- Chong, M.F.F.; MacDonald, R.; Lovegrove, J.A. Fruit Polyphenols and CVD Risk: A Review of Human Intervention Studies. Br. J. Nutr. 2010, 104 (Suppl. 3), S28–S39. [Google Scholar] [CrossRef]
- Xu, Y.J.; Kaur, M.; Dhillon, R.S.; Tappia, P.S.; Dhalla, N.S. Health Benefits of Sea Buckthorn for the Prevention of Cardiovascular Diseases. J. Funct. Foods 2011, 3, 2–12. [Google Scholar] [CrossRef]
- Yang, F.; Suo, Y.; Chen, D.; Tong, L. Protection against Vascular Endothelial Dysfunction by Polyphenols in Sea Buckthorn Berries in Rats with Hyperlipidemia. Biosci. Trends 2016, 10, 188–196. [Google Scholar] [CrossRef]
- Xue, Y.; Miao, Q.; Zhao, A.; Zheng, Y.; Zhang, Y.; Wang, P.; Kallio, H.; Yang, B. Effects of Sea Buckthorn (Hippophaë Rhamnoides) Juice and L-Quebrachitol on Type 2 Diabetes Mellitus in Db/Db Mice. J. Funct. Foods 2015, 16, 223–233. [Google Scholar] [CrossRef]
- Guo, C.; Han, L.; Li, M.; Yu, L. Seabuckthorn (Hippophaë Rhamnoides) Freeze-Dried Powder Protects against High-Fat Diet-Induced Obesity, Lipid Metabolism Disorders by Modulating the Gut Microbiota of Mice. Nutrients 2020, 12, 265. [Google Scholar] [CrossRef]
- Sayegh, M.; Miglio, C.; Ray, S. Potential Cardiovascular Implications of Sea Buckthorn Berry Consumption in Humans. Int. J. Food Sci. Nutr. 2014, 65, 521–528. [Google Scholar] [CrossRef]
- Lee, S.G.; Kim, B.; Yang, Y.; Pham, T.X.; Park, Y.-K.; Manatou, J.; Koo, S.I.; Chun, O.K.; Lee, J.-Y. Berry Anthocyanins Suppress the Expression and Secretion of Proinflammatory Mediators in Macrophages by Inhibiting Nuclear Translocation of NF-ΚB Independent of NRF2-Mediated Mechanism. J. Nutr. Biochem. 2014, 25, 404–411. [Google Scholar] [CrossRef]
- Lehtonen, H.M.; Suomela, J.P.; Tahvonen, R.; Yang, B.; Venojärvi, M.; Viikari, J.; Kallio, H. Different Berries and Berry Fractions Have Various but Slightly Positive Effects on the Associated Variables of Metabolic Diseases on Overweight and Obese Women. Eur. J. Clin. Nutr. 2011, 65, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Granado-Serrano, A.B.; Martín-Garí, M.; Sánchez, V.; Riart Solans, M.; Berdún, R.; Ludwig, I.A.; Rubió, L.; Vilaprinyó, E.; Portero-Otín, M.; Serrano, J.C.E. Faecal Bacterial and Short-Chain Fatty Acids Signature in Hypercholesterolemia. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between Phenolics and Gut Microbiota: Role in Human Health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef] [PubMed]
- Attri, S.; Sharma, K.; Raigond, P.; Goel, G. Colonic Fermentation of Polyphenolics from Sea Buckthorn (Hippophae rhamnoides) Berries: Assessment of Effects on Microbial Diversity by Principal Component Analysis. Food Res. Int. 2018, 105, 324–332. [Google Scholar] [CrossRef]
- Yuan, H.; Shi, F.; Meng, L.; Wang, W. Effect of Sea Buckthorn Protein on the Intestinal Microbial Community in Streptozotocin-Induced Diabetic Mice. Int. J. Biol. Macromol. 2018, 107, 1168–1174. [Google Scholar] [CrossRef]
- Nissen, S.E.; Tsunoda, T.; Tuzcu, E.M.; Schoenhagen, P.; Cooper, C.J.; Yasin, M.; Eaton, G.M.; Lauer, M.A.; Sheldon, W.S.; Grines, C.L.; et al. Effect of Recombinant ApoA-I Milano on Coronary Atherosclerosis in Patients with Acute Coronary Syndromes: A Randomized Controlled Trial. JAMA 2003, 290, 2292–2300. [Google Scholar] [CrossRef]
- Chen, K.; Wei, X.; Zhang, J.; Pariyani, R.; Jokioja, J.; Kortesniemi, M.; Linderborg, K.M.; Heinonen, J.; Sainio, T.; Zhang, Y.; et al. Effects of Anthocyanin Extracts from Bilberry (Vaccinium myrtillus L.) and Purple Potato (Solanum tuberosum L. Var. ’Synkeä Sakari’) on the Plasma Metabolomic Profile of Zucker Diabetic Fatty Rats. J. Agric. Food Chem. 2020, 68, 9436–9450. [Google Scholar] [CrossRef]
- Chen, K.; Wei, X.; Kortesniemi, M.; Pariyani, R.; Zhang, Y.; Yang, B. Effects of Acylated and Nonacylated Anthocyanins Extracts on Gut Metabolites and Microbiota in Diabetic Zucker Rats: A Metabolomic and Metagenomic Study. Food Res. Int. 2022, 153, 110978. [Google Scholar] [CrossRef]
- Janssens, Y.; Nielandt, J.; Bronselaer, A.; Debunne, N.; Verbeke, F.; Wynendaele, E.; Van Immerseel, F.; Vandewynckel, Y.P.; De Tré, G.; De Spiegeleer, B. Disbiome Database: Linking the Microbiome to Disease. BMC Microbiol. 2018, 18. [Google Scholar] [CrossRef]
- Tang, W.; Yao, X.; Xia, F.; Yang, M.; Chen, Z.; Zhou, B.; Liu, Q. Modulation of the Gut Microbiota in Rats by Hugan Qingzhi Tablets during the Treatment of High-Fat-Diet-Induced Nonalcoholic Fatty Liver Disease. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Jiang, Y.; Tan, A.; Ye, J.; Xian, X.; Xie, Y.; Wang, Q.; Yao, Z.; Mo, Z. 16S RRNA Gene Sequencing Reveals Altered Composition of Gut Microbiota in Individuals with Kidney Stones. Urolithiasis 2018, 46, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Zhang, A.H.; Miao, J.H.; Sun, H.; Yan, G.L.; Wu, F.F.; Wang, X.J. Gut Microbiota as Important Modulator of Metabolism in Health and Disease. RSC Adv. 2018, 8, 42380–42389. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The Gut Microbiome in Atherosclerotic Cardiovascular Disease. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Maya-Lucas, O.; Murugesan, S.; Nirmalkar, K.; Alcaraz, L.D.; Hoyo-Vadillo, C.; Pizano-Zárate, M.L.; García-Mena, J. The Gut Microbiome of Mexican Children Affected by Obesity. Anaerobe 2019, 55, 11–23. [Google Scholar] [CrossRef]
- Walldius, G.; Jungner, I. The ApoB/ApoA-I Ratio: A Strong, New Risk Factor for Cardiovascular Disease and a Target for Lipid-Lowering Therapy--a Review of the Evidence. J. Intern. Med. 2006, 259, 493–519. [Google Scholar] [CrossRef]
- Barter, P.J.; Rye, K.A. The Rationale for Using ApoA-I as a Clinical Marker of Cardiovascular Risk. J. Intern. Med. 2006, 259, 447–454. [Google Scholar] [CrossRef]
- Kurutas, E.B. The Importance of Antioxidants Which Play the Role in Cellular Response against Oxidative/Nitrosative Stress: Current State. Nutr. J. 2016, 15, 1–22. [Google Scholar] [CrossRef]
- Shulman, G.I. Unraveling the Cellular Mechanism of Insulin Resistance in Humans: New Insights from Magnetic Resonance Spectroscopy. Physiology 2004, 19, 183–190. [Google Scholar] [CrossRef]
- Wang, D.; Sun, Y.; Liu, H.; Zhang, X.; Ding, X.; Liu, S.; Han, B.; Wang, H.; Duan, X.; Sun, T. Low Lactic Acid and Hypercholesterolemia Reduce 90-Day Mortality in Patients Suffering From Septic Shock According to the Sepsis-3 Definition. 2021. Available online: https://www.researchsquare.com/article/rs-380037/v1 (accessed on 16 August 2022).
- Crawford, S.O.; Hoogeveen, R.C.; Brancati, F.L.; Astor, B.C.; Ballantyne, C.M.; Schmidt, M.I.; Young, J.H. Association of Blood Lactate with Type 2 Diabetes: The Atherosclerosis Risk in Communities Carotid MRI Study. Int. J. Epidemiol. 2010, 39, 1647–1655. [Google Scholar] [CrossRef]
- Hegazy, A.M.; Azeem, A.A.; Shahy, E.M.; El-Sayed, E.M. Comparative Study of Cholinergic and Oxidative Stress Biomarkers in Brains of Diabetic and Hypercholesterolemic Rats. Hum. Exp. Toxicol. 2016, 35, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Denis McGarry, J. Dysregulation of Fatty Acid Metabolism in the Etiology of Type 2 Diabetes. Diabetes 2002, 51, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Dolinger, M.; Ritaccio, G.; Mazurkiewicz, J.; Conti, D.; Zhu, X.; Huang, Y. Leucine Stimulates Insulin Secretion via Down-Regulation of Surface Expression of Adrenergic A2A Receptor through the MTOR (Mammalian Target of Rapamycin) Pathway: IMPLICATION IN NEW-ONSET DIABETES IN RENAL TRANSPLANTATION*. J. Biol. Chem. 2012, 287, 24795. [Google Scholar] [CrossRef] [PubMed]
- Doi, M.; Yamaoka, I.; Nakayama, M.; Mochizuki, S.; Sugahara, K.; Yoshizawa, F. Isoleucine, a Blood Glucose-Lowering Amino Acid, Increases Glucose Uptake in Rat Skeletal Muscle in the Absence of Increases in AMP-Activated Protein Kinase Activity. J. Nutr. 2005, 135, 2103–2108. [Google Scholar] [CrossRef] [PubMed]
- Schaeffner, E.S.; Kurth, T.; Curhan, G.C.; Glynn, R.J.; Rexrode, K.M.; Baigent, C.; Buring, J.E.; Gaziano, J.M. Cholesterol and the Risk of Renal Dysfunction in Apparently Healthy Men. J. Am. Soc. Nephrol. 2003, 14, 2084–2091. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and Creatinine Metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Earnest, C.P.; Almada, A.L.; Mitchell, T.L. High-Performance Capillary Electrophoresis-Pure Creatine Monohydrate Reduces Blood Lipids in Men and Women. Clin. Sci. 1996, 91, 113–118. [Google Scholar] [CrossRef]
- Li, B.; Lu, X.; Wang, J.; He, X.; Gu, Q.; Wang, L.; Yang, Y. The Metabonomics Study of P-Selectin Glycoprotein Ligand-1 (PSGL-1) Deficiency Inhibiting the Progression of Atherosclerosis in LDLR-/- Mice. Int. J. Biol. Sci. 2018, 14, 36. [Google Scholar] [CrossRef]
- Fechner, A.; Kiehntopf, M.; Jahreis, G. The Formation of Short-Chain Fatty Acids Is Positively Associated with the Blood Lipid-Lowering Effect of Lupin Kernel Fiber in Moderately Hypercholesterolemic Adults. J. Nutr. 2014, 144, 599–607. [Google Scholar] [CrossRef]
- Hu, J.; Kyrou, I.; Tan, B.K.; Dimitriadis, G.K.; Ramanjaneya, M.; Tripathi, G.; Patel, V.; James, S.; Kawan, M.; Chen, J.; et al. Short-Chain Fatty Acid Acetate Stimulates Adipogenesis and Mitochondrial Biogenesis via GPR43 in Brown Adipocytes. Endocrinology 2016, 157, 1881–1894. [Google Scholar] [CrossRef]
- Amiya, E. Interaction of Hyperlipidemia and Reactive Oxygen Species: Insights from the Lipid-Raft Platform. World J. Cardiol. 2016, 8, 689. [Google Scholar] [CrossRef] [PubMed]
- Patel, C.; Ghanim, H.; Ravishankar, S.; Sia, C.L.; Viswanathan, P.; Mohanty, P.; Dandona, P. Prolonged Reactive Oxygen Species Generation and Nuclear Factor-ΚB Activation after a High-Fat, High-Carbohydrate Meal in the Obese. J. Clin. Endocrinol. Metab. 2007, 92, 4476–4479. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z. Dietary Polyphenols as Potential Nutraceuticals in Management of Diabetes: A Review. J. Diabetes Metab. Disord. 2013, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of Gut Microbiota with Dietary Polyphenols and Consequences to Human Health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Wang, P.Y.; Wang, X.; Wan, Y.L.; Liu, Y.C. Butyrate Enhances Intestinal Epithelial Barrier Function via Up-Regulation of Tight Junction Protein Claudin-1 Transcription. Dig. Dis. Sci. 2012, 57, 3126–3135. [Google Scholar] [CrossRef]
- Su, H.; Xie, L.; Xu, Y.; Ke, H.; Bao, T.; Li, Y.; Chen, W. Pelargonidin-3- O-Glucoside Derived from Wild Raspberry Exerts Antihyperglycemic Effect by Inducing Autophagy and Modulating Gut Microbiota. J. Agric. Food Chem. 2020, 68, 13025–13037. [Google Scholar] [CrossRef]
- Brahe, L.K.; Le Chatelier, E.; Prifti, E.; Pons, N.; Kennedy, S.; Hansen, T.; Pedersen, O.; Astrup, A.; Ehrlich, S.D.; Larsen, L.H. Specific Gut Microbiota Features and Metabolic Markers in Postmenopausal Women with Obesity. Nutr. Diabetes 2015, 5, e159. [Google Scholar] [CrossRef]
- Lamichhane, S.; Sen, P.; Alves, M.A.; Ribeiro, H.C.; Raunioniemi, P.; Hyötyläinen, T.; Orešič, M. Linking Gut Microbiome and Lipid Metabolism: Moving beyond Associations. Metabolites 2021, 11, 55. [Google Scholar] [CrossRef]
- Henneke, L.; Schlicht, K.; Andreani, N.A.; Hollstein, T.; Demetrowitsch, T.; Knappe, C.; Hartmann, K.; Jensen-Kroll, J.; Rohmann, N.; Pohlschneider, D.; et al. A Dietary Carbohydrate–Gut Parasutterella–Human Fatty Acid Biosynthesis Metabolic Axis in Obesity and Type 2 Diabetes. Gut Microbes 2022, 14, 2057778. [Google Scholar] [CrossRef]
- Yamada, T.; Takahashi, D.; Hase, K. The Diet-Microbiota-Metabolite Axis Regulates the Host Physiology. J. Biochem. 2016, 160, 1–10. [Google Scholar] [CrossRef]
- Wang, X.; Xia, J.; Jiang, C. Role of Gut Microbiota in the Development of Non-Alcoholic Fatty Liver Disease. Liver Res. 2019, 3, 25–30. [Google Scholar] [CrossRef]
- Kübeck, R.; Bonet-Ripoll, C.; Hoffmann, C.; Walker, A.; Müller, V.M.; Schüppel, V.L.; Lagkouvardos, I.; Scholz, B.; Engel, K.H.; Daniel, H.; et al. Dietary Fat and Gut Microbiota Interactions Determine Diet-Induced Obesity in Mice. Mol. Metab. 2016, 5, 1162. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
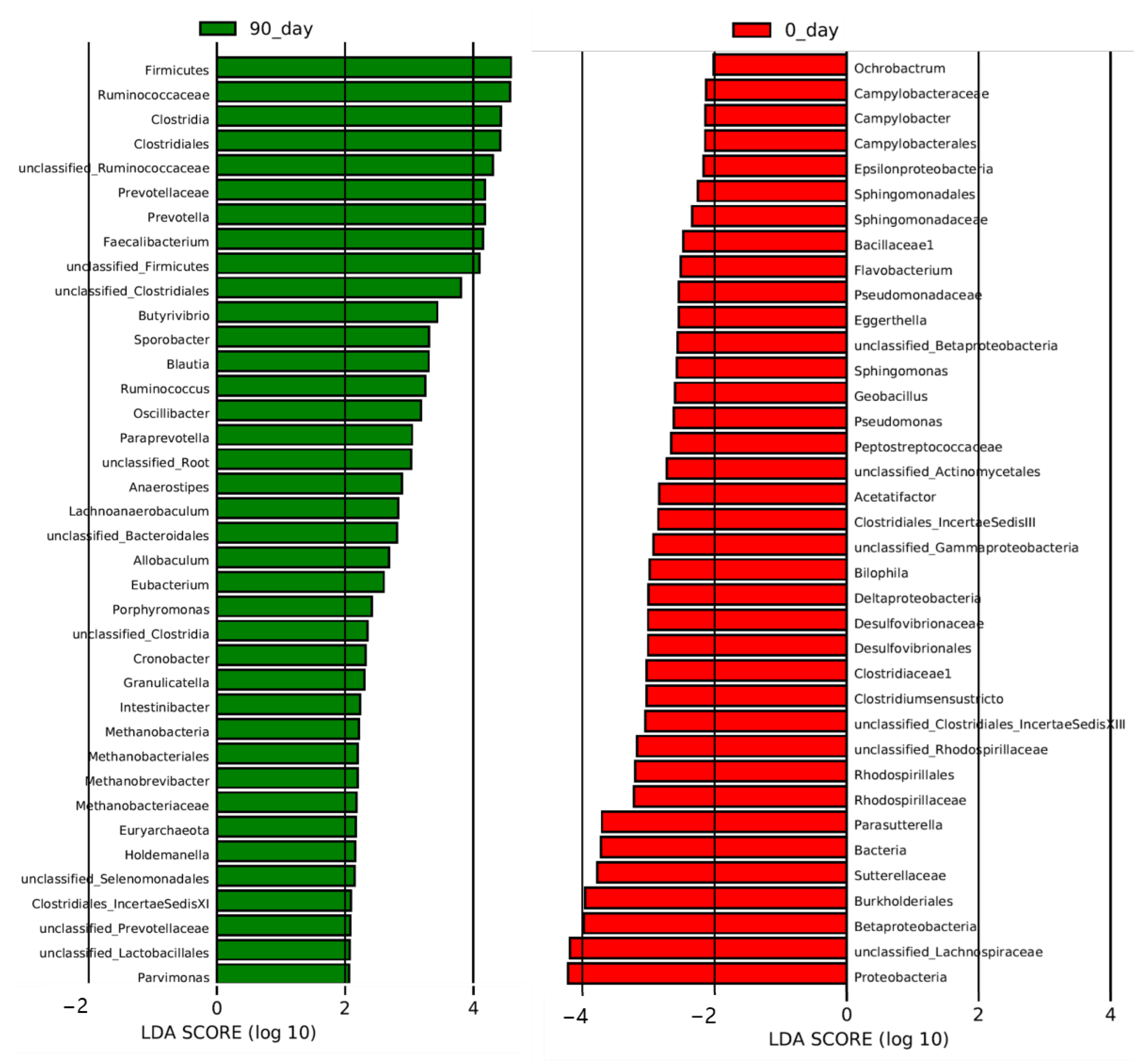
| Organisms | Characteristics | LDA Score | Regulation |
|---|---|---|---|
| Bacteria. Proteobacteria. Alphaproteobacteria. Rhizobialles. Brucellacese. Ochrobactrum | Oral pathogen | 2.01 | Downregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Clostridiaceae1. Clostridiumsensustricto | Butyrate producer | 3.03 | Downregulation |
| Bacteria. Proteobacteria. Beta proteobacteria. Burkholderiales. Sutterellaceae. Parasutterella | Potential pathogen, related to kidney stone and hypertension | 3.71 | Downregulation |
| Bacteria. Proteobacteria. Gammaproteobacteria Pseudomonadales. Pseudomonadaceae. Pseudomonas | Pathogen, related to many diseases | 2.62 | Downregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Lachnospiraceae. unclassified_Lachnospiraceae | Butyrate producer | 4.19 | Downregulation |
| Bacteria. Firmicutes. Clostridia Clostridiales. Clostridiales_IncertaeSedisXIll. Unclassified_Clostridiales_IncertaeSedis XI | Butyrate producer | 3.05 | Downregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Lachnospiraceae. Acetatifactor | Potential pathogen, related to obesity and lipid acid metabolism | 2.83 | Downregulation |
| Bacteria. Bacteroidetes. Flavobacteria. Flavobatteriales. Flavobacteriaceae. Flavobacterium | Pathogen | 2.51 | Downregulation |
| Bacteria. Proteobacteria. Alphaproteobacteria. Sphingomonadales. Sphingomonadaceae. Sphingomonas | Potential pathogen, related to many diseases | 2.57 | Downregulation |
| Bacteria. Actinobacteria. Actinobacteria. Coriobacteriales. Coriobacteriaceae. Eggerthella | Potential pathogen, related to coronary heart disease and chronic kidney disease | 2.54 | Downregulation |
| Bacteria. Proteobacteria. Delta proteobacteria. Desulfovibrionales. Desulfovibrionaceae. Bilophila | Potential pathogen, related to obesity | 2.99 | Downregulation |
| Bacteria. Firmicutes. Bacilli. Bacillales. Bacillaceas1. Geobacillus | Potential pathogen, related to psoriasis | 2.6 | Downregulation |
| Bacteria. Proteobacteria. Epsilonproteobacteria. Campylobacterales. Campylobacteraceae. Campylobacter | Pathogen | 2.13 | Downregulation |
| Bacteria. Proteobacteria. Alphaproteobacteria. Rhodospirilales. Rhodospirillaceae. unclassified_Rhodospirillaceae | Potential pathogen, related to diabetes and celiac disease | 3.17 | Downregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Lachnospiraceae. Anaerostipes | Butyrate producer | 2.88 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Lachnospiraceae. Lachnoanaerobaculum | Oral pathogen | 2.83 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Ruminococcaceae. Ruminococcus | Butyrate producer | 3.25 | Upregulation |
| Bacteria. Firmicutes. Erysipelotrichia. Erysipelotrichales. Erysipelotrichaceae. Holdemanella | Potential pathogen, related to metabolic diseases | 2.15 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Peptostreptococcaceae. Intestinibacter | Potential pathogen | 2.23 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Ruminococcaceae. Oscilllibacter | Butyrate producer | 3.18 | Upregulation |
| Bacteria. Bacteroidetes. Bacteroidia. Bacteroidales. Prevotellaceae. unclassified_Prevotellaceae | Related to high-fiber diet | 2.08 | Upregulation |
| Bacteria. Firmicutes. Bacilli. Lactoacillales. Carnobacteriaceae. Granulicatella | Potential pathogen | 2.3 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Lachnospiraceae. Butyrivibrio | Butyrate producer | 3.44 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Lachnospiraceae. Blautia | Butyrate producer | 3.3 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Eubacteriaceae. Eubacterium | Butyrate producer | 2.6 | Upregulation |
| Bacteria. Bacteroidetes. Bacteroidia. Bacteroidales. Porphyromonadaceae. Porphyromonas | Oral pathogen, also butyrate producer | 2.41 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Ruminococcaceae. Faecalibacterium | Butyrate producer | 4.15 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. ClostridialesIncertaeSedisXl. Parvimonas | Potential pathogen | 2.06 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Ruminococcaceae. unclassified_Ruminococcaceae | Butyrate producer | 4.3 | Upregulation |
| Bacteria. Firmicutes. Erysipelotrichia. Erysipelotrichales. Erysipelotrichaceae. Allobaculum | Potential pathogen, related to metabolic diseases | 2.69 | Upregulation |
| Bacteria. Bacteroidetes. Bacteroidia. Bacteroidales. Prevotellaceae. Prevotella | Related to high-fiber diet | 4.18 | Upregulation |
| Bacteria. Proteobacteria. Gammaproteobacteria. Enterobacteriales. Enterobacteriaceae. Cronobacter | Potential pathogen | 2.32 | Upregulation |
| Bacteria. Firmicutes. Clostridia. Clostridiales. Ruminococcaceae. Sporobacter | Butyrate producer | 3.31 | Upregulation |
| Archaea. Euryarchaeota. Methanobacteria. Methanobacteriales. Methanobacteriaceae. Methanobrevibacter | Efficient digestion of polysaccharides, producing methane | 2.19 | Upregulation |
| Bacteria. Bacteroidetes. Bacteroidia. Bacteroidales. Prevotellaceae. Paraprevotella | Potential pathogen | 3.04 | Upregulation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.; Zhou, F.; Zhang, J.; Li, P.; Zhang, Y.; Yang, B. Dietary Supplementation with Sea Buckthorn Berry Puree Alters Plasma Metabolomic Profile and Gut Microbiota Composition in Hypercholesterolemia Population. Foods 2022, 11, 2481. https://doi.org/10.3390/foods11162481
Chen K, Zhou F, Zhang J, Li P, Zhang Y, Yang B. Dietary Supplementation with Sea Buckthorn Berry Puree Alters Plasma Metabolomic Profile and Gut Microbiota Composition in Hypercholesterolemia Population. Foods. 2022; 11(16):2481. https://doi.org/10.3390/foods11162481
Chicago/Turabian StyleChen, Kang, Fangfei Zhou, Jian Zhang, Pin Li, Yumei Zhang, and Baoru Yang. 2022. "Dietary Supplementation with Sea Buckthorn Berry Puree Alters Plasma Metabolomic Profile and Gut Microbiota Composition in Hypercholesterolemia Population" Foods 11, no. 16: 2481. https://doi.org/10.3390/foods11162481
APA StyleChen, K., Zhou, F., Zhang, J., Li, P., Zhang, Y., & Yang, B. (2022). Dietary Supplementation with Sea Buckthorn Berry Puree Alters Plasma Metabolomic Profile and Gut Microbiota Composition in Hypercholesterolemia Population. Foods, 11(16), 2481. https://doi.org/10.3390/foods11162481