
Freeze-Dried Tuna Pepsin Powder Stabilized by Some Cryoprotectants: In Vitro Simulated Gastric Digestion toward Different Proteins and Its Storage Stability




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
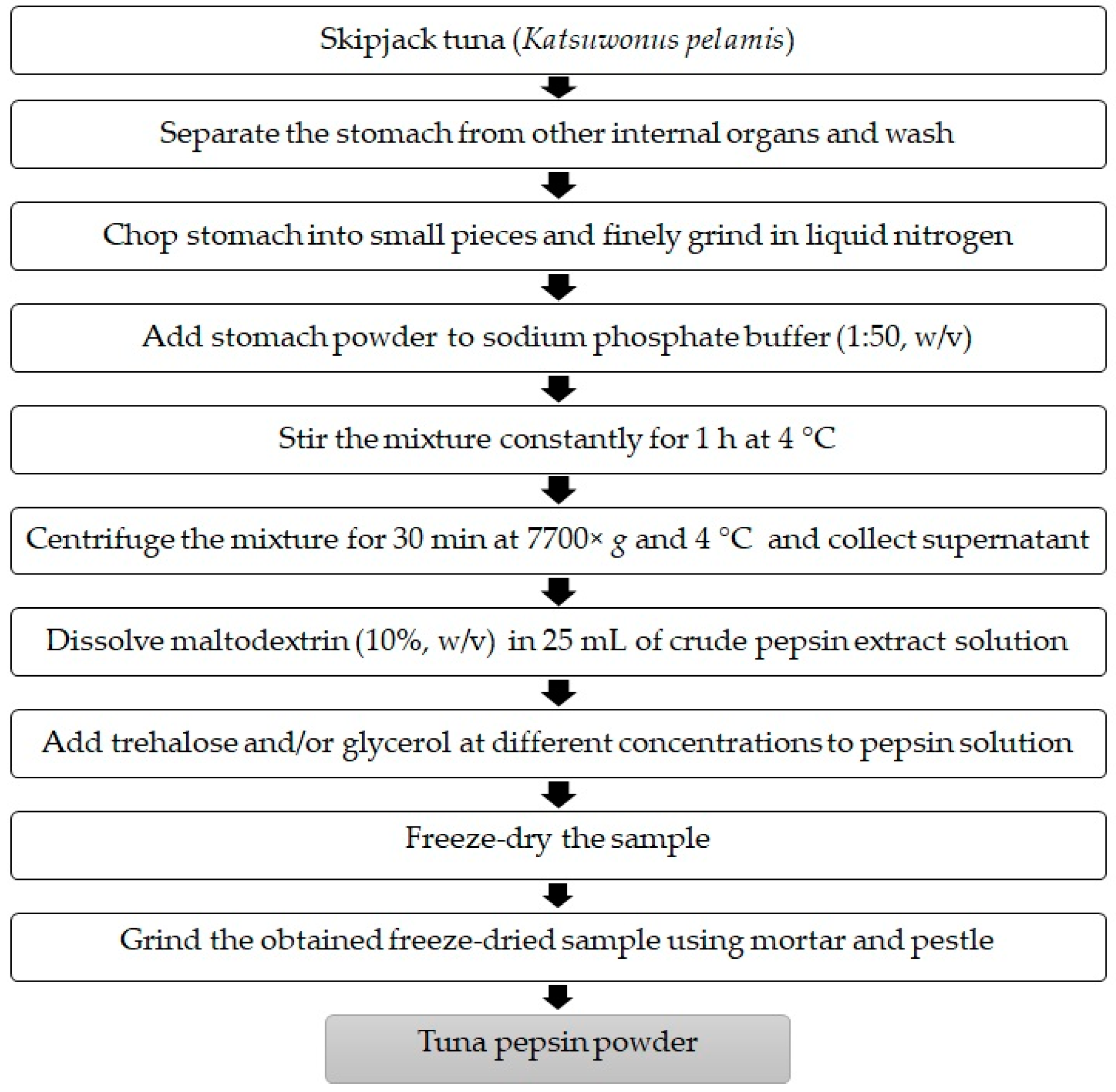
2.2. Preparation of Skipjack Tuna Stomach Powder
2.3. Preparation of Crude Pepsin Extract
2.4. Proteolytic Activity Assay
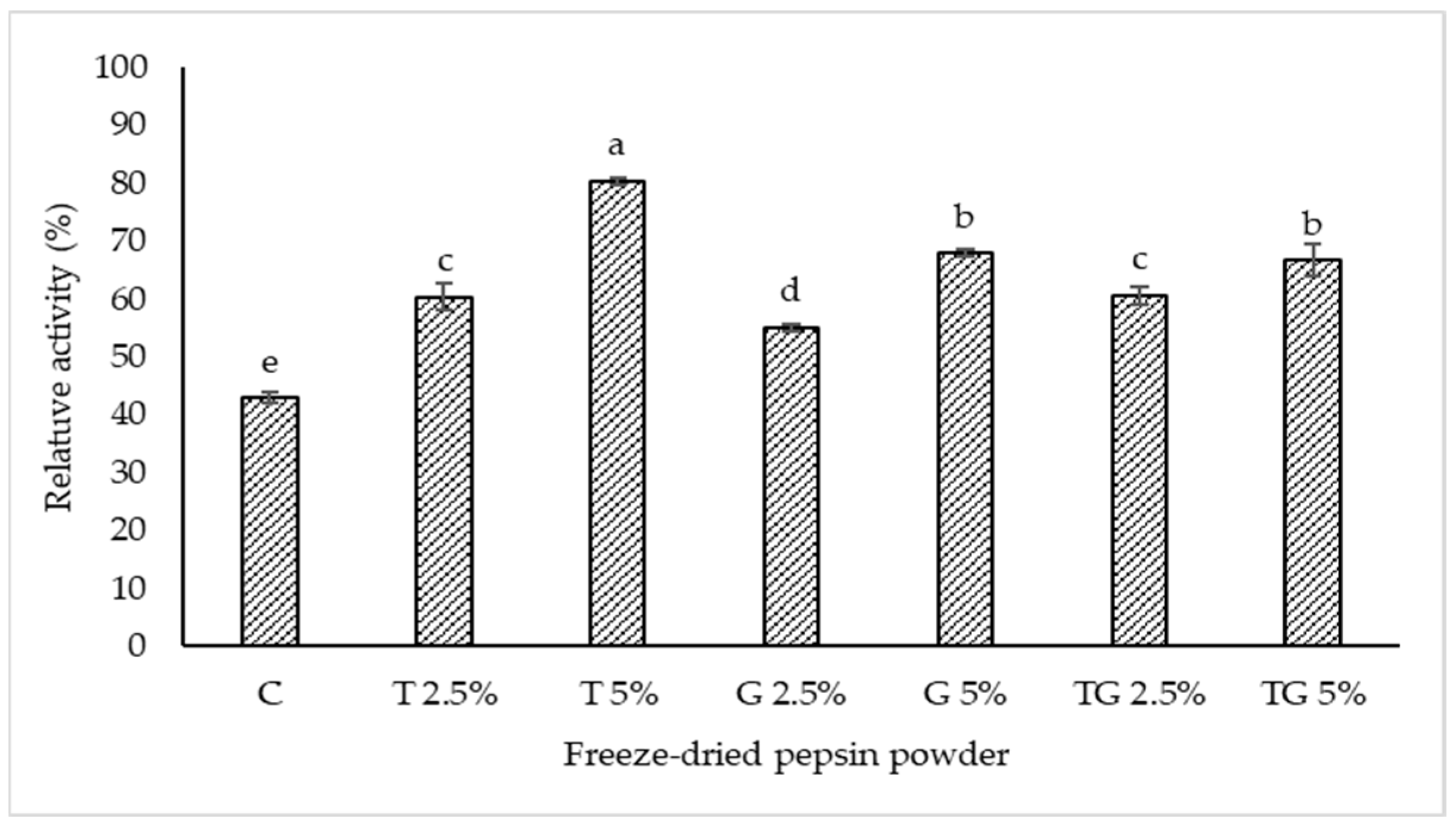
2.5. Effect of Cryoprotectants on Relative Pepsin Activity after Freeze-Drying
Relative Activity of TPP
2.6. Characterization of Selected TPP
2.6.1. Microstructure
2.6.2. Particle Size, Poly-Dispersity Index (PDI), and Zeta (ζ) Potential
2.6.3. Density and Flowability
2.6.4. Wettability and Solubility
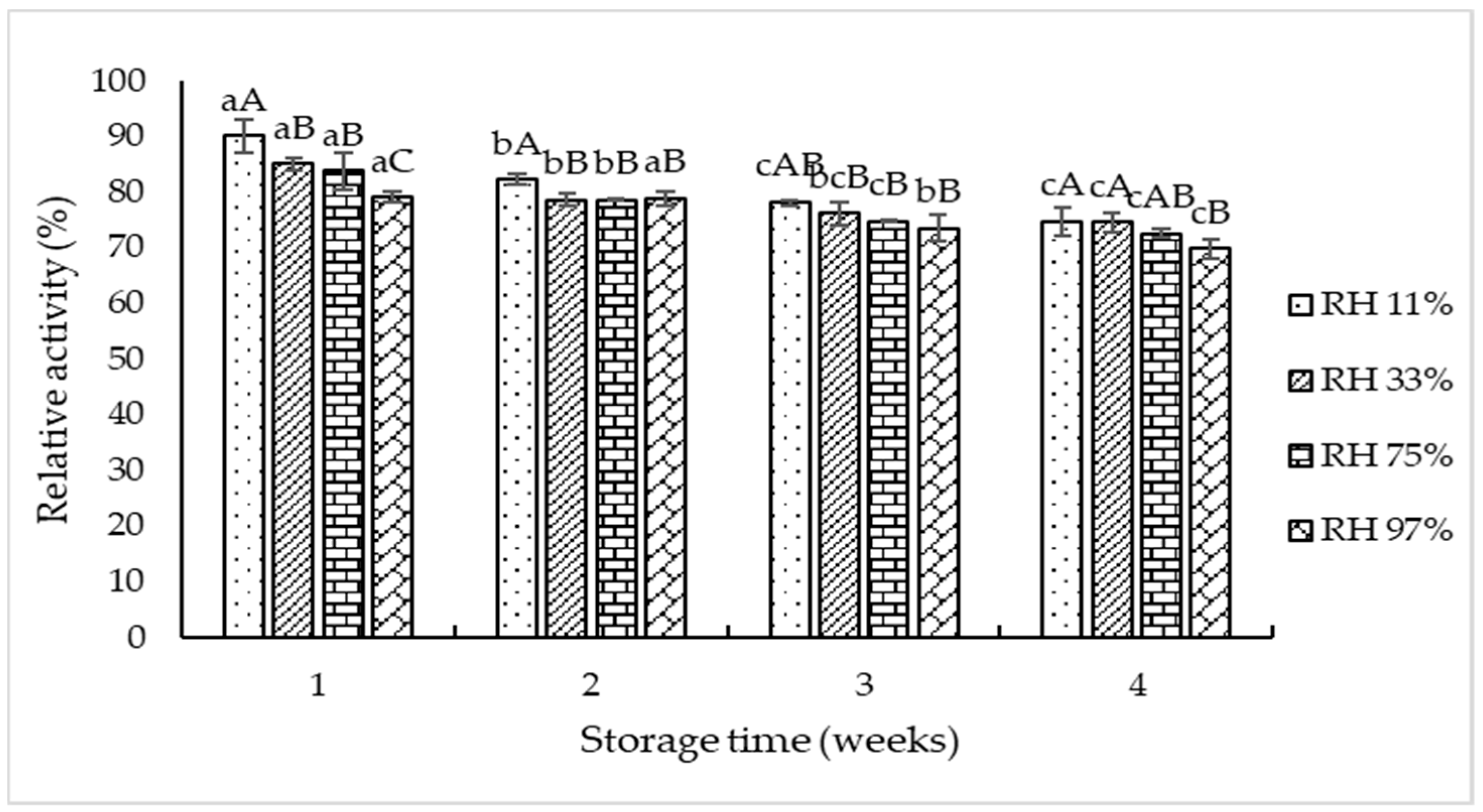
2.7. Storage Stability of the Selected TPP at Different Relative Humidities (RHs)
2.8. Characterization of Commercial and the Selected TPP
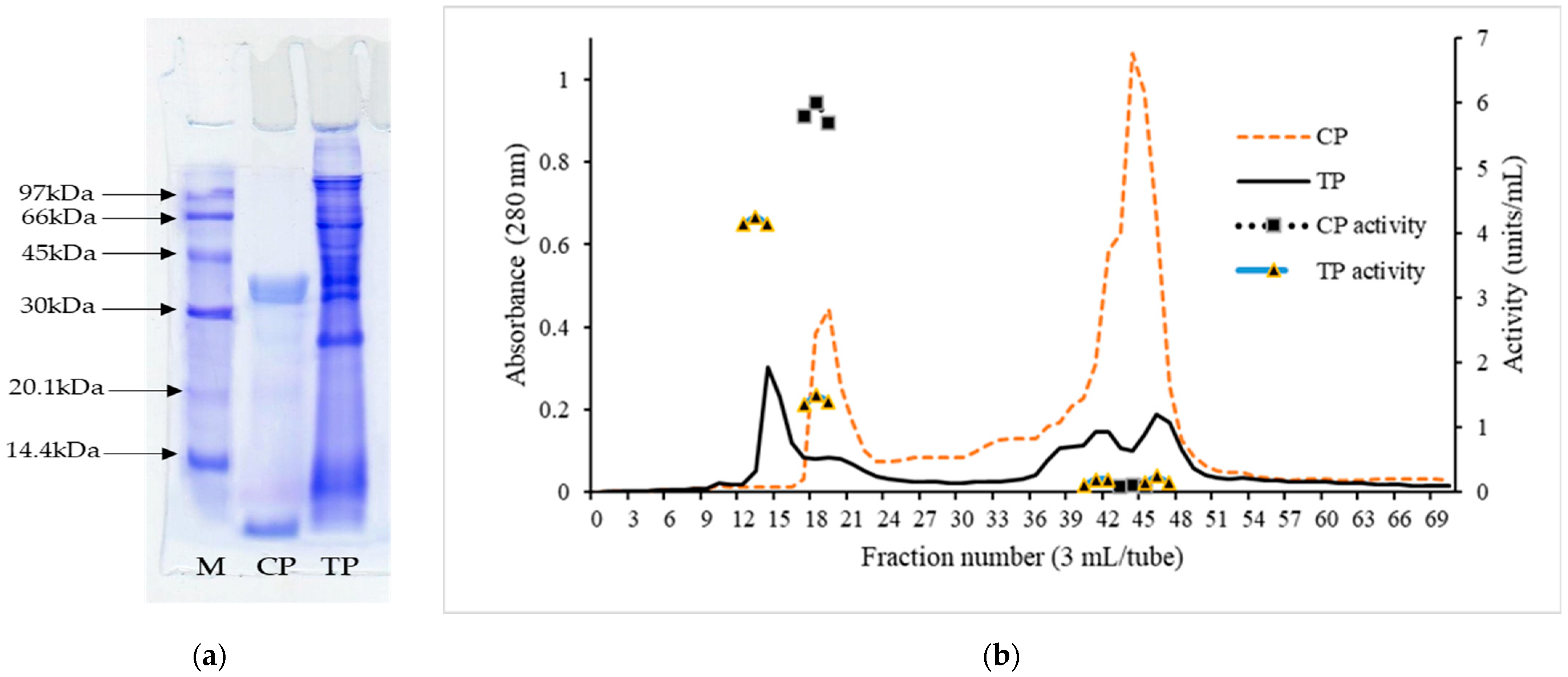
2.8.1. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.8.2. Molecular Weight Distribution
2.9. In Vitro Simulated Gastric Digestion of Different Proteins by the Selected TPP and CP
2.9.1. SDS-PAGE of Digests
2.9.2. Degree of Hydrolysis (DH) of Digests
2.9.3. Size Distribution of Digests
2.10. Statistical Analysis
3. Results and Discussion
3.1. Effect of Maltodextrin and Trehalose and/or Glycerol at Different Levels on Pepsin Activity after Freeze-Drying
3.2. Characteristics of Freeze-Dried Tuna Pepsin Powder Containing 10% Maltodextrin and 5% Trehalose (TPP-T5)
3.2.1. Microstructure
3.2.2. Particle Size, Poly-Dispersity Index (PDI), and Zeta (ζ) Potential
3.2.3. Density and Flowability
3.2.4. Wettability and Solubility
3.3. Storage Stability of TPP-T5 at Different Relative Humidities (RHs)
3.4. Characterization of Tuna Pepsin (TP) and Commercial Pepsin (CP)
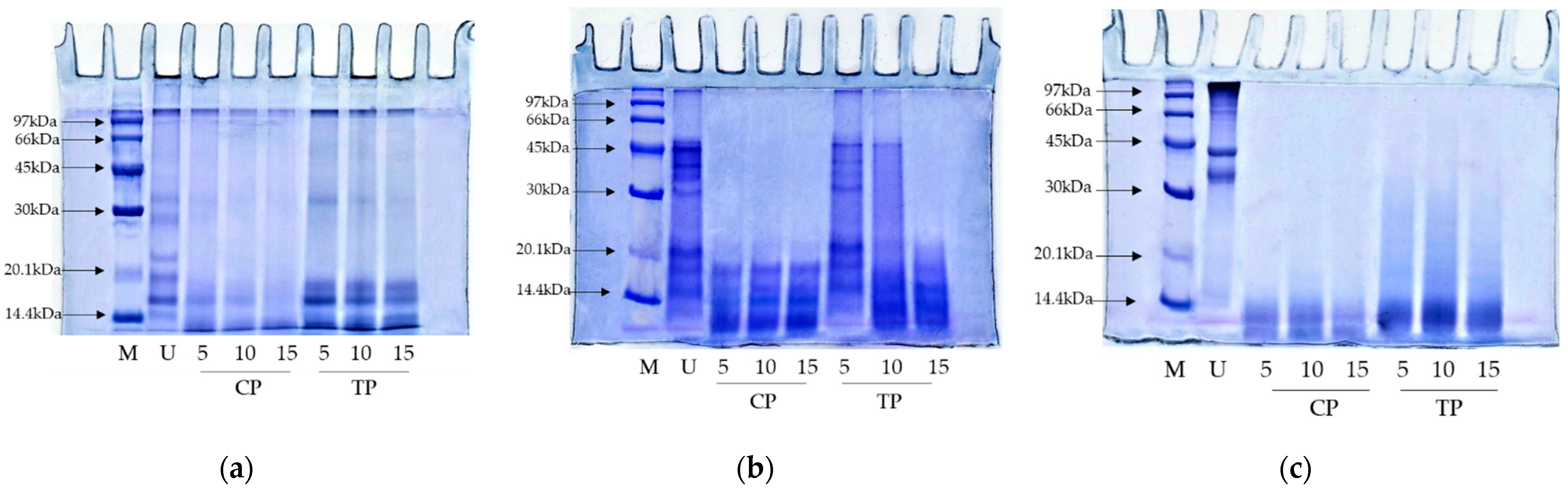
3.4.1. SDS-PAGE Protein Pattern
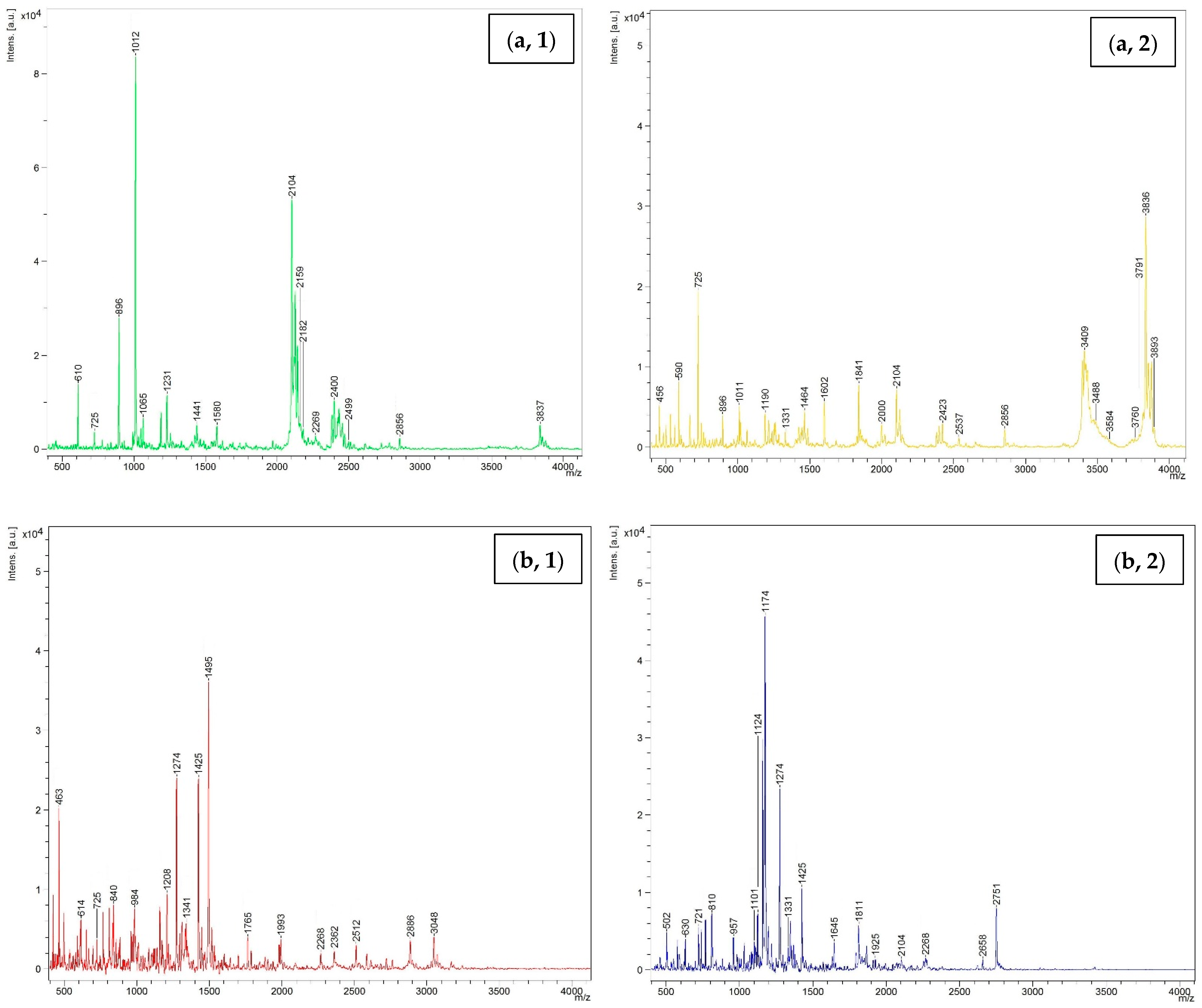
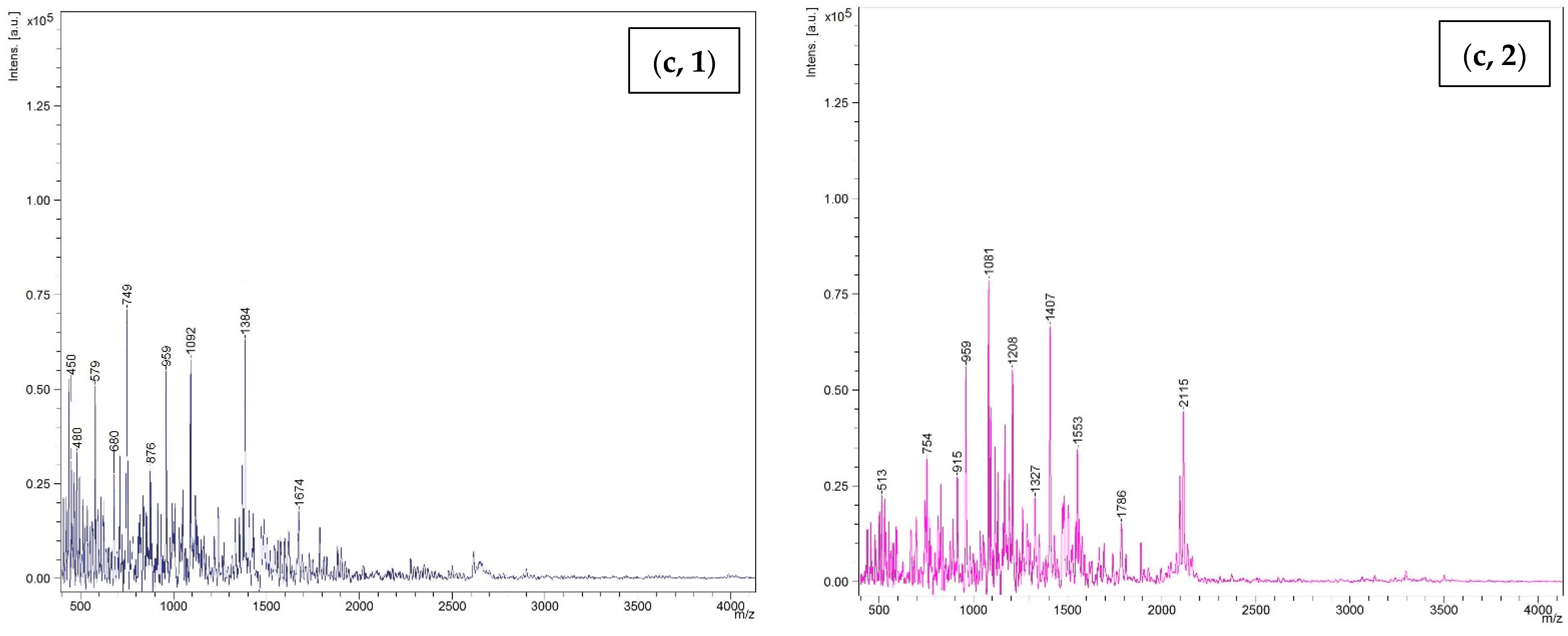
3.4.2. Molecular Weight Distribution
3.5. In Vitro Simulated Gastric Digestion of Different Proteins by TP and CP
3.5.1. Protein Patterns
3.5.2. Degree of Hydrolysis (DH)
3.5.3. Size Distribution
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heda, R.; Toro, F.; Tombazzi, C.R. Physiology, Pepsin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Zhao, L.; Budge, S.M.; Ghaly, A.E.; Brooks, M.S.; Dave, D. Extraction, purification and characterization of fish pepsin: A critical review. J. Food Process. Technol. 2011, 2, 1000126. [Google Scholar] [CrossRef] [Green Version]
- Moraes, G.; de Almeida, L.C. Nutrition and functional aspects of digestion in fish. In Biology and Physiology of Freshwater Neotropical Fish; Academic Press: Cambridge, MA, USA, 2020; pp. 251–271. [Google Scholar]
- Lee, S.Y.; Kang, J.H.; Kim, J.H.; Kim, H.W.; Oh, D.H.; Jeong, J.W.; Kim, B.K.; Hur, S.J. Effect of age-related in vitro human digestion with gut microbiota on antioxidative activity and stability of vitamins. LWT 2022, 159, 113243. [Google Scholar] [CrossRef]
- Buamard, N.; Aluko, R.E.; Benjakul, S. Stability of tuna trypsin-loaded alginate-chitosan beads in acidic stomach fluid and the release of active enzyme in a simulated intestinal tract environment. J. Food Biochem. 2020, 44, e13455. [Google Scholar] [CrossRef] [PubMed]
- Jurado, E.; Vicari, J.; Lechug, M.; Moya-Ramirez, I. Pepsin extraction process from swine wastes. Procedia Eng. 2012, 42, 1346–1350. [Google Scholar] [CrossRef] [Green Version]
- Imelio, N.; Marini, A.; Spelzini, D.; Picó, G.; Farruggia, B. Pepsin extraction from bovine stomach using aqueous two-phase systems: Molecular mechanism and influence of homogenate mass and phase volume ratio. J. Chromatogr. B 2008, 873, 133–138. [Google Scholar] [CrossRef]
- Nalinanon, S.; Benjakul, S.; Kishimura, H. Purification and biochemical properties of pepsins from the stomach of skipjack tuna (Katsuwonus pelamis). Eur. Food Res. Technol. 2010, 231, 259–269. [Google Scholar] [CrossRef]
- Klomklao, S.; Benjakul, S.; Visessanguan, W.; Kishimura, H.; Simpson, B.K. Purification and characterisation of trypsins from the spleen of skipjack tuna (Katsuwonus pelamis). Food Chem. 2007, 100, 1580–1589. [Google Scholar] [CrossRef]
- Tohmadlae, P.; Worawattanamateekul, W.; Hinsui, J. Characterization of acidic tuna protease and its application for extraction of tilapia collagen hydrolysate. Chiang Mai Univ. J. Nat. Sci. 2020, 19, 206–221. [Google Scholar] [CrossRef]
- Zarei, M.; Muhialdin, B.J.; Hassanzadeh, K.; Yea, C.S.; Ahmadi, R. Enzymatic Hydrolysis of Proteins. In Bioactive Peptides from Food: Sources, Analysis, and Functions; CRC Press: Boca Raton, FL, USA, 2022; pp. 189–208. [Google Scholar]
- Chen, Y.; Mutukuri, T.T.; Wilson, N.E.; Zhou, Q.T. Pharmaceutical protein solids: Drying technology, solid-state characterization and stability. Adv. Drug Deliv. Rev. 2021, 172, 211–233. [Google Scholar] [CrossRef]
- Corveleyn, S.; Remon, J.-P. Maltodextrins as lyoprotectants in the lyophilization of a model protein, LDH. Pharm. Res. 1996, 13, 146–150. [Google Scholar] [CrossRef]
- Sameti, M.; Bohr, G.; Kumar, M.R.; Kneuer, C.; Bakowsky, U.; Nacken, M.; Schmidt, H.; Lehr, C.-M. Stabilisation by freeze-drying of cationically modified silica nanoparticles for gene delivery. Int. J. Pharm. 2003, 266, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Gulzar, S.; Balange, A.K.; Nagarajarao, R.C.; Zhao, Q.; Benjakul, S. Microcapsules of shrimp oil using kidney bean protein isolate and κ-carrageenan as wall materials with the aid of ultrasonication or high-pressure microfluidization: Characteristics and oxidative stability. Foods 2022, 11, 1431. [Google Scholar] [CrossRef]
- Gulzar, S.; Benjakul, S. Nanoliposome powder containing shrimp oil increases free flowing behavior and storage stability. Eur. J. Lipid Sci. Technol. 2020, 122, 2000049. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Robinson, H.W.; Hogden, C.G. The biuret reaction in the determination of serum proteins. 1. A study of the conditions necessary for the production of a stable color which bears a quantitative relationship to the protein concentration. J. Biol. Chem. 1940, 135, 707–725. [Google Scholar] [CrossRef]
- Patil, U.; Benjakul, S. Comparative study on extraction of virgin coconut oil with the aid of partially purified protease from seabass pyloric caeca and commercial trypsin. J. Food Biochem. 2019, 43, e13024. [Google Scholar] [CrossRef] [PubMed]
- Benjakul, S.; Karnjanapratum, S.; Visessanguan, W. Hydrolysed collagen from Lates calcarifer skin: Its acute toxicity and impact on cell proliferation and collagen production of fibroblasts. Int. J. Food Sci. Technol. 2018, 53, 1871–1879. [Google Scholar] [CrossRef]
- Steel, R.; Torrie, J. Principles and Practices of Statistics; McGraw Book Coy Inc.: New York, NY, USA, 1980. [Google Scholar]
- Jain, N.K.; Roy, I. Effect of trehalose on protein structure. Protein Sci. 2009, 18, 24–36. [Google Scholar] [CrossRef]
- Santagapita, P.R.; Mazzobre, M.F.; del Pilar Buera, M. Invertase stability in alginate beads: Effect of trehalose and chitosan inclusion and of drying methods. Food Res. Int. 2012, 47, 321–330. [Google Scholar] [CrossRef]
- Nowak, D.; Jakubczyk, E. The freeze-drying of foods-The characteristic of the process course and the effect of its parameters on the physical properties of food materials. Foods 2020, 9, 1488. [Google Scholar] [CrossRef]
- Caparino, O.; Tang, J.; Nindo, C.; Sablani, S.; Powers, J.; Fellman, J. Effect of drying methods on the physical properties and microstructures of mango (Philippine ‘Carabao’var.) powder. J. Food Eng. 2012, 111, 135–148. [Google Scholar] [CrossRef]
- Ismail, E.A.; Aly, A.A.; Atallah, A.A. Quality and microstructure of freeze-dried yoghurt fortified with additives as protective agents. Heliyon 2020, 6, e05196. [Google Scholar] [CrossRef]
- Benjakul, S.; Mad-Ali, S.; Senphan, T.; Sookchoo, P. Biocalcium powder from precooked skipjack tuna bone: Production and its characteristics. J. Food Biochem. 2017, 41, e12412. [Google Scholar] [CrossRef]
- Bailey, J.E.; Cho, Y. Immobilization of glucoamylase and glucose oxidase in activated carbon: Effects of particle size and immobilization conditions on enzyme activity and effectiveness. Biotechnol. Bioeng. 1983, 25, 1923–1935. [Google Scholar] [CrossRef]
- Soleimanifard, M.; Feizy, J.; Maestrelli, F. Nanoencapsulation of propolis extract by sodium caseinate-maltodextrin complexes. Food Bioprod. Process. 2021, 128, 177–185. [Google Scholar] [CrossRef]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Hasanzadeh Davarani, F.; Javanmard, R.; Dokhani, A.; Khorasani, S.; Mozafari, M. Impact of particle size and polydispersity index on the clinical applications of lipidic nanocarrier systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agustinisari, I.; Mulia, K.; Nasikin, M. The effect of eugenol and chitosan concentration on the encapsulation of eugenol using whey protein–maltodextrin conjugates. Appl. Sci. 2020, 10, 3205. [Google Scholar] [CrossRef]
- Shnoudeh, A.J.; Hamad, I.; Abdo, R.W.; Qadumii, L.; Jaber, A.Y.; Surchi, H.S.; Alkelany, S.Z. Synthesis, characterization, and applications of metal nanoparticles. In Biomaterials and Bionanotechnology; Academic Press: Cambridge, MA, USA, 2019; pp. 527–612. [Google Scholar]
- Marcon, L.; Oliveras, J.; Puntes, V.F. In Situ nanoremediation of soils and groundwaters from the nanoparticle’s standpoint: A review. Sci. Total Environ. 2021, 791, 148324. [Google Scholar] [CrossRef] [PubMed]
- Albert, K.; Tóth, C.; Verasztó, B.; Vatai, G.; Koris, A. Microencapsulation analysis based on membrane technology: Basic research of spherical, solid precursor microcapsule production. Period. Polytech. Chem. Eng. 2016, 60, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Fonte, P.; Soares, S.; Costa, A.; Andrade, J.C.; Seabra, V.; Reis, S.; Sarmento, B. Effect of cryoprotectants on the porosity and stability of insulin-loaded PLGA nanoparticles after freeze-drying. Biomatter 2012, 2, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Bae, E.; Lee, S. Microencapsulation of avocado oil by spray drying using whey protein and maltodextrin. J. Microencapsul. 2008, 25, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Teo, A.; Lam, Y.; Lee, S.J.; Goh, K.K. Spray drying of whey protein stabilized nanoemulsions containing different wall materials–maltodextrin or trehalose. LWT 2021, 136, 110344. [Google Scholar] [CrossRef]
- Seth, D.; Mishra, H.N.; Deka, S.C. Functional and reconstitution properties of spray-dried sweetened yogurt powder as influenced by processing conditions. Int. J. Food Prop. 2017, 20, 1603–1611. [Google Scholar] [CrossRef]
- Kawai, K.; Suzuki, T. Stabilizing effect of four types of disaccharide on the enzymatic activity of freeze-dried lactate dehydrogenase: Step by step evaluation from freezing to storage. Pharm. Res. 2007, 24, 1883–1890. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Pepsin, A. Handbook of Proteolytic Enzymes; Academic Press: Cambridge, MA, USA, 2004; pp. 19–28. [Google Scholar]
- Bougatef, A.; Balti, R.; Zaied, S.B.; Souissi, N.; Nasri, M. Pepsinogen and pepsin from the stomach of smooth hound (Mustelus mustelus): Purification, characterization and amino acid terminal sequences. Food Chem. 2008, 107, 777–784. [Google Scholar] [CrossRef]
- Kageyama, T. Pepsinogens, progastricsins, and prochymosins: Structure, function, evolution, and development. Cell Mol. Life Sci. 2002, 59, 288–306. [Google Scholar] [CrossRef]
- Madureira, A.R.; Pereira, C.I.; Gomes, A.M.; Pintado, M.E.; Malcata, F.X. Bovine whey proteins–Overview on their main biological properties. Food Res. Int. 2007, 40, 1197–1211. [Google Scholar] [CrossRef]
- Uniacke-Lowe, T.; Fox, P.F. Chapter 4—Chymosin, Pepsins and Other Aspartyl Proteinases: Structures, Functions, Catalytic Mechanism and Milk-Clotting Properties. In Cheese, 4th ed.; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: San Diego, CA, USA, 2017; pp. 69–113. [Google Scholar]
- Shahidi, F.; Kamil, Y.J. Enzymes from fish and aquatic invertebrates and their application in the food industry. Trends Food Sci Technol. 2001, 12, 435–464. [Google Scholar] [CrossRef]
- Yildirim-Elikoglu, S.; Vural, H.; Erdem, Y.K. Effect of phenolic compounds on the activity of proteolytic enzymes during rennet induced coagulation of milk and ripening of miniature cheese. LWT 2021, 136, 110337. [Google Scholar] [CrossRef]
- Kaewudom, P.; Benjakul, S.; Kijroongrojana, K. Effect of bovine and fish gelatin in combination with microbial transglutaminase on gel properties of threadfin bream surimi. Int. Aquat. Res. 2012, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; Zhou, G.; Song, S.; Xu, X.; Voglmeir, J.; Liu, L.; Zhao, F.; Li, M.; Li, L.; Yu, X. Discrimination of in vitro and in vivo digestion products of meat proteins from pork, beef, chicken, and fish. Proteomics 2015, 15, 3688–3698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibilla, S.; Godfrey, M.; Brewer, S.; Budh-Raja, A.; Genovese, L. An overview of the beneficial effects of hydrolysed collagen as a nutraceutical on skin properties: Scientific background and clinical studies. Open Nutraceuticals J. 2015, 8, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Guo, Z.; Wei, J.; Han, L.; Yu, Q.-l.; Chen, H.; Chen, Y.; Zhang, W. Extraction of low molecular weight peptides from bovine bone using ultrasound-assisted double enzyme hydrolysis: Impact on the antioxidant activities of the extracted peptides. LWT 2021, 146, 111470. [Google Scholar] [CrossRef]
- Gu, R.-Z.; Li, C.-Y.; Liu, W.-Y.; Yi, W.-X.; Cai, M.-Y. Angiotensin I-converting enzyme inhibitory activity of low-molecular-weight peptides from Atlantic salmon (Salmo salar L.) skin. Food Res. Int. 2011, 44, 1536–1540. [Google Scholar] [CrossRef]
- Lee, J.-M.; You, S.-G.; Kim, S.-M. Functional activities of low molecular weight peptides purified from enzymatic hydrolysates of seaweeds. J. Korean Soc. Food Sci. Nutr. 2005, 34, 1124–1129. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Physical Properties | Mean ± SD * |
|---|---|---|
| 1 | Particle size (µm) | 65.42 ± 57.60 |
| 2 | Poly dispersion index (PDI) | 0.474 ± 0.01 |
| 3 | Zeta potential (mV) | −19.95 ± 1.00 |
| 4 | Untapped density (kg m−3) | 0.52 ± 0.03 |
| 5 | Tapped density (kg m−3) | 0.79 ± 0.05 |
| 6 | Carr index | 34.76 ± 0.61 |
| 7 | Wettability (min) | 16.36 ± 0.73 |
| 8 | Solubility (%) | 93.58 ± 1.25 |
| Samples | Enzyme Level (units/g Protein) | Degree of Hydrolysis (%) | |
|---|---|---|---|
| CP * | TP ** | ||
| Whey protein isolate (WPI) | 5 | 59.69 ± 1.66 Ac | 32.54 ± 0.81 Bc |
| 10 | 74.17 ± 3.88 Ab | 40.54 ± 1.29 Bb | |
| 15 | 94.61 ± 0.85 Aa | 63.21 ± 0.64 Ba | |
| Kidney bean protein isolate (KBPI) | 5 | 33.71 ± 1.22 Ac | 11.97 ± 1.37 Bc |
| 10 | 50.13 ± 2.19 Ab | 29.10 ± 1.08 Bb | |
| 15 | 58.59 ± 0.89 Aa | 44.25 ± 1.30 Ba | |
| Threadfin bream fish mince (TBFM) | 5 | 69.93 ± 1.40 Ac | 65.65 ± 1.29 Bc |
| 10 | 80.94 ± 1.06 Ab | 72.26 ± 0.69 Bb | |
| 15 | 96.60 ± 1.51 Aa | 83.24 ± 0.97 Ba | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patil, U.; Nikoo, M.; Zhang, B.; Benjakul, S. Freeze-Dried Tuna Pepsin Powder Stabilized by Some Cryoprotectants: In Vitro Simulated Gastric Digestion toward Different Proteins and Its Storage Stability. Foods 2022, 11, 2292. https://doi.org/10.3390/foods11152292
Patil U, Nikoo M, Zhang B, Benjakul S. Freeze-Dried Tuna Pepsin Powder Stabilized by Some Cryoprotectants: In Vitro Simulated Gastric Digestion toward Different Proteins and Its Storage Stability. Foods. 2022; 11(15):2292. https://doi.org/10.3390/foods11152292
Chicago/Turabian StylePatil, Umesh, Mehdi Nikoo, Bin Zhang, and Soottawat Benjakul. 2022. "Freeze-Dried Tuna Pepsin Powder Stabilized by Some Cryoprotectants: In Vitro Simulated Gastric Digestion toward Different Proteins and Its Storage Stability" Foods 11, no. 15: 2292. https://doi.org/10.3390/foods11152292
APA StylePatil, U., Nikoo, M., Zhang, B., & Benjakul, S. (2022). Freeze-Dried Tuna Pepsin Powder Stabilized by Some Cryoprotectants: In Vitro Simulated Gastric Digestion toward Different Proteins and Its Storage Stability. Foods, 11(15), 2292. https://doi.org/10.3390/foods11152292