Fate of Salmonella enterica Typhimurium and Listeria monocytogenes in Black Soldier Fly (Hermetia illucens) Larvae Reared on Two Artificial Diets

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rearing Conditions of H. illucens
2.2. Pathogen Dynamics in BSF and Two Growing Substrates
2.2.1. Salmonella Typhimurium and Listeria monocytogenes Challenge Tests
2.2.2. Physicochemical Analyses
2.3. Statistical Analysis
3. Results
3.1. Pathogen Dynamics in BSF and the Growing Substrates
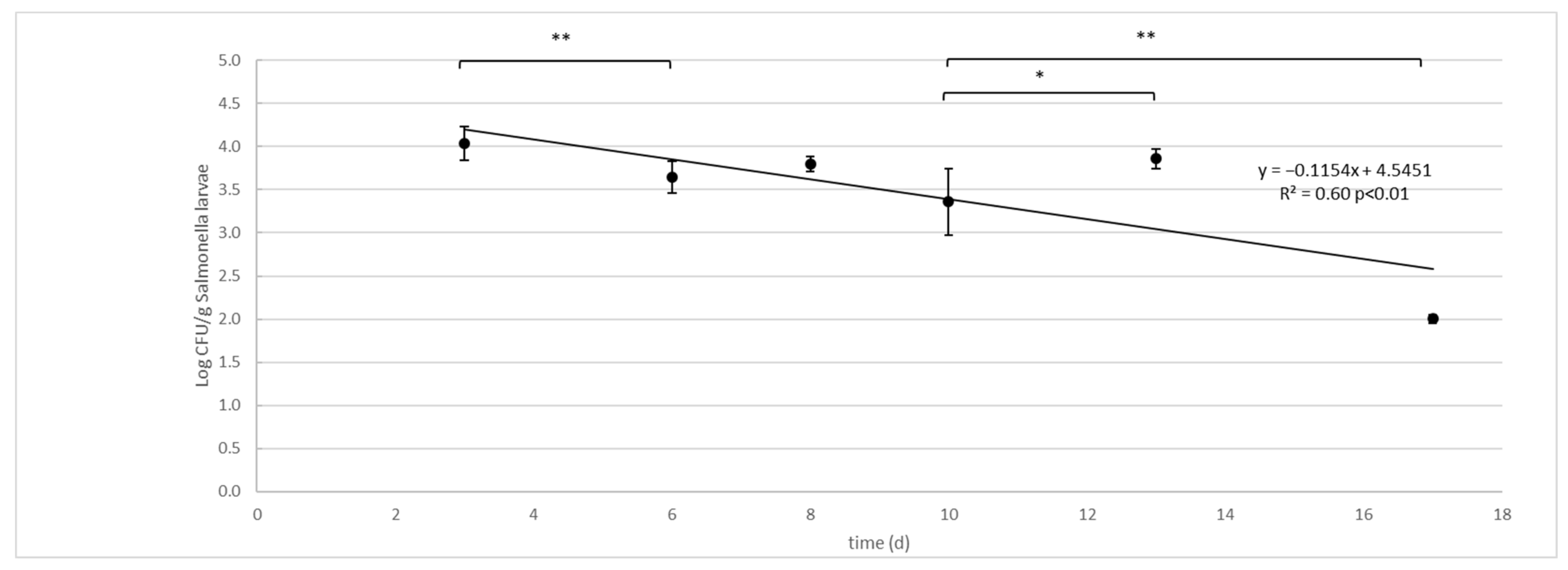
3.2. Salmonella enterica Typhimurium Challenge Test
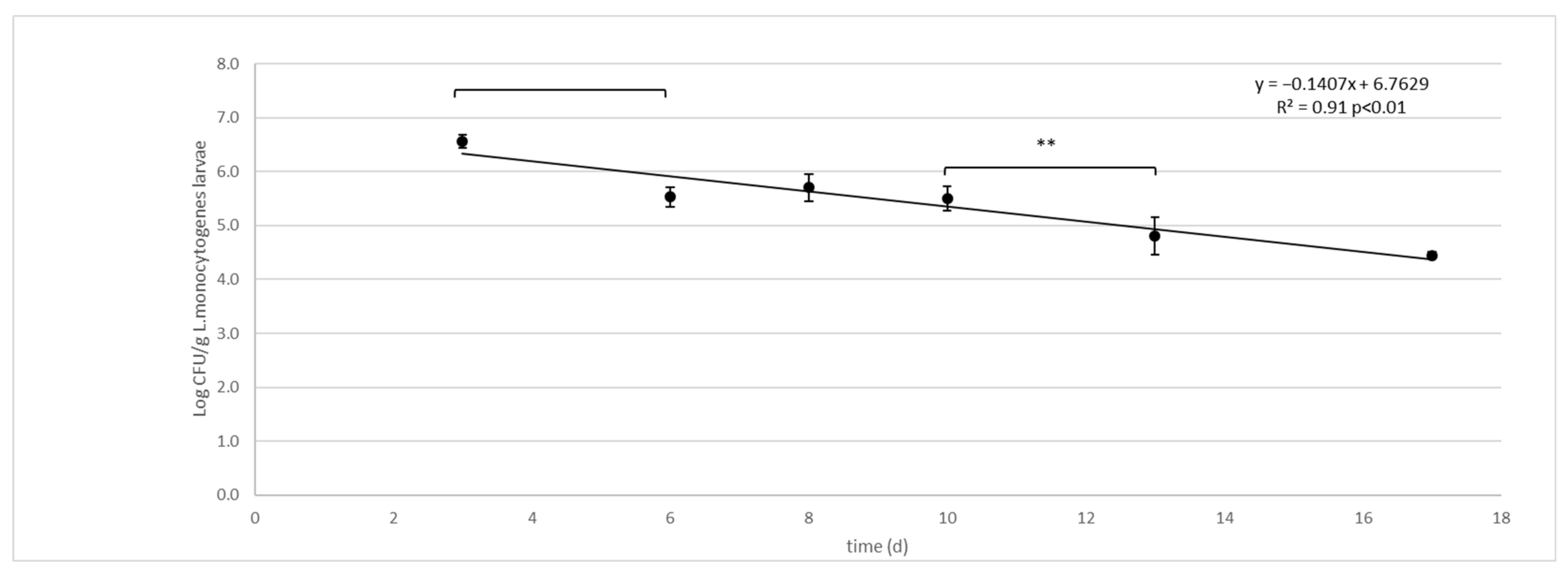
3.3. Listeria monocytogenes Challenge Test
3.4. Comparison of Salmonella enterica Typhimurium and L. monocytogenes Challenge Test
3.5. Physicochemical Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vandeweyer, D.; Crauwels, S.; Lievens, B.; Van Campenhout, L. Microbial counts of mealworm larvae (Tenebrio molitor) and crickets (Acheta domesticus and Gryllodes sigillatus) from different rearing companies and different production batches. Int. J. Food Microbiol. 2017, 242, 13–18. [Google Scholar] [CrossRef] [PubMed]
- European Union. Regulation (EU) 2283/2015 of the European Parliament and Council of the of 25 November 2015 on novel foods, amending regulation (EU) no 1169/2011 of the European Parliament and of the Council and repealing Regulation (EC) no 258/97 of the European Parliament and of the Council and Commission Regulation (EC) no 1852/2001 OJ 327/1. Off. J. Eur. Union 2015, 1–22. [Google Scholar]
- Smetana, S.; Schmitt, E.; Mathys, A. Sustainable use of Hermetia illucens insect biomass for feed and food: Attributional and consequential life cycle assessment. Resour. Conserv. Recycl. 2019, 144, 285–296. [Google Scholar] [CrossRef]
- Dickie, F.; Miyamoto, M.; Collins, M.C. The Potential of Insect Farming to Increase Food Security; IntechOpen: London, UK, 2019. [Google Scholar]
- Pinotti, L.; Giromini, C.; Ottoboni, M.; Tretola, M.; Marchis, D. Insects and former foodstuffs for upgrading food waste biomasses/streams to feed ingredients for farm animals. Animal 2019, 13, 1365–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.K.; Yong, H.I.; Kim, Y.B.; Kim, H.W.; Choi, Y.S. Edible Insects as a Protein Source: A Review of Public Perception, Processing Technology, and Research Trends. Food Sci. Anim. Resour. 2019, 4, 521–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessa, L.W.; Pieterse, E.; Marais, J.; Hoffman, L.C. Why for feed and not for human consumption? The black soldier fly larvae. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2747–2763. [Google Scholar] [CrossRef] [PubMed]
- Kouřimská, L.; Adámková, A. Nutritional and sensory quality of edible insects. NFS J. 2016, 4, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.S.; Shelomi, M. Review of Black Soldier Fly (Hermetia illucens) as Animal Feed and Human Food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- De Marco, M.; Martínez, S.; Hernandez, F.; Madrid, J.; Gai, F.; Rotolo, L.; Belforti, M.; Bergero, D.; Katz, H.; Dabbou, S. Nutritional value of two insect larval meals (Tenebrio molitor and Hermetia illucens) for broiler chickens: Apparent nutrient digestibility, apparent ileal amino acid digestibility and apparent metabolizable energy. Anim. Feed Sci. Technol. 2015, 209, 211–218. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.D.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly prepupae as a feedstuff for rainbow trout, Oncorhynchus mykiss. J. World Aquac. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Payne, C.L.R.; Scarborough, P.; Rayner, M.; Nonaka, K. A systematic review of nutrient composition data available for twelve commercially available edible insects, and comparison with reference values. Trends Food Sci. Technol. 2016, 47, 69–77. [Google Scholar] [CrossRef]
- Newton, G.L.; Booram, C.V.; Barker, R.W.; Hale, O.M. Dried Hermetia illucens larvae meal as a supplement for swine. J. Anim. Sci. 1977, 44, 395–400. [Google Scholar] [CrossRef]
- Fuso, A.; Barbi, S.; Macavei, L.I.; Luparelli, A.V.; Maistrello, L.; Montorsi, M.; Sforza, S.; Caligiani, A. Effect of the Rearing Substrate on Total Protein and Amino Acid Composition in Black Soldier Fly. Foods 2021, 10, 1773. [Google Scholar] [CrossRef]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black soldier fly biowaste treatment—Assessment of global warming potential. Waste Manag. 2019, 84, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Bosch, G.; van Zanten, H.H.E.; Zamprogna, A.; Veenenbos, M.; Meijer, N.P.; van der Fels-Klerx, H.J.; van Loon, J.J.A. Conversion of organic resources by black soldier fly larvae: Legislation, efficiency and environmental impact. J. Clean. Prod. 2019, 222, 355–363. [Google Scholar] [CrossRef]
- Jagtap, S.; Garcia-Garcia, G.; Duong, L.; Swainson, M.; Martindale, W. Codesign of Food System and Circular Economy Approaches for the Development of Livestock Feeds from Insect Larvae. Foods 2021, 10, 1701. [Google Scholar] [CrossRef]
- Belluco, S.; Losasso, C.; Maggioletti, M.; Alonzi, C.C.; Paoletti, M.G.; Ricci, A. Edible Insects in a Food Safety and Nutritional Perspective: A Critical Review. CRFSFS 2013, 12, 296–313. [Google Scholar] [CrossRef]
- European Union. Commission Regulation (EU) 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs OJ 338/1. Off. J. Eur. Union 2005, 1–26. [Google Scholar]
- EFSA. Risk profile related to production and consumption of insects as food and feed. EFSA J. 2015, 13, 4257. [Google Scholar] [CrossRef] [Green Version]
- Madigan, M.T.; Martinko, J.M.; Dunlap, P.V.; Clark, D.P. Brock Biology of Microorganisms, 12th ed.; Pearsons Education Inc.: San Francisco, CA, USA, 2009. [Google Scholar]
- European Union. Regulation (EC) 1069/2009 of the European Parliament and of the Council of 21 October 2009 laying down health rules as regards animal by-products and derived products not intended for human consumption and repealing Regulation (EC) No 1774/2002 (Animal by-products Regulation). Off. J. Eur. Union 2009, 1–33. [Google Scholar]
- Swinscoe, I.; Oliver, D.M.; Ørnsrud, R.; Quilliam, R.S. The microbial safety of seaweed as a feed component for black soldier fly (Hermetia illucens) larvae. Food Microbiol. 2020, 91, 103535. [Google Scholar] [CrossRef] [PubMed]
- Park, S.I.; Chang, B.S.; Yoe, S.M. Detection of antimicrobial substances from larvae of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Entomol. Res. 2014, 44, 58–64. [Google Scholar] [CrossRef]
- Lalander, C.H.; Fidjeland, J.; Diener, S. High waste-to-biomass conversion and efficient Salmonella spp. reduction using black soldier fly for waste recycling. Agron. Sustain. Dev. 2015, 35, 261–271. [Google Scholar] [CrossRef]
- Erickson, M.C.; Islam, M.; Sheppard, C.; Liao, J.; Doyle, M.P. Reduction of Escherichia coli O157:H7 and Salmonella enterica serovar Enteritidis in chicken manure by larvae of the black soldier fly. J. Food Prot. 2004, 67, 685–690. [Google Scholar] [CrossRef] [Green Version]
- Hogsette, J.A. New diets for production of house flies and stable flies (Diptera: Muscidae) in the laboratory. J. Econ. Entomol. 1992, 85, 2291–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheppard, D.C.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). J. Med. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oonincx, D.G.; van Itterbeeck, J.; Heetkamp, M.J.; van den Brand, H.; van Loon, J.J.A.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [Green Version]
- Nakagaki, B.J.; Defoliart, G.R. Comparison of diets for mass-rearing Acheta domesticus (Orthoptera: Gryllidae) as a novelty food, and comparison of food conversion efficiency with values reported for livestock. J. Econ. Entomol. 1991, 84, 891–896. [Google Scholar] [CrossRef]
- Oonincx, D.G.; van Broekhoven, S.; Van Huis, A.; van Loon, J.J. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [Green Version]
- Huis, A.V.; Itterbeeck, J.V.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; FAO Forestry Paper: Rome, Italy, 2013; p. 171. [Google Scholar]
- Sheppard, C. Housefly and lesser fly control utilizing the black soldier fly in manure management-systems for caged laying hens. Environ. Entomol. 1983, 12, 1439–1442. [Google Scholar] [CrossRef]
- Madau, F.A.; Arru, B.; Furesi, R.; Pulina, P. Insect farming for feed and food production from a circular business model perspective. Sustainability 2020, 12, 5418. [Google Scholar] [CrossRef]
- Choi, W.H.; Yun, J.H.; Chu, J.P.; Chu, K. Antibacterial effect of extracts of Hermetia illucens (Diptera: Stratiomyidae) larvae against Gram-negative bacteria. Entomol. Res. 2012, 42, 219–226. [Google Scholar] [CrossRef]
- Pinotti, L.; Ottoboni, M. Substrate as insect feed for bio-mass production. J. Insects Food Feed 2021, 7, 585–596. [Google Scholar] [CrossRef]
- Varotto Boccazzi, I.; Ottoboni, M.; Martin, E.; Comandatore, F.; Vallone, L.; Spranghers, T.; Eeckhout, M.; Epis, S. A survey of the mycobiota associated with larvae of the black soldier fly (Hermetia illucens) reared for feed production. PLoS ONE 2017, 12, e0182533. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Hack, M.E.; Shafi, M.E.; Alghamdi, W.Y.; Abdelnour, S.A.; Shehata, A.M.; Noreldin, A.E.; Ashour, E.A.; Swelum, A.A.; Al-Sagan, A.A.; Alkhateeb, M.; et al. Black Soldier Fly (Hermetia illucens) Meal as A Promising Feed Ingredient for Poultry: A Comprehensive Review. Agriculture 2020, 10, 339. [Google Scholar] [CrossRef]
- Caparros Megido, R.; Desmedt, S.; Blecker, C.; Béra, F.; Haubruge, É.; Alabi, T.; Francis, F. Microbiological load of edible insects found in Belgium. Insects 2017, 8, 12. [Google Scholar] [CrossRef]
- Bessa, L.W. The Evaluation of the Potential of Tenebrio molitor, Blatta lateralis, Blaptica dubia, Hermetia illucens and Naupheta cinerea for Human Consumption. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 2016. [Google Scholar]

{kind=link}
{kind=link}
| Diet D—Log CFU/g (SD) | Diet G—Log CFU/g (SD) | |||||
|---|---|---|---|---|---|---|
| Days | Substrate | Larvae | CTRL | Substrate | Larvae | CTRL |
| 1 | 7.26 (0.05) | 7.48 (0.02) | 7.32 (0.05) | 7.91 (0.00) | ||
| 2 | 6.83 (0.02) | 7.45 (0.01) | 6.86 (0.03) | 7.74 (0.01) | ||
| 3 | 4.34 (0.09) | 4.00 (0.21) | 7.30 (0.07) | 4.64 (0.21) | 4.08 (0.23) | 7.32 (0.04) |
| 6 | 3.86 (0.14) | 3.64 (0.22) | 7.60 (0.05) | 3.49 (0.20) | 3.64 (0.19) | 7.32 (0.08) |
| 8 | 3.65 (0.11) | 3.80 (0.09) | 5.30 (0.04) | 3.79 (0.10) | 3.79 (0.10) | 4.99 (0.09) |
| 10 | 3.65 (0.15) | 3.04 (0.16) | 5.32 (0.04) | 3.42 (0.10) | 3.68 (0.21) | 5.78 (0.04) |
| 13 | 4.81 (0.11) | 3.80 (0.10) | 4.70 (0.01) | 4.65 (0.12) | 3.92 (0.11) | 5.95 (0.05) |
| 17 | 3.78 (0.06) | 2.00 (0.07) | 4.30 (0.02) | 3.70 (0.04) | 2.00 (0.04) | 4.17 (0.09) |
| Log CFU/g (SD) | |||
|---|---|---|---|
| Days | Substrate | Larvae | CTRL |
| 1 | 7.29 (0.06) a,x | 7.69 (0.24) c,x | |
| 2 | 6.84 (0.24) a,y | 7.59 (0.16) c,x | |
| 3 | 4.49 (0.22) a,z | 4.04 (0.20) b,x | 7.31 (0.05) c,x |
| 6 | 3.67 (0.25) a,j | 3.64 (0.18) a,y | 7.46 (0.16) c,x |
| 8 | 3.72 (0.12) a,j | 3.80 (0.09) a,y | 5.15 (0.18) c,y |
| 10 | 3.54 (0.17) a,j | 3.36 (0.39) a,z | 5.55 (0.25) c,y |
| 13 | 4.73 (0.13) a,k | 3.86 (0.11) b,y | 5.33 (0.69) a,y |
| 17 | 3.74 (0.06) a,w | 2.00 (0.05) b,k | 4.24 (0.09) c,z |
| Diet D—Log CFU/g (SD) | Diet G—Log CFU/g (SD) | |||||
|---|---|---|---|---|---|---|
| days | Substrate | Larvae | CTRL | Substrate | Larvae | CTRL |
| 1 | 7.30 (0.09) | 7.72 (0.02) | 7.17 (0.10) | 7.79 (0.01) | ||
| 2 | 6.98 (0.04) | 7.48 (0.06) | 6.95 (0.05) | 7.38 (0.02) | ||
| 3 | 6.62 (0.02) | 6.65 (0.10) | 7.00 (0.09) | 6.55 (0.03) | 6.48 (0.08) | 7.00 (0.04) |
| 6 | 6.15 (0.06) | 5.37 (0.03) | 7.84 (0.15) | 6.21 (0.07) | 5.70 (0.01) | 7.40 (0.06) |
| 8 | 6.74 (0.05) | 5.48 (0.06) | 7.59 (0.10) | 6.40 (0.05) | 5.93 (0.05) | 7.40 (0.19) |
| 10 | 6.40 (0.35) | 5.70 (0.05) | 7.96 (0.08) | 6.48 (0.33) | 5.30 (0.04) | 7.68 (0.15) |
| 13 | 6.53 (0.04) | 4.67 (0.35) | 7.08 (0.12) | 6.08 (0.07) | 4.96 (0.33) | 7.79 (0.11) |
| 17 | 5.96 (0.11) | 4.49 (0.04) | 7.46 (0.08) | 5.91 (0.09) | 4.40 (0.03) | 7.49 (0.08) |
| Log CFU/g (SD) | |||
|---|---|---|---|
| Days | Substrate | Larvae | CTRL |
| 1 | 7.24 (0.11) a,x | 7.75 (0.04) c,x | |
| 2 | 6.96 (0.17) a,x | 7.43 (0.07) c,x | |
| 3 | 6.59 (0.05) a,y | 6.56 (0.12) a,x | 7.00 (0.06) c,y |
| 6 | 6.18 (0.07) a,z | 5.53 (0.18) b,y | 7.62 (0.26) c,x |
| 8 | 6.57 (0.19) a,y | 5.70 (0.25) b,y | 7.49 (0.17) c,x |
| 10 | 6.44 (0.26) a,y | 5.50 (0.22) b,y | 7.82 (0.19) c,x |
| 13 | 6.30 (0.26) a,y | 4.81 (0.34) b,z | 7.43 (0.40) c,x |
| 17 | 5.93 (0.09) a,z | 4.45 (0.06) b,z | 7.48 (0.07) c,x |
| Log CFU/g (SD) in Larvae | ||
|---|---|---|
| Days | Salmonella enterica Typhimurium | L. monocytogenes |
| 3 | 4.04 (0.20) a | 6.56 (0.12) b |
| 6 | 3.64 (0.18) a | 5.53 (0.18) b |
| 8 | 3.80 (0.09) a | 5.70 (0.25) b |
| 10 | 3.36 (0.39) a | 5.50 (0.22) b |
| 13 | 3.86 (0.11) b | 4.81 (0.34) b |
| 17 | 2.00 (0.05) b | 4.45 (0.06) b |
| Diet D (SD) | Diet G (SD) | |||||||
|---|---|---|---|---|---|---|---|---|
| aw | pH | aw | pH | |||||
| Days | CTRL | larvae | CTRL | Larvae | CTRL | Larvae | CTRL | Larvae |
| 1 | 0.980 (0.001) | 0.980 (0.001) | 6.69 (0.01) | 6.87 (0.01) | 0.98 (0.02) | 0.980 (0.02) | 6.59 (0.02) | 7.53 (0.01) |
| 2 | 0.982 (0.002) | 0.980 (0.001) | 6.58 (0.02) | 7.47 (0.02) | 0.98 (0.02) | 0.943 (0.01) | 6.83 (0.02) | 6.64 (0.02) |
| 6 | 0.982 (0.001) | 0.970 (0.003) | 8.08 (0.01) | 8.23 (0.01) | 0.98 (0.01) | 0.971 (0.01) | 7.00 (0.02) | 6.88 (0.01) |
| 8 | 0.971 (0.002) | 0.980 (0.002) | 7.95 (0.02) | 8.52 (0.01) | 0.97 (0.01) | 0.971 (0.01) | 7.78 (0.01) | 6.97 (0.02) |
| 10 | 0.943 (0.002) | 0.970 (0.003) | 7.76 (0.01) | 7.47 (0.01) | 0.945 (0.01) | 0.972 (0.03) | 7.08 (0.02) | 7.23 (0.01) |
| 13 | 0.773 (0.002) | 0.970 (0.001) | 7.30 (0.02) | 8.23 (0.02) | 0.85 (0.01) | 0.973 (0.01) | 7.15 (0.02) | 6.64 (0.01) |
| 17 | 0.661 (0.005) | 0.980 (0.002) | 6.98 (0.01) | 8.35 (0.01) | 0.64 (0.01) | 0.971 (0.02) | 7.18 (0.01) | 6.97 (0.01) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grisendi, A.; Defilippo, F.; Lucchetti, C.; Listorti, V.; Ottoboni, M.; Dottori, M.; Serraino, A.; Pinotti, L.; Bonilauri, P. Fate of Salmonella enterica Typhimurium and Listeria monocytogenes in Black Soldier Fly (Hermetia illucens) Larvae Reared on Two Artificial Diets. Foods 2022, 11, 2208. https://doi.org/10.3390/foods11152208
Grisendi A, Defilippo F, Lucchetti C, Listorti V, Ottoboni M, Dottori M, Serraino A, Pinotti L, Bonilauri P. Fate of Salmonella enterica Typhimurium and Listeria monocytogenes in Black Soldier Fly (Hermetia illucens) Larvae Reared on Two Artificial Diets. Foods. 2022; 11(15):2208. https://doi.org/10.3390/foods11152208
Chicago/Turabian StyleGrisendi, Annalisa, Francesco Defilippo, Chiara Lucchetti, Valeria Listorti, Matteo Ottoboni, Michele Dottori, Andrea Serraino, Luciano Pinotti, and Paolo Bonilauri. 2022. "Fate of Salmonella enterica Typhimurium and Listeria monocytogenes in Black Soldier Fly (Hermetia illucens) Larvae Reared on Two Artificial Diets" Foods 11, no. 15: 2208. https://doi.org/10.3390/foods11152208
APA StyleGrisendi, A., Defilippo, F., Lucchetti, C., Listorti, V., Ottoboni, M., Dottori, M., Serraino, A., Pinotti, L., & Bonilauri, P. (2022). Fate of Salmonella enterica Typhimurium and Listeria monocytogenes in Black Soldier Fly (Hermetia illucens) Larvae Reared on Two Artificial Diets. Foods, 11(15), 2208. https://doi.org/10.3390/foods11152208