
Characterization of Purified Mulberry Leaf Glycoprotein and Its Immunoregulatory Effect on Cyclophosphamide-Treated Mice

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
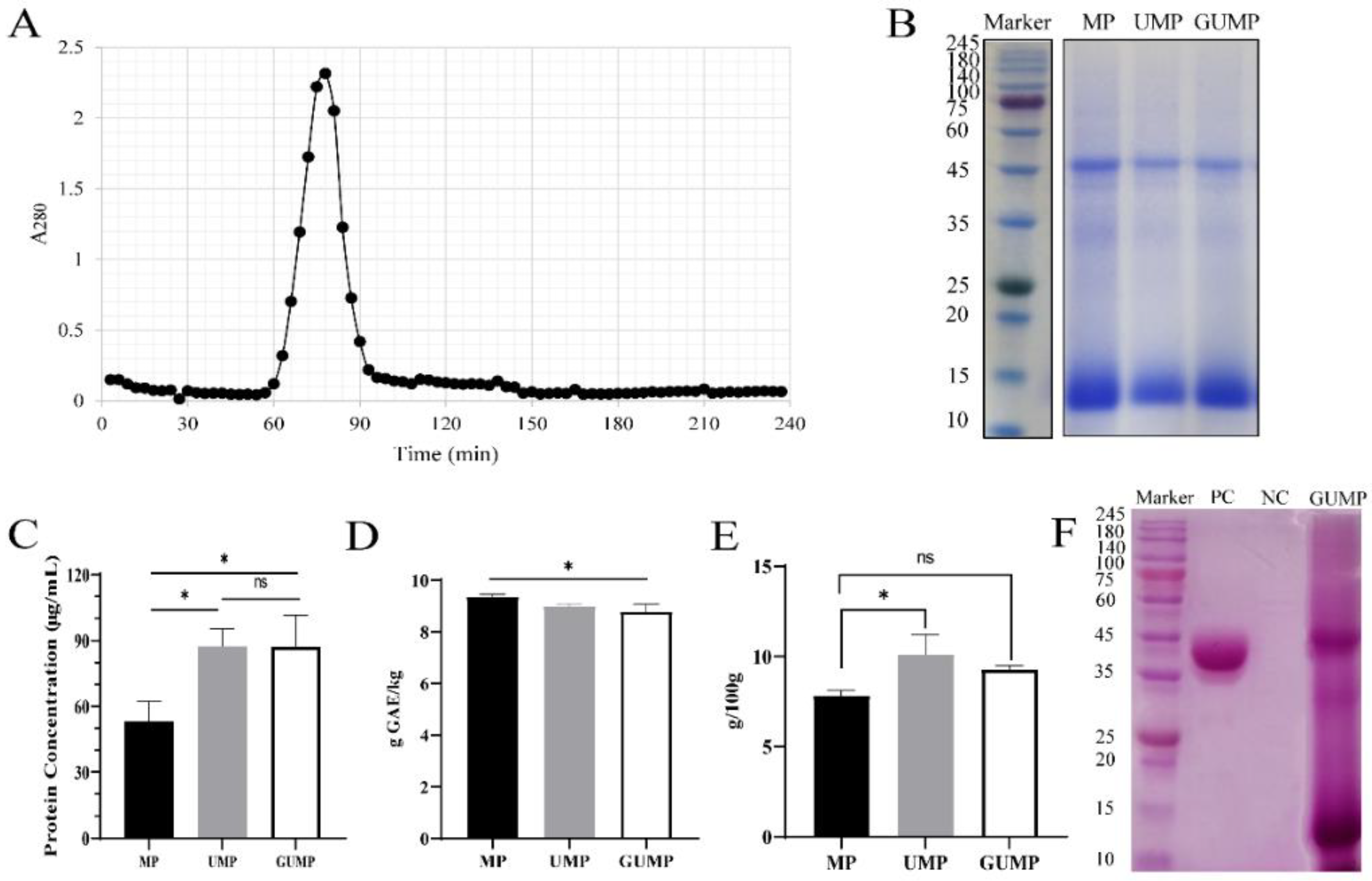
2.2. Ultrafiltration and Column Chromatography
2.3. Protein, Total Sugar, and Total Phenol Content (TPC) of MP, UMP, and GUMP
2.4. SDS-PAGE and Glycoprotein Staining
2.5. Determination of Monosaccharide Composition
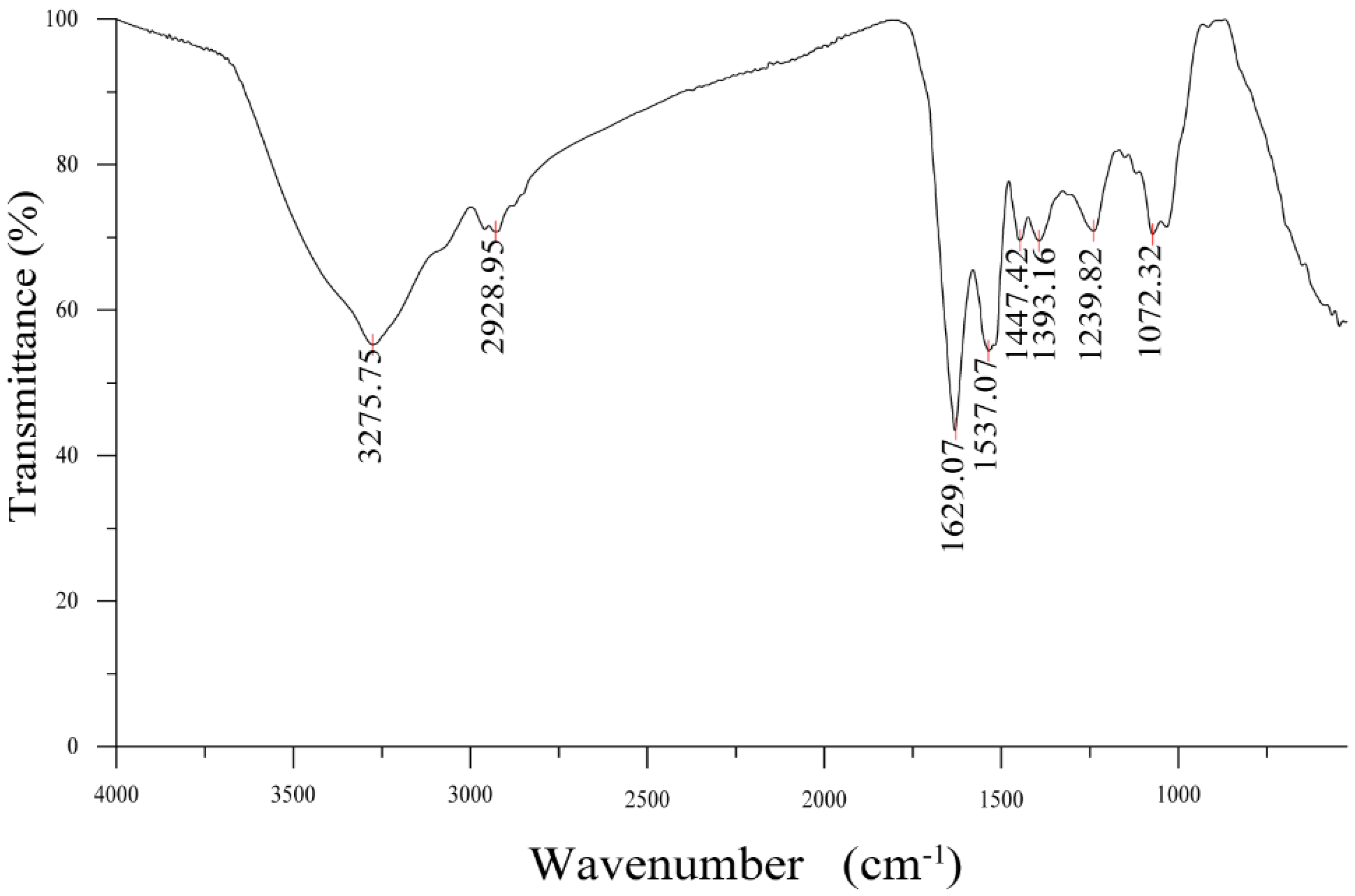
2.6. Fourier Transform Infrared Spectroscopy (FTIR)
2.7. LC–MS/MS Analysis
2.8. Animals and Experimental Design
2.9. Determination of Cytokines and Immunoglobulins
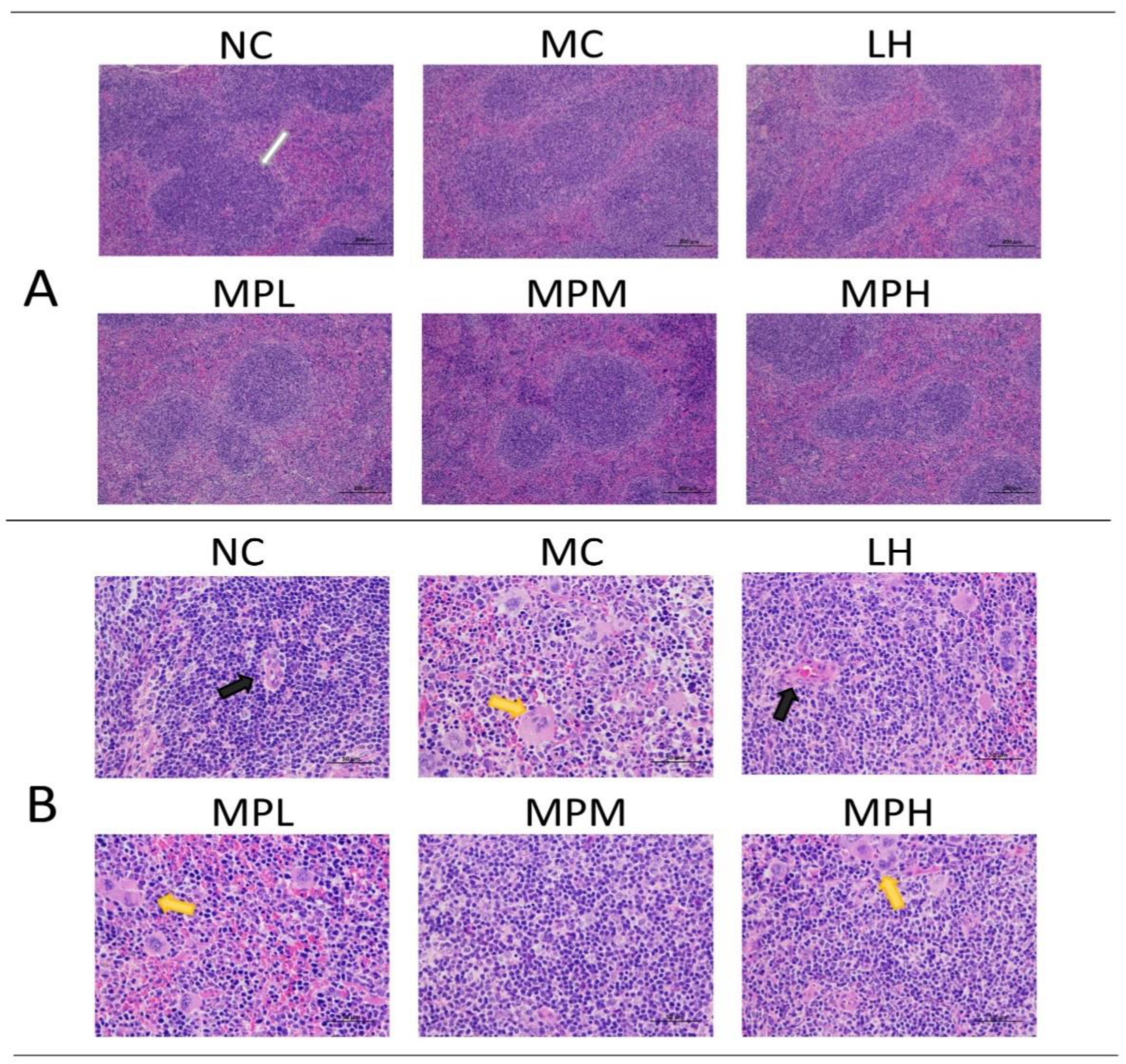
2.10. Histopathological Staining of the Spleen
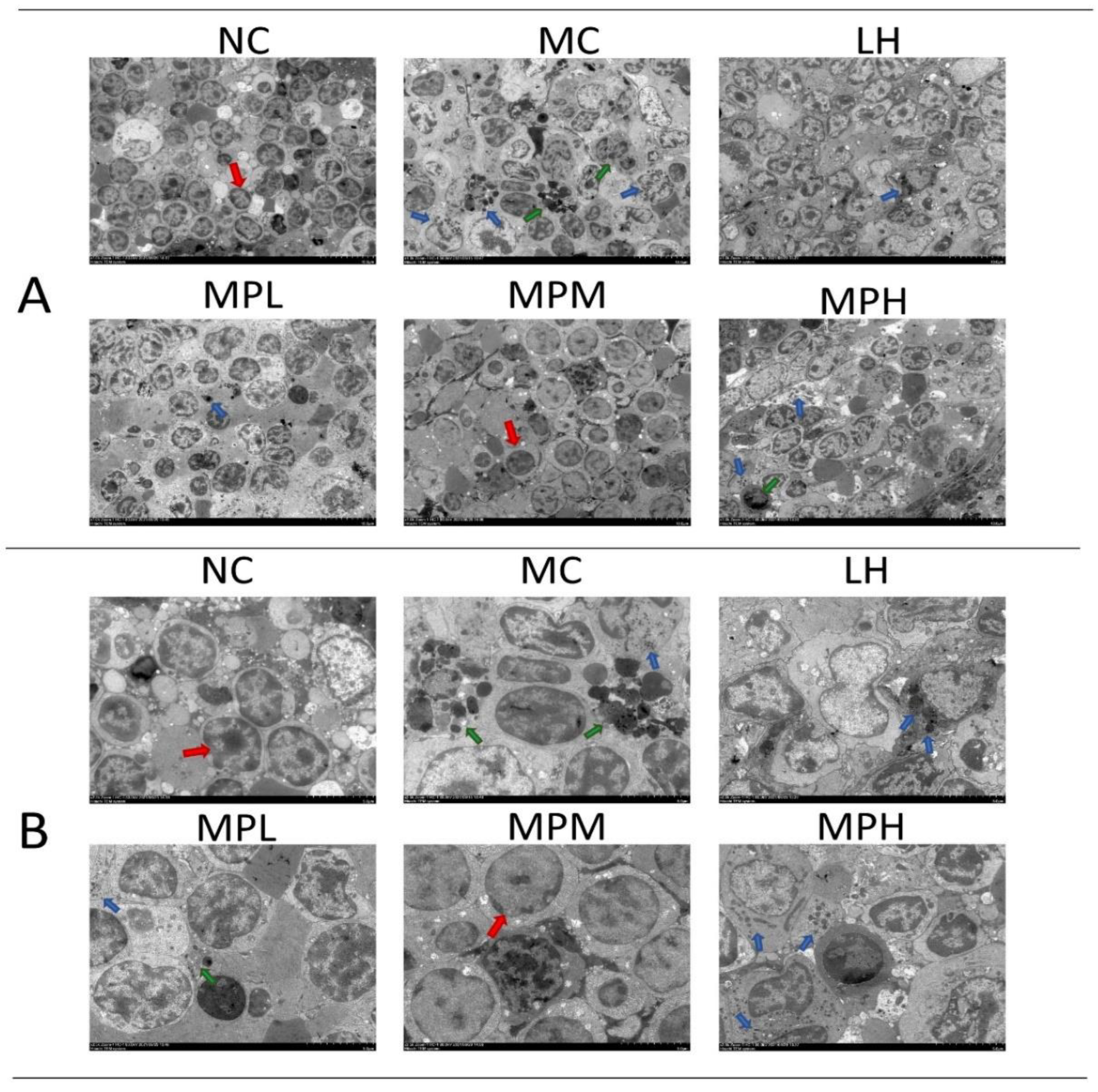
2.11. Transmission Electron Microscopy (TEM) Analysis
2.12. Determination of Fatty Acids
2.13. Gut Microbiota Analysis
2.14. Correlation Analysis
2.15. Statistical Analysis
3. Results and Discussion
3.1. Purification, Molecular Weight Distribution, and Chemical Composition of MP, UMP, and GUMP
3.2. Monosaccharide Compositions of GUMP
3.3. Characterization of GUMP
3.3.1. FTIR Analysis
3.3.2. LC–MS/MS Analysis of GUMP
3.4. Immunomodulatory Ability of GUMP
3.4.1. Effects of GUMP on the Body Weight and Thymus Indexes in CP-Treated Mice
3.4.2. Effects of GUMP on Serum Cytokine and Immunoglobulin in CP-Treated Mice
3.4.3. The Effects of GUMP on Histomorphology of Spleen in CP-Treated Mice
3.4.4. Effects of GUMP on SCFA Production in Mouse Feces
3.4.5. The Effects of GUMP on the Gut Microbiota Composition
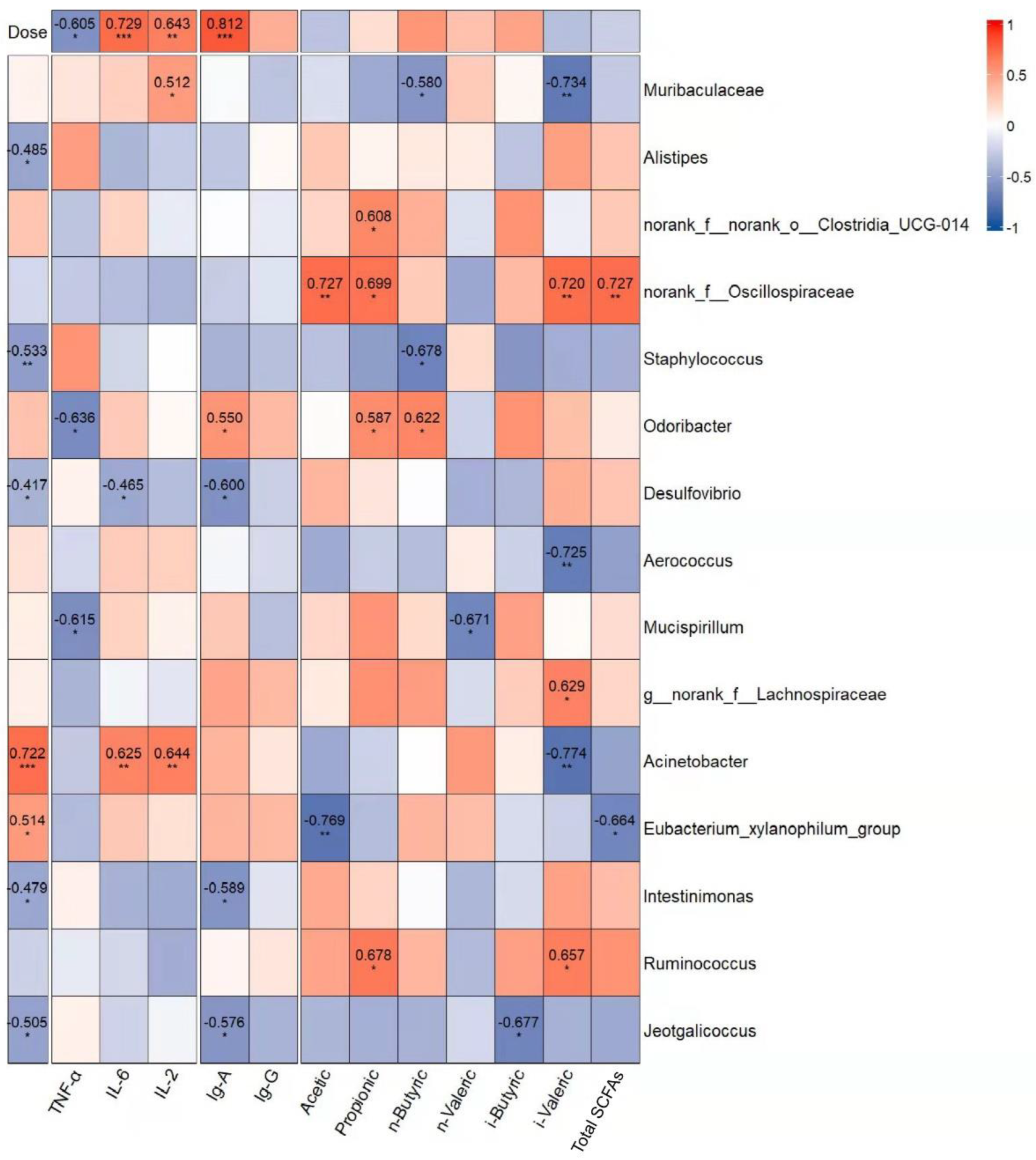
3.4.6. Correlation Analysis of Gavage Dose, Dominant Genera, and Immune Indexes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Boven, J.F.M.; de Jong-van den Berg, L.T.W.; Vegter, S. Inhaled Corticosteroids and the Occurrence of Oral Candidiasis: A Prescription Sequence Symmetry Analysis. Drug Saf. 2013, 36, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Waxman, D.J. Immunogenic Chemotherapy: Dose and Schedule Dependence and Combination with Immunotherapy. Cancer Lett. 2018, 419, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Ahlmann, M.; Hempel, G. The Effect of Cyclophosphamide on the Immune System: Implications for Clinical Cancer Therapy. Cancer Chemother. Pharmacol. 2016, 78, 661–671. [Google Scholar] [CrossRef]
- Voelcker, G. The Mechanism of Action of Cyclophosphamide and Its Consequences for the Development of a New Generation of Oxazaphosphorine Cytostatics. Sci. Pharm. 2020, 88, 42. [Google Scholar] [CrossRef]
- Puebla, L.E.J. Fungal Infections in Immunosuppressed Patients; IntechOpen: London, UK, 2012; ISBN 978-953-51-0791-0. [Google Scholar]
- Moreau, R.; Arroyo, V. Immunosuppression and Increased Risk of Bacterial Infections in Cirrhosis. Infect. Microbes Dis. 2021, 3, 115–116. [Google Scholar] [CrossRef]
- PubChem Levamisole Hydrochloride. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/27944 (accessed on 26 November 2021).
- Ding, Y.; Yan, Y.; Chen, D.; Ran, L.; Mi, J.; Lu, L.; Jing, B.; Li, X.; Zeng, X.; Cao, Y. Modulating Effects of Polysaccharides from the Fruits of Lycium Barbarum on the Immune Response and Gut Microbiota in Cyclophosphamide-Treated Mice. Food Funct. 2019, 10, 3671–3683. [Google Scholar] [CrossRef]
- Zhu, G.; Jiang, Y.; Yao, Y.; Wu, N.; Luo, J.; Hu, M.; Tu, Y.; Xu, M. Ovotransferrin Ameliorates the Dysbiosis of Immunomodulatory Function and Intestinal Microbiota Induced by Cyclophosphamide. Food Funct. 2019, 10, 1109–1122. [Google Scholar] [CrossRef]
- Wu, F.; Huang, H. Surface Morphology and Protective Effect of Hericium Erinaceus Polysaccharide on Cyclophosphamide-Induced Immunosuppression in Mice. Carbohydr. Polym. 2021, 251, 116930. [Google Scholar] [CrossRef]
- Moore, F.A.; Phillips, S.; McClain, C.; Patel, J.J.; Martindale, R. Nutrition Support for Persistent Inflammation, Immunosuppression, and Catabolism Syndrome. Nutr. Clin. Pract. 2017, 32, 121S–127S. [Google Scholar] [CrossRef]
- Ueda, N.; Kayashita, J.; Moriguchi, S.; Kishino, Y. Effect of Dietary Protein Levels on Immune Function of Exercised Rats. Nutr. Res. 1990, 10, 429–437. [Google Scholar] [CrossRef]
- Chandra, R.K. Nutrition and the Immune System: An Introduction. Am. J. Clin. Nutr. 1997, 66, 460S–463S. [Google Scholar] [CrossRef]
- Huang, J.; Huang, J.; Li, Y.; Wang, Y.; Wang, F.; Qiu, X.; Liu, X.; Li, H. Sodium Alginate Modulates Immunity, Intestinal Mucosal Barrier Function, and Gut Microbiota in Cyclophosphamide-Induced Immunosuppressed BALB/c Mice. J. Agric. Food Chem. 2021, 69, 7064–7073. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Yan, Y.; Chen, D.; Zhao, Y.; Dong, W.; Zeng, X.; Cao, Y. Ascorbic Acid Derivative 2-O-β-d-Glucopyranosyl-l-Ascorbic Acid from the Fruit of Lycium Barbarum Modulates Microbiota in the Small Intestine and Colon and Exerts an Immunomodulatory Effect on Cyclophosphamide-Treated BALB/c Mice. J. Agric. Food Chem. 2020, 68, 11128–11143. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Bai, B.; Zhou, Q.; Niu, J.; Yuan, J.; Zhang, H.; Jia, J.; Zhao, W.; Chen, H. Dietary Supplementation with Polysaccharides from Ziziphus Jujuba Cv. Pozao Intervenes in Immune Response via Regulating Peripheral Immunity and Intestinal Barrier Function in Cyclophosphamide-Induced Mice. Food Funct. 2020, 11, 5992–6006. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Xie, J.; Liu, X.; Shen, M. Sulfated Modification Enhances the Modulatory Effect of Yam Polysaccharide on Gut Microbiota in Cyclophosphamide-Treated Mice. Food Res. Int. 2021, 145, 110393. [Google Scholar] [CrossRef]
- Xiang, X.-W.; Zheng, H.-Z.; Wang, R.; Chen, H.; Xiao, J.-X.; Zheng, B.; Liu, S.-L.; Ding, Y.-T. Ameliorative Effects of Peptides Derived from Oyster (Crassostrea Gigas) on Immunomodulatory Function and Gut Microbiota Structure in Cyclophosphamide-Treated Mice. Mar. Drugs 2021, 19, 456. [Google Scholar] [CrossRef]
- Chen, S.; Wang, J.; Fang, Q.; Dong, N.; Fang, Q.; Cui, S.W.; Nie, S. A Polysaccharide from Natural Cordyceps Sinensis Regulates the Intestinal Immunity and Gut Microbiota in Mice with Cyclophosphamide-Induced Intestinal Injury. Food Funct. 2021, 12, 6271–6282. [Google Scholar] [CrossRef]
- Lown, M.; Fuller, R.; Lightowler, H.; Fraser, A.; Gallagher, A.; Stuart, B.; Byrne, C.; Lewith, G. Mulberry-Extract Improves Glucose Tolerance and Decreases Insulin Concentrations in Normoglycaemic Adults: Results of a Randomised Double-Blind Placebo-Controlled Study. PLoS ONE 2017, 12, e0172239. [Google Scholar] [CrossRef]
- Sun, C.; Wu, W.; Ma, Y.; Min, T.; Lai, F.; Wu, H. Physicochemical, Functional Properties, and Antioxidant Activities of Protein Fractions Obtained from Mulberry (Morus Atropurpurea Roxb.) Leaf. Int. J. Food Prop. 2017, 20, S3311–S3325. [Google Scholar] [CrossRef]
- Sun, C.; Shan, Y.; Tang, X.; Han, D.; Wu, X.; Wu, H.; Hosseininezhad, M. Effects of Enzymatic Hydrolysis on Physicochemical Property and Antioxidant Activity of Mulberry (Morus Atropurpurea Roxb.) Leaf Protein. Food Sci. Nutr. 2021, 9, 5379–5390. [Google Scholar] [CrossRef]
- Sun, C.; Tang, X.; Shao, X.; Han, D.; Zhang, H.; Shan, Y.; Gooneratne, R.; Shi, L.; Wu, X.; Hosseininezhad, M. Mulberry (Morus Atropurpurea Roxb.) Leaf Protein Hydrolysates Ameliorate Dextran Sodium Sulfate-Induced Colitis via Integrated Modulation of Gut Microbiota and Immunity. J. Funct. Foods 2021, 84, 104575. [Google Scholar] [CrossRef]
- Sun, C.; Wu, W.; Yin, Z.; Fan, L.; Ma, Y.; Lai, F.; Wu, H. Effects of Simulated Gastrointestinal Digestion on the Physicochemical Properties, Erythrocyte Haemolysis Inhibitory Ability and Chemical Antioxidant Activity of Mulberry Leaf Protein and Its Hydrolysates. Int. J. Food Sci. Technol. 2018, 53, 282–295. [Google Scholar] [CrossRef]
- Nielsen, S.S. Phenol-Sulfuric Acid Method for Total Carbohydrates. In Food Analysis Laboratory Manual; Nielsen, S.S., Ed.; Food Science Texts Series; Springer: Boston, MA, USA, 2010; pp. 47–53. ISBN 978-1-4419-1463-7. [Google Scholar]
- Ibrahim, H.; Aoussar, N.; Mhand, R.A.; Rhallabi, N.; Oili, A.D.; Mellouki, F. In Vitro Antioxidant and Antistaphylococcal Properties of Leaf Extracts of Ocotea Comorensis Kosterm (Lauraceae). Biocatal. Agric. Biotechnol. 2021, 31, 101892. [Google Scholar] [CrossRef]
- Lei, Y.; Guo, K.; Zhang, Y.; Zhang, X.; Qin, L.; Wang, X.; Zhu, H.; Guo, Y.; Yang, W.; Li, B.; et al. Adhesive Property and Mechanism of Silkworm Egg Glue Protein. Acta Biomater. 2021, 134, 499–512. [Google Scholar] [CrossRef]
- Feng, W.; Dong, T.; Li, K.; Wang, T.; Chen, Z.; Wang, R. Characterization of Binding Behaviors of Cd2+ to Rice Proteins. Food Chem. 2019, 275, 186–192. [Google Scholar] [CrossRef]
- Gautam, R.; Vanga, S.; Ariese, F.; Umapathy, S. Review of Multidimensional Data Processing Approaches for Raman and Infrared Spectroscopy. EPJ Tech. Instrum. 2015, 2, 1–38. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal Sample Preparation Method for Proteome Analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Zhao, G.; Nyman, M.; Åke Jönsson, J. Rapid Determination of Short-Chain Fatty Acids in Colonic Contents and Faeces of Humans and Rats by Acidified Water-Extraction and Direct-Injection Gas Chromatography. Biomed. Chromatogr. 2006, 20, 674–682. [Google Scholar] [CrossRef]
- Shao, X.; Sun, C.; Tang, X.; Zhang, X.; Han, D.; Liang, S.; Qu, R.; Hui, X.; Shan, Y.; Hu, L.; et al. Anti-Inflammatory and Intestinal Microbiota Modulation Properties of Jinxiang Garlic (Allium Sativum L.) Polysaccharides toward Dextran Sodium Sulfate-Induced Colitis. J. Agric. Food Chem. 2020, 68, 12295–12309. [Google Scholar] [CrossRef]
- Izumi, M.; Tsunoda, H.; Suzuki, Y.; Makino, A.; Ishida, H. RBCS1A and RBCS3B, Two Major Members within the Arabidopsis RBCS Multigene Family, Function to Yield Sufficient Rubisco Content for Leaf Photosynthetic Capacity. J. Exp. Bot. 2012, 63, 2159–2170. [Google Scholar] [CrossRef]
- Di Stefano, E.; Agyei, D.; Njoku, E.N.; Udenigwe, C.C. Plant RuBisCo: An Underutilized Protein for Food Applications. J. Am. Oil Chem. Soc. 2018, 95, 1063–1074. [Google Scholar] [CrossRef]
- Martin, A.H.; Castellani, O.; de Jong, G.A.; Bovetto, L.; Schmitt, C. Comparison of the Functional Properties of RuBisCO Protein Isolate Extracted from Sugar Beet Leaves with Commercial Whey Protein and Soy Protein Isolates. J. Sci. Food Agric. 2019, 99, 1568–1576. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, K.; Bose, A.; Chakraborty, T.; Sarkar, K.; Goswami, S.; Pal, S.; Baral, R. Restoration of Dysregulated CC Chemokine Signaling for Monocyte/Macrophage Chemotaxis in Head and Neck Squamous Cell Carcinoma Patients by Neem Leaf Glycoprotein Maximizes Tumor Cell Cytotoxicity. Cell. Mol. Immunol. 2010, 7, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Xie, M.; Fu, Z.; Wan, Y.; Yan, A. Study on the Purification and Chemical Compositions of Tea Glycoprotein. Carbohydr. Polym. 2008, 71, 626–633. [Google Scholar] [CrossRef]
- Ji, H.-Y.; Dai, K.-Y.; Liu, C.; Yu, J.; Jia, X.-Y.; Liu, A.-J. Preparation, Antioxidant and Immunoregulatory Activities of a Macromolecular Glycoprotein from Salvia Miltiorrhiza. Foods 2022, 11, 705. [Google Scholar] [CrossRef] [PubMed]
- Crafts-Brandner, S.J.; Salvucci, M.E. The Rubisco Complex Protein: A Protein Induced by Fruit Removal That Forms a Complex with Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase. Planta 1994, 194, 110–116. [Google Scholar] [CrossRef]
- Yun, L.; Wu, T.; Li, W.; Zhang, M. Wheat Germ Glycoprotein Regionally Modulates Immunosuppressed Mouse Intestinal Immunity Function from Early Life to Adulthood. Food Funct. 2021, 12, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, X. Effects of Cyclophosphamide on Immune System and Gut Microbiota in Mice. Microbiol. Res. 2015, 171, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.C.; Schrama, D. The Dark Side of Cyclophosphamide: Cyclophosphamide-Mediated Ablation of Regulatory T Cells. J. Investig. Dermatol. 2013, 133, 1462–1465. [Google Scholar] [CrossRef]
- Zhu, G.; Luo, J.; Du, H.; Jiang, Y.; Tu, Y.; Yao, Y.; Xu, M. Ovotransferrin Enhances Intestinal Immune Response in Cyclophosphamide-Induced Immunosuppressed Mice. Int. J. Biol. Macromol. 2018, 120, 1–9. [Google Scholar] [CrossRef]
- Kamimura, D.; Hirano, T.; Murakami, M. Interleukin-6. In Reference Module in Neuroscience and Biobehavioral Psychology; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 978-0-12-809324-5. [Google Scholar]
- Lahans, T. Chapter 1—General Pathophysiology and Treatment: Conventional and Chinese Medicine. In Integrating Conventional and Chinese Medicine in Cancer Care; Lahans, T., Ed.; Churchill Livingstone: Edinburgh, UK, 2007; pp. 1–34. ISBN 978-0-443-10063-5. [Google Scholar]
- Wang, H.; Wang, M.; Chen, J.; Tang, Y.; Dou, J.; Yu, J.; Xi, T.; Zhou, C. A Polysaccharide from Strongylocentrotus Nudus Eggs Protects against Myelosuppression and Immunosuppression in Cyclophosphamide-Treated Mice. Int. Immunopharmacol. 2011, 11, 1946–1953. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, P.; Quinello, C.; Silveira-Lessa, A.L.; Zago, C.A.; Carneiro-Sampaio, M. IgG Placental Transfer in Healthy and Pathological Pregnancies. Clin. Dev. Immunol. 2011, 2012, 985646. [Google Scholar] [CrossRef] [PubMed]
- Daha, M.R.; van Kooten, C. Role of Complement in IgA Nephropathy. J. Nephrol. 2016, 29, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Cao, N.; Chen, F.; Qian, L.; Wang, Y.; Huang, Y.; Tian, Y.; Xu, D.; Li, W. Polysaccharide of Atractylodes Macrocephala Koidz (PAMK) Alleviates Cyclophosphamide-Induced Immunosuppression in Mice by Upregulating CD28/IP3R/PLCγ-1/AP-1/NFAT Signal Pathway. Front. Pharmacol. 2020, 11, 529657. [Google Scholar] [CrossRef]
- Diaz-Montero, C.M.; Wang, Y.; Shao, L.; Feng, W.; Zidan, A.-A.; Pazoles, C.J.; Montero, A.J.; Zhou, D. The Glutathione Disulfide Mimetic NOV-002 Inhibits Cyclophosphamide-Induced Hematopoietic and Immune Suppression by Reducing Oxidative Stress. Free Radic. Biol. Med. 2012, 52, 1560–1568. [Google Scholar] [CrossRef]
- Madondo, M.T.; Quinn, M.; Plebanski, M. Low Dose Cyclophosphamide: Mechanisms of T Cell Modulation. Cancer Treat. Rev. 2016, 42, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Osawa, T.; Davies, D.; Hartley, J.A. Mechanism of Cell Death Resulting from DNA Interstrand Cross-Linking in Mammalian Cells. Cell Death Dis. 2011, 2, e187. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Li, L.; Rezaei, A.; Eslamfam, S.; Che, D.; Ma, X. Metabolites of Dietary Protein and Peptides by Intestinal Microbes and Their Impacts on Gut. Curr. Protein Pept. Sci. 2015, 16, 646–654. [Google Scholar] [CrossRef]
- Cani, P.D.; Van Hul, M.; Lefort, C.; Depommier, C.; Rastelli, M.; Everard, A. Microbial Regulation of Organismal Energy Homeostasis. Nat. Metab. 2019, 1, 34–46. [Google Scholar] [CrossRef]
- Ratajczak, W.; Rył, A.; Mizerski, A.; Walczakiewicz, K.; Sipak, O.; Laszczyńska, M. Immunomodulatory Potential of Gut Microbiome-Derived Short-Chain Fatty Acids (SCFAs). Acta Biochim. Pol. 2019, 66, 1–12. [Google Scholar] [CrossRef]
- Yip, W.; Hughes, M.R.; Li, Y.; Cait, A.; Hirst, M.; Mohn, W.W.; McNagny, K.M. Butyrate Shapes Immune Cell Fate and Function in Allergic Asthma. Front. Immunol. 2021, 12, 299. [Google Scholar] [CrossRef] [PubMed]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chang, Y.; Gao, Y.; Wang, X.; Chen, X.; Wang, Y.; Xue, C.; Tang, Q. Dietary Fucoidan of Acaudina Molpadioides Alters Gut Microbiota and Mitigates Intestinal Mucosal Injury Induced by Cyclophosphamide. Food Funct. 2017, 8, 3383–3393. [Google Scholar] [CrossRef] [PubMed]
- Moertel, C.G.; Fleming, T.R.; Macdonald, J.S.; Haller, D.G.; Laurie, J.A.; Goodman, P.J.; Ungerleider, J.S.; Emerson, W.A.; Tormey, D.C.; Glick, J.H.; et al. Levamisole and Fluorouracil for Adjuvant Therapy of Resected Colon Carcinoma. N. Engl. J. Med. 1990, 322, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.; Feng, Z.; Hu, S.; Cui, L.; Qiao, T.; Dai, L.; Qi, P.; Zhang, L.; Liu, Y.; Li, J. Effects of Polysaccharides from Wild Morels on Immune Response and Gut Microbiota Composition in Non-Treated and Cyclophosphamide-Treated Mice. Food Funct. 2020, 11, 4291–4303. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic Distribution of Three Pathways for Propionate Production within the Human Gut Microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Gálvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and Cultivation Study of Muribaculaceae Reveals Novel Species, Host Preference, and Functional Potential of This yet Undescribed Family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A Candidate for the next-Generation Probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Xing, C.; Wang, M.; Ajibade, A.A.; Tan, P.; Fu, C.; Chen, L.; Zhu, M.; Hao, Z.-Z.; Chu, J.; Yu, X.; et al. Microbiota Regulate Innate Immune Signaling and Protective Immunity against Cancer. Cell Host Microbe 2021, 29, 959–974. [Google Scholar] [CrossRef]
- Waite, D.W.; Chuvochina, M.; Pelikan, C.; Parks, D.H.; Yilmaz, P.; Wagner, M.; Loy, A.; Naganuma, T.; Nakai, R.; Whitman, W.B.; et al. Proposal to Reclassify the Proteobacterial Classes Deltaproteobacteria and Oligoflexia, and the Phylum Thermodesulfobacteria into Four Phyla Reflecting Major Functional Capabilities. Int. J. Syst. Evol. Microbiol. 2020, 70, 5972–6016. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, F.; Lu, J.; Shi, J.; Guan, J.; Yan, F.; Li, B.; Huo, G. Probiotic Mixture of Lactobacillus Plantarum Strains Improves Lipid Metabolism and Gut Microbiota Structure in High Fat Diet-Fed Mice. Front. Microbiol. 2020, 11, 512. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monosaccharide | Molar Ratio |
|---|---|
| Fucose | 0.07 |
| Galactosamine hydrochloride | 0.02 |
| Rhamnose | 0.50 |
| Arabinose | 2.22 |
| Glucosamine hydrochloride | 0.32 |
| Galactose | 0.98 |
| Glucose | 2.48 |
| Xylose | 0.17 |
| Mannose | 0.30 |
| Galacturonic acid | 0.25 |
| Secondary Structure | Amide I Band Analysis | ||
|---|---|---|---|
| Characteristic Peaks (cm−1) | Peak Area | Percentage Composition | |
| α-helix | 1667 | 16.86 | 17.4% |
| β-sheet | 1608, 1630 | 45.65 | 47.1% |
| β-turn | 1691 | 18.83 | 19.4% |
| Random coil | 1648 | 15.58 | 16.1% |
| Accession | −10 lgP | Coverage (%) | Peptides | Unique | Spec GUMP | Average Mass | Description |
|---|---|---|---|---|---|---|---|
| AHW81907.1 | 364.27 | 81 | 51 | 51 | 79 | 16,951 | mannose-binding lectin (Morus alba var. atropurpurea) |
| AJF21883.1 | 309.36 | 75 | 33 | 33 | 45 | 16,752 | 18 kD winter accumulating protein (Morus alba var. atropurpurea) |
| QVD39028.1 | 155.87 | 12 | 7 | 6 | 9 | 52,668 | ribulose-1 5-bisphosphate carboxylase/oxygenase large subunit (chloroplast) (Morus alba var. atropurpurea) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, Y.; Sun, C.; Li, J.; Shao, X.; Wu, J.; Zhang, M.; Yao, H.; Wu, X. Characterization of Purified Mulberry Leaf Glycoprotein and Its Immunoregulatory Effect on Cyclophosphamide-Treated Mice. Foods 2022, 11, 2034. https://doi.org/10.3390/foods11142034
Shan Y, Sun C, Li J, Shao X, Wu J, Zhang M, Yao H, Wu X. Characterization of Purified Mulberry Leaf Glycoprotein and Its Immunoregulatory Effect on Cyclophosphamide-Treated Mice. Foods. 2022; 11(14):2034. https://doi.org/10.3390/foods11142034
Chicago/Turabian StyleShan, Yangwei, Chongzhen Sun, Jishan Li, Xin Shao, Junfeng Wu, Mengmeng Zhang, Hong Yao, and Xiyang Wu. 2022. "Characterization of Purified Mulberry Leaf Glycoprotein and Its Immunoregulatory Effect on Cyclophosphamide-Treated Mice" Foods 11, no. 14: 2034. https://doi.org/10.3390/foods11142034
APA StyleShan, Y., Sun, C., Li, J., Shao, X., Wu, J., Zhang, M., Yao, H., & Wu, X. (2022). Characterization of Purified Mulberry Leaf Glycoprotein and Its Immunoregulatory Effect on Cyclophosphamide-Treated Mice. Foods, 11(14), 2034. https://doi.org/10.3390/foods11142034